A Synthetic Approach for Biosynthesis of Miquelianin and Scutellarin A in Escherichia coli

 , ,
, , 
(This article belongs to the Section Applied Biosciences and Bioengineering)
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Cloning and Expression of Recombinant Proteins
2.3. Construction of Sugar Cassettes
2.4. Whole-Cell Biotransformation and Flavonoid Extraction
2.5. Product Analysis and Quantification
3. Results and Discussion
3.1. Selection of Plant UGTs and Their Activities
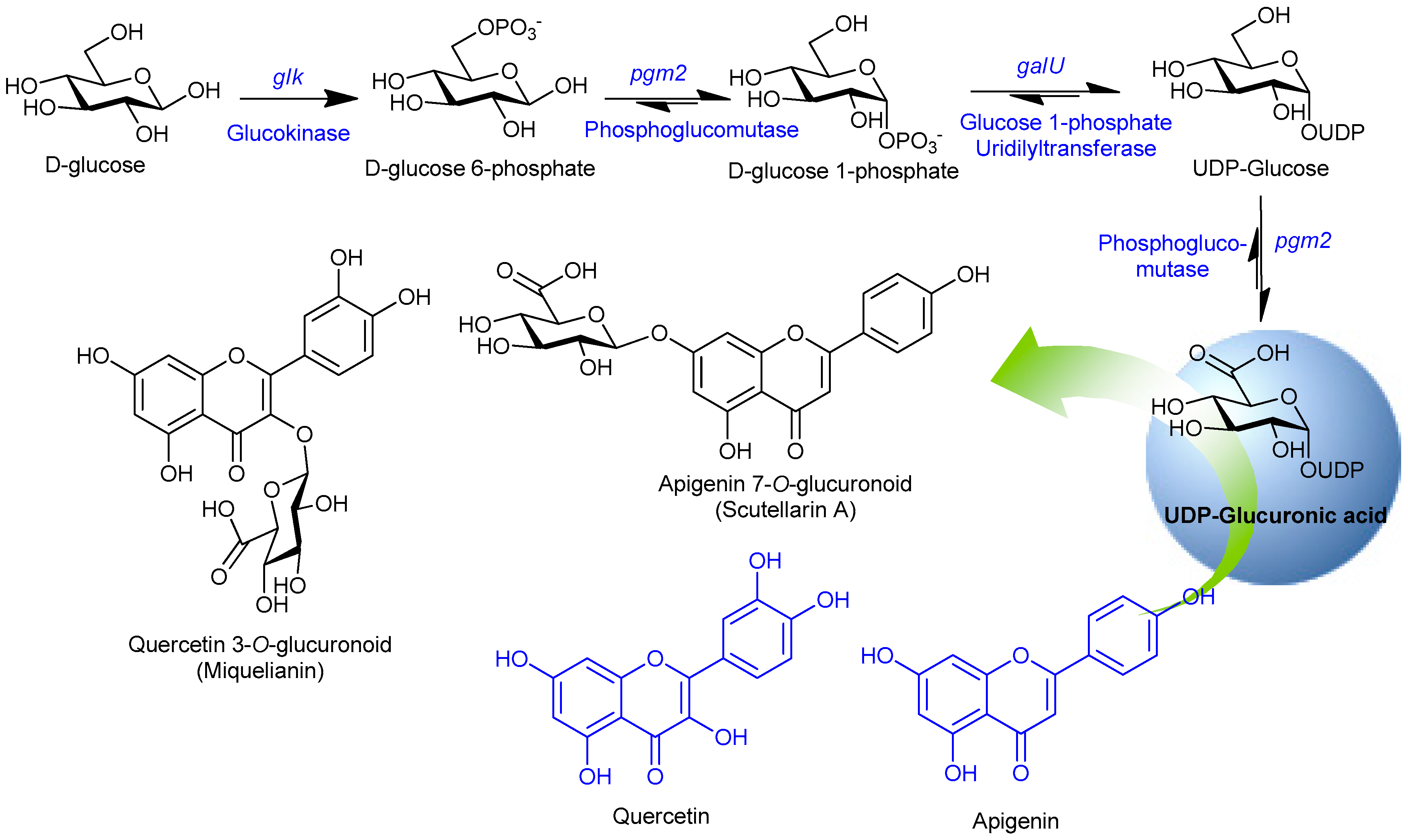
3.2. Cloning and Expression of NDP-Sugar Pathway Genes and Assembly in a Single Vector
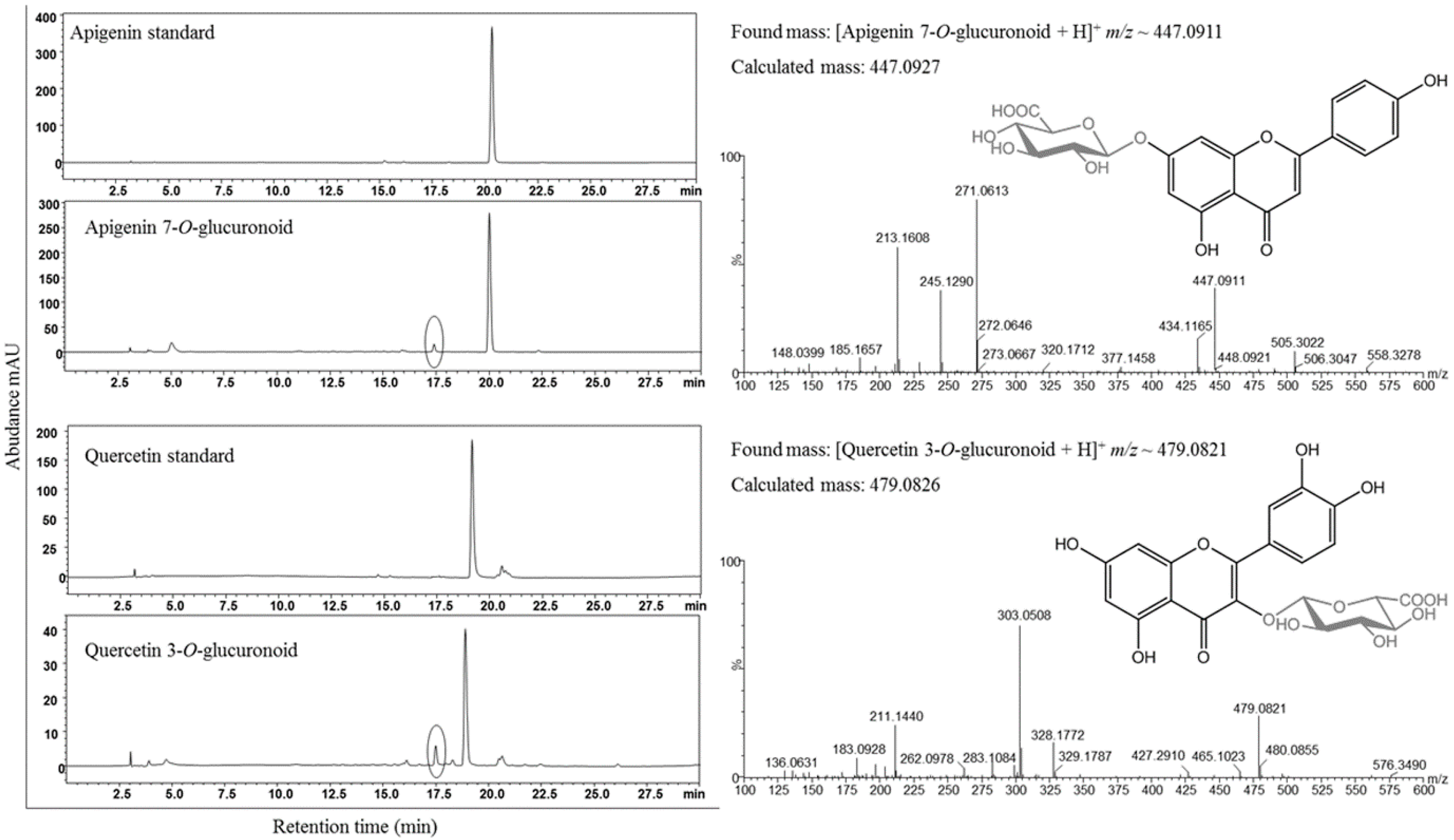
3.3. Synthesis of Glucuronoids in Recombinant Strains
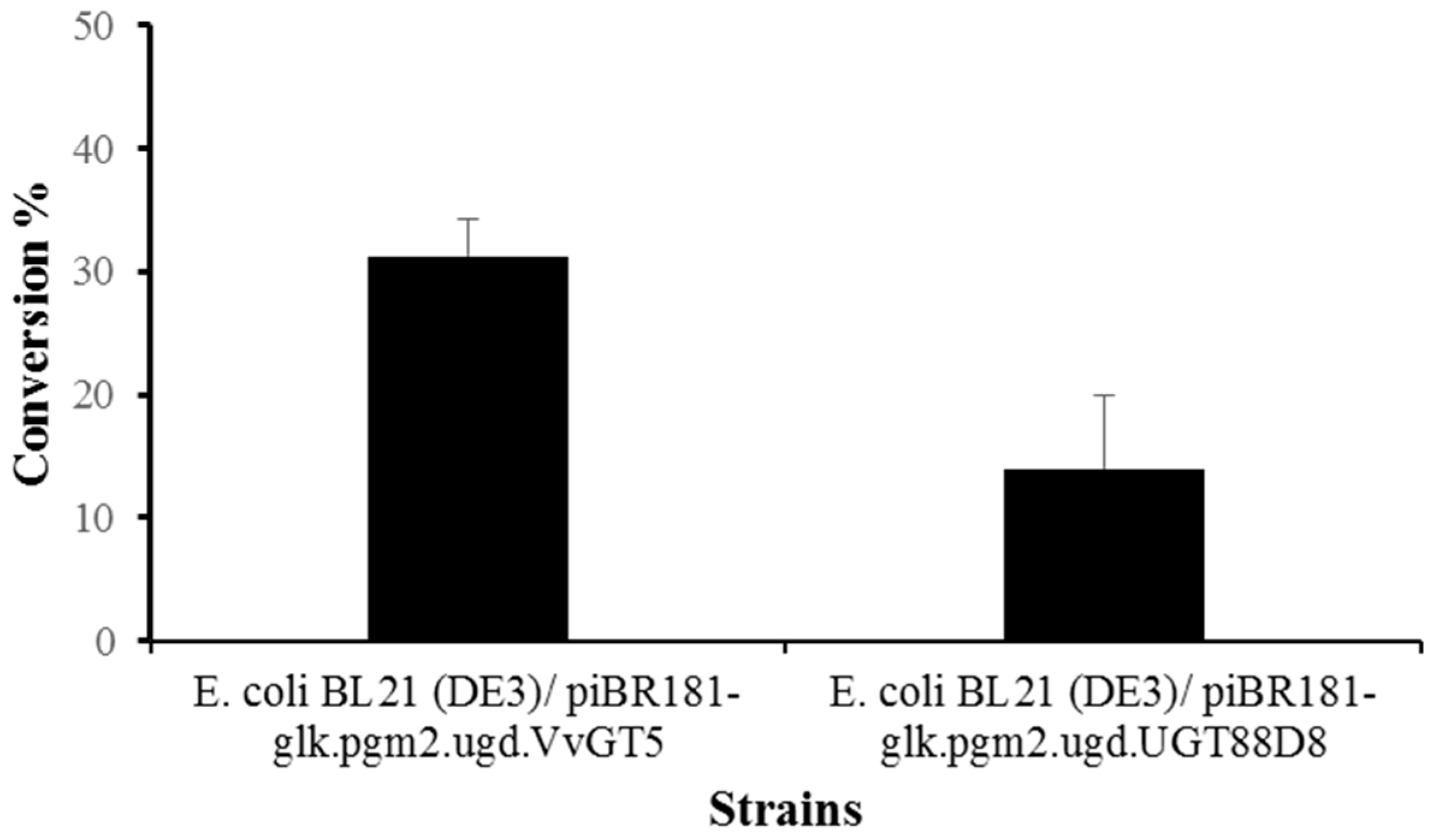
3.4. Enhanced Synthesis of Miquelianin and Scutellarin A
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Falcone Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 222. [Google Scholar] [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed]
- Airoldi, C.; La Ferla, B.; D Orazio, G.; Ciaramelli, C.; Palmioli, A. Flavonoids in the treatment of Alzheimer’s and other neurodegenerative diseases. Curr. Med. Chem. 2018, 25, 3228–3246. [Google Scholar] [CrossRef] [PubMed]
- Winkel-Shirley, B. Flavonoid biosynthesis, A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef]
- Song, M.C.; Kim, E.J.; Kim, E.; Rathwell, K.; Nam, S.J.; Yoon, Y.J. Microbial biosynthesis of medicinally important plant secondary metabolites. Nat. Prod. Rep. 2014, 31, 1497–1509. [Google Scholar] [CrossRef]
- Veitch, N.C.; Grayer, R.J. Flavonoids and their glycosides, including anthocyanins. Nat. Prod. Rep. 2008, 25, 555–611. [Google Scholar] [CrossRef] [PubMed]
- Min, Y.S.; Yim, S.H.; Bai, K.L.; Choi, H.J.; Jeong, J.H.; Song, H.J.; Park, S.H.; Ham, I.; Whang, W.K.; Sohn, U.D. The effects of apigenin-7-O-β-d-glucuronopyranoside on reflux oesophagitis and fastritis in rats. Auton. Autacoid Pharmacol. 2005, 25, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Gupta, S. Apigenin: A promising molecule for cancer prevention. Pharm. Res. 2010, 27, 962–978. [Google Scholar] [CrossRef]
- Gurung, R.B.; Pandey, R.P.; Sohng, J.K. Role of apigenin in cancer prevention. In Apigenin and Naringenin: Natural Sources, Pharmacology and Role in Cancer Prevention; Stacks, N.M., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2016. [Google Scholar]
- Cermak, R.; Wolffiram, S. The potential of flavonoids to influence drug metabolism and pharmacokinetics by local gastrointestinal mechanisms. Curr. Drug Metab. 2006, 7, 729–744. [Google Scholar] [CrossRef] [PubMed]
- Theodosiou, E.; Loutrari, H.; Stamatis, H.; Roussos, C.; Kolisis, F.N. Biocatalytic synthesis and antitumor activities of novel silybin acylated derivatives with dicarboxylic acids. New Biotechnol. 2011, 28, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Desmet, T.; Soetaert, W.; Bojarova, P.; Kren, V.; Dijkhuizen, L.; Eastwick-Field, V.; Schiller, A. Enzymatic glycosylation of small molecules: Challenging substrates required tailored catalysts. Chemistry 2012, 18, 10786–10801. [Google Scholar] [CrossRef] [PubMed]
- Makino, T.; Shimizu, R.; Kanemaru, M.; Suzuki, Y.; Moriwaki, M.; Mizukami, H. Enzymatically modified isoquercitrin, alpha-oligoglucosyl quercetin 3-O-glucoside, is absorbed more easily than other quercetin glycosides or aglycone after oral administration in rats. Biol. Pharm. Bull. 2009, 32, 2034–2040. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.P.; Parajuli, P.; Koffas, M.A.G.; Sohng, J.K. Microbial production of natural and non-natural flavonoids: Pathway engineering, directed evolution and systems/synthetic biology. Biotechnol. Adv. 2016, 34, 634–662. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Lv, M.; Hu, J.; Huang, K.; Xu, H. Glycosylation and activities of natural products. Mini-Rev. Med. Chem. 2016, 16, 1013–1016. [Google Scholar] [CrossRef]
- King, C.D.; Rios, G.R.; Green, M.D.; Tephyl, T.R. UDP-glucuronosyltransferases. Curr. Drug Metab. 2000, 1, 143–161. [Google Scholar] [CrossRef]
- Sarma, P.S.; Ssastry, K.S. Glucuronic acid, a precursor of ascorbic acid in Aspergillus niger. Nature 1957, 179, 44–45. [Google Scholar]
- Yang, T.; Bar-Peled, Y.; Smith, J.A.; Glushka, J.; Bar-Peled, M. In-microbe formation of nucleotide sugars in engineered Escherichia coli. Anal. Biochem. 2012, 421, 691–698. [Google Scholar] [CrossRef]
- Wang, S. Structure, mechanism and engineering of plant natural product glycosyltransferases. FEBS Lett. 2009, 583, 3303–3309. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Chaudhary, A.K.; Park, J.W.; Yoon, Y.J.; Kim, B.G.; Sohng, J.K. Re-engineering genetic circuit for 2-deoxystreptamine (2-DOS) biosynthesis in Escherichia coli BL21 (DE3). Biotechnol. Lett. 2013, 35, 285–293. [Google Scholar] [CrossRef]
- Parajuli, P.; Pandey, R.P.; Pokhrel, A.R.; Ghimire, G.P.; Sohng, J.K. Enzymatic glycosylation of the topical antibiotic mupirocin. Glycoconj. J. 2014, 31, 563–572. [Google Scholar] [CrossRef]
- Siedler, S.; Bringer, S.; Blank, L.M.; Bott, M. Engineering yield and rate of reductive biotransformation in Escherichia coli by partial cyclization of the pentose phosphate pathway and PTS-independent glucose transport. Appl. Microbiol. Biotechnol. 2012, 93, 1459–1467. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Muzashvili, T.S.; Georgiev, M.I. Advancees in the biotechnological glycosylation of valuable flavonoids. Biotechnol. Adv. 2014, 32, 1145–1156. [Google Scholar] [CrossRef] [PubMed]
- Ono, E.; Ruike, M.; Iwashita, T.; Nomoto, K.; Fukui, Y. Co-pigmentation and flavonoid glycosyltransferases in blue veronica a persica flowers. Phytochemistry 2010, 71, 726–735. [Google Scholar] [CrossRef]
- Mizohata, E.; Okuda, T.; Hatanaka, S.; Nakayama, T.; Horikawa, M.; Nakayama, T.; Ono, E.; Inoue, T. Crystallization and preliminary X-ray chrystallographic analysis of UDP-glucuronic acid: Flavonol-3-O-glucuronosyltransferase (VvGT5) from the grapevine Vitis vinifera. Acta Crystallogr. Sect. F 2013, 69, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Simkhada, D.; Lee, H.C.; Sohng, J.K. Genetic engineering approach for the production of rhamnosyl and allosyl flavonoids from Escherichia coli. Biotechnol. Bioeng. 2010, 107, 154–162. [Google Scholar] [CrossRef]
- Malla, S.; Pandey, R.P.; Kim, B.G.; Sohng, J.K. Regiospecific modifications of naringenin for astragalin production in Escherichia coli. Biotechnol. Bioeng. 2013, 110, 2525–2535. [Google Scholar] [CrossRef]
- Kim, B.G.; Jung, N.R.; Joe, E.J.; Hur, H.G.; Lim, Y.; Chong, Y.; Ahn, J.H. Bacterial synthesis of a flavonoid deoxyaminosugar conjugate in Escherichia coli expressing a glycosyltransferase of Arabidopsis thaliana. ChemBioChem 2010, 11, 2389–2392. [Google Scholar] [CrossRef]
- Kim, B.G.; Sung, S.H.; Ahn, J.H. Biological synthesis of quercetin 3-O-N-acetylglucosamine conjugate using engineered Escherichia coli expressing UGT78D2. Appl. Microbiol. Biotechnol. 2012, 93, 2447–2453. [Google Scholar] [CrossRef]
- Pandey, R.P.; Malla, S.; Simkhada, D.; Kim, B.G.; Sohng, J.K. Production of 3-O-xylosyl quercetin in Escherichia coli. Appl. Microbiol. Biotechnol. 2013, 97, 1889–1901. [Google Scholar] [CrossRef]
- Han, S.H.; Kim, B.G.; Yoon, J.A.; Chong, Y.; Ahn, J.H. Synthesis of flavonoid O-pentosides by Escherichia coli through engineering of nucleotide sugar pathways and glycosyltransferase. Appl. Environ. Microbiol. 2014, 80, 2754–2762. [Google Scholar] [CrossRef]
- Thuan, N.H.; Pandey, R.P.; Thuy, T.T.; Park, J.W.; Sohng, J.K. Improvement of regio-specific production of myricetin-3-O-α-L-rhamnoside in engineered Escherichia coli. Appl. Biochem. Biotechnol. 2013, 171, 1956–1967. [Google Scholar] [CrossRef]
- Parajuli, P.; Pandey, R.P.; Trang, N.T.; Chaudhary, A.K.; Sohng, J.K. Synthetic sugar cassettes for the efficient production of flavonol glycosides in Escherichia coli. Microb. Cell Factories 2015, 14, 76. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, A.; Pandey, R.P.; Dhakal, D.; Parajuli, P.; Sohng, J.K. Biosynthesis of flavone C-glucosides in engineered Escherichia coli. Appl. Microbiol. Biotechnol. 2018, 102, 1251–1267. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.P.; Parajuli, P.; Sohng, J.K. Metabolic engineering of glycosylated polyketides biosynthesis. Emerg. Top. Life Sci. 2018. [Google Scholar] [CrossRef]
- Kim, S.Y.; Lee, H.R.; Park, K.S.; Kim, B.G.; Ahn, J.H. Metabolic engineering of Escherichia coli for the biosynthesis of flavonoid-O-glucuronides and flavonoid-O-galactoside. Appl. Microbiol. Biotechnol. 2015, 99, 2233–2242. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Oligonucleotide Sequences (5′-3′) | Total PCR Size | Restriction Sites |
|---|---|---|---|
| glk-F | TCTAGAATGGAAATTGTTGCGATTGACATCGGT | 984 bp | XbaI |
| glk-R | AAGCTTTTATTCAACTTCAGAATATTTGTTGGC | 984 bp | HindIII |
| pgm2-F | TCTAGAATGAGCTGGAGAACGAGCTATGAACGC | 1749 bp | XbaI |
| pgm2-R | AAGCTTTTACGAATTTGAGGTCGCTTTTACAAT | 1749 bp | HindIII |
| galU-F | TCTAGAATGGCTGCCATTAATACGAAAGTCAAA | 908 bp | XbaI |
| galU-R | AAGCTTTTACTTCTTAATGCCCATCTCTTCTTC | 908 bp | HindIII |
| ugd-F | TCTAGAATGAAAATCACCATTTCCGGTACTGGC | 1167 bp | XbaI |
| ugd-R | AAGCTTTTAGTCGCTGCCAAAGAGATCGCGGGT | 1167 bp | HindIII |
| UGT88D8-F | TCTAGAATGGAAGACACGATTATTCTGTATGCC | XbaI | |
| UGT88D8-R | AAGCTTTGATGAACTTATCCAGATCCACCACGC | HindIII | |
| VvGT5-F | TCTAGAATGACCACGACCGCCTCCTCAATGGAT | XbaI | |
| VvGT5-R | AAGCTTCGTGTCCAGCGGCAGTTTAGAGGTGGT | HindIII |
| Strains/Plasmids | Description | Source/Reference |
|---|---|---|
| Escherichia coli XL-1 Blue (MRF′) | General cloning host | Stratagene, CA USA |
| E. coli BL21 (DE3) | ompT hsdT hsdS (rB-mB-) gal (DE3) | Novagen, Madison, WI, USA |
| E. coli BL21 (DE3)/piBR181-UGT88D8 | E. coli BL21 (DE3) carrying piBR181-UGT88D | This study |
| E. coli BL21 (DE3)/piBR181-VvGT5 | E. coli BL21 (DE3) carrying piBR181-VvGT5 | This study |
| E. coli BL21 (DE3)/piBR181-glk.pgm2.ugd.UGT88D8 | E. coli BL21 (DE3) carrying piBR181.glk, pgm2, galU, ugd and UGT88D8 | This study |
| E. coli BL21 (DE3)/piBR181-glk.pgm2.ugd.VvGT5 | E. coli BL21 (DE3) carrying piBR181.glk, pgm2, galU, ugd, and VvGT5 | This study |
| Plasmid and Vectors | ||
| pGEM-T® easy vector | General cloning vector, T7, and SP6 promoters, f1 ori, Ampr | Promega, Madison, WI, USA |
| pIBR181 | Mono-cistronic vector | [21] |
| piBR181 piBR181-UGT88D8 | The piBR181 vector carrying UGT88D8, codon optimized flavonoid 3-O-glucuronosyltransferase from blue flowers of Veronica persica | This study |
| piBR181 piBR181-VvGT5 | The piBR181 vector carrying VvGT5, codon optimized flavonoid 7-O-glucuronosyltransferase from grapevine (Vitis vinifera) | This study |
| piBR181-glk.pgm2.galU.ugd. UGT88D8 | The piBR181 vector carrying glk from Zymomonas mobilis, pgm2 from Bacillus licheniformis, galU and ugd from E. coli K12 and UGT88D8 from blue flowers of Veronica persica | This study |
| piBR181-glk.pgm2.galU.ugd. VvGT5 | The piBR181 vector carrying glk from Zymomonas mobilis, pgm2 Bacillus licheniformis, galU and ugd from E. coli K12 and VvGT5 from grapevine (Vitis vinifera) | This study |
| pVWEx1-glf-glk | pVWEx1 carrying glk | [23] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pandey, R.P.; Jung, H.Y.; Parajuli, P.; Nguyen, T.H.T.; Bashyal, P.; Sohng, J.K. A Synthetic Approach for Biosynthesis of Miquelianin and Scutellarin A in Escherichia coli. Appl. Sci. 2019, 9, 215. https://doi.org/10.3390/app9020215
Pandey RP, Jung HY, Parajuli P, Nguyen THT, Bashyal P, Sohng JK. A Synthetic Approach for Biosynthesis of Miquelianin and Scutellarin A in Escherichia coli. Applied Sciences. 2019; 9(2):215. https://doi.org/10.3390/app9020215
Chicago/Turabian StylePandey, Ramesh Prasad, Ha Young Jung, Prakash Parajuli, Thi Huyen Trang Nguyen, Puspalata Bashyal, and Jae Kyung Sohng. 2019. "A Synthetic Approach for Biosynthesis of Miquelianin and Scutellarin A in Escherichia coli" Applied Sciences 9, no. 2: 215. https://doi.org/10.3390/app9020215
APA StylePandey, R. P., Jung, H. Y., Parajuli, P., Nguyen, T. H. T., Bashyal, P., & Sohng, J. K. (2019). A Synthetic Approach for Biosynthesis of Miquelianin and Scutellarin A in Escherichia coli. Applied Sciences, 9(2), 215. https://doi.org/10.3390/app9020215