15N Natural Abundance, Nitrogen and Carbon Pools in Soil-Sorghum System Amended with Natural and NH4+-Enriched Zeolitites

 ,
,  , and
, and
Abstract
:1. Introduction
- (1)
- Evaluate how the addition of NZ and NEZ affects N and C pool dynamics in the soil-plant system over one agronomic year;
- (2)
- Determine if the use of NZ or NEZ can allow a reduction in the application of chemical fertilizers to soil;
- (3)
- Determine if the use of NZ or NEZ have beneficial effects on crop yield.
2. Materials and Methods
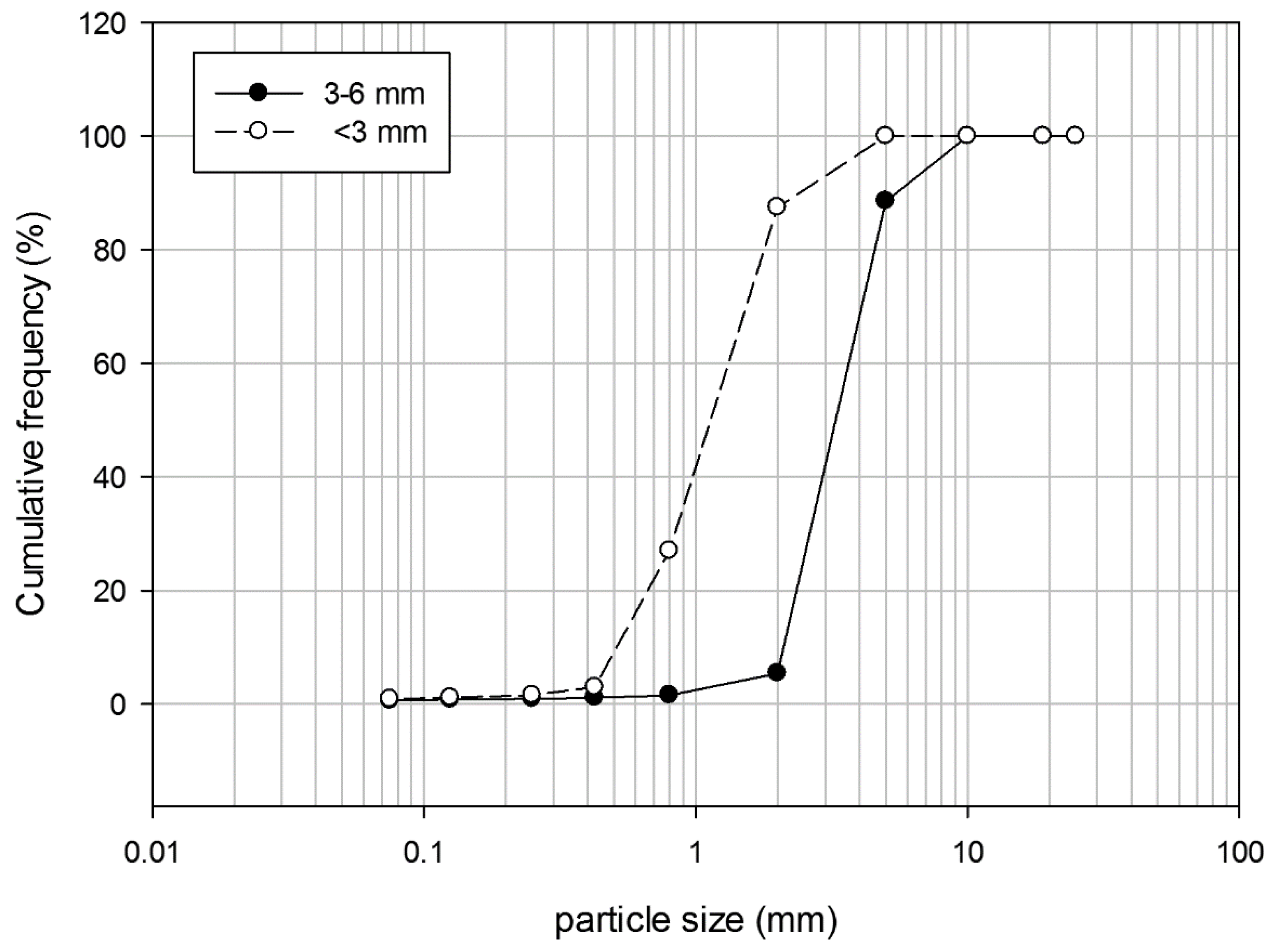
2.1. Zeolitites
2.2. Experimental Field Setting
2.3. Soil and Plant Sampling
2.4. Analytical Techniques
2.5. Statistical Analysis
3. Results
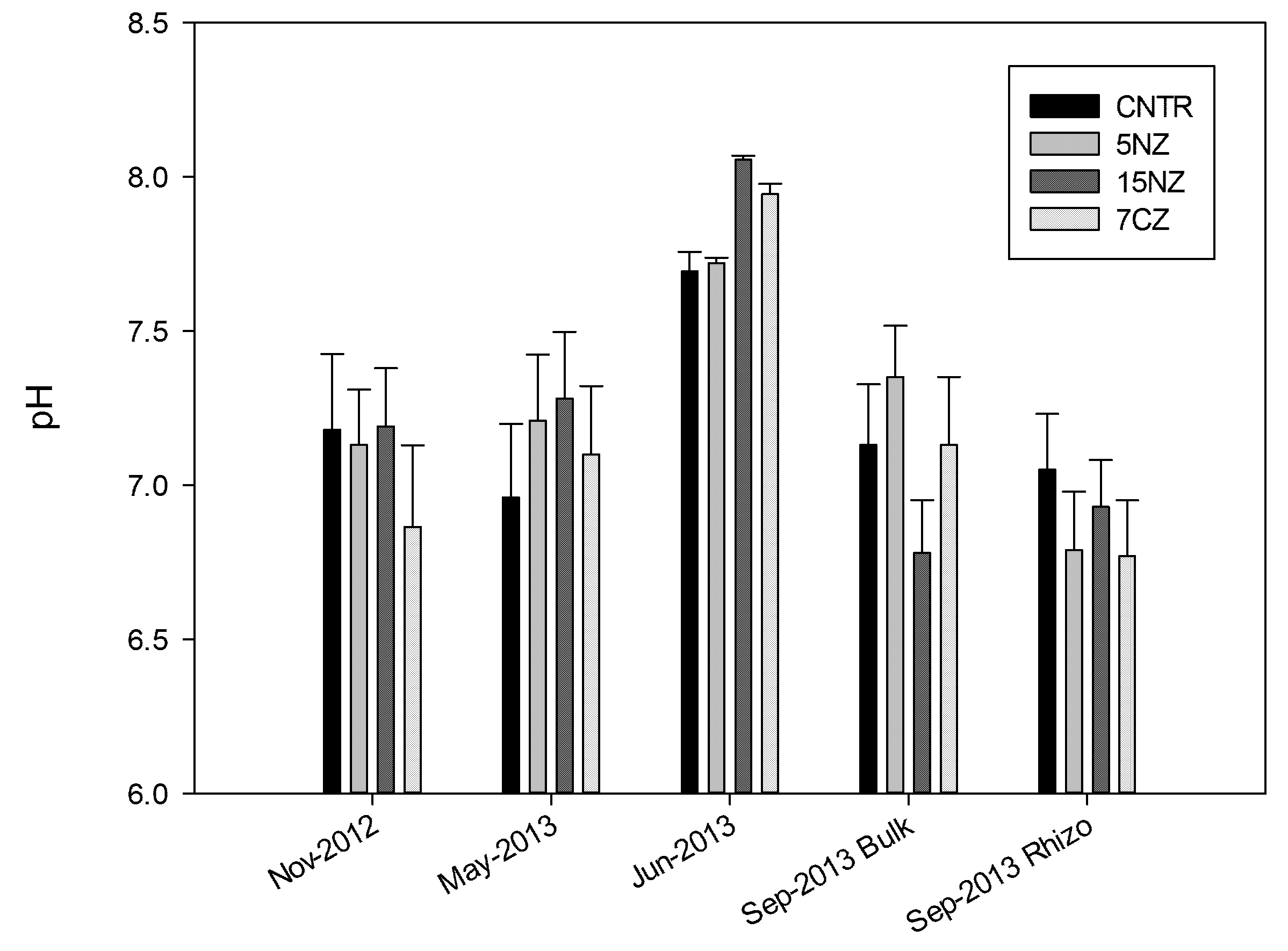
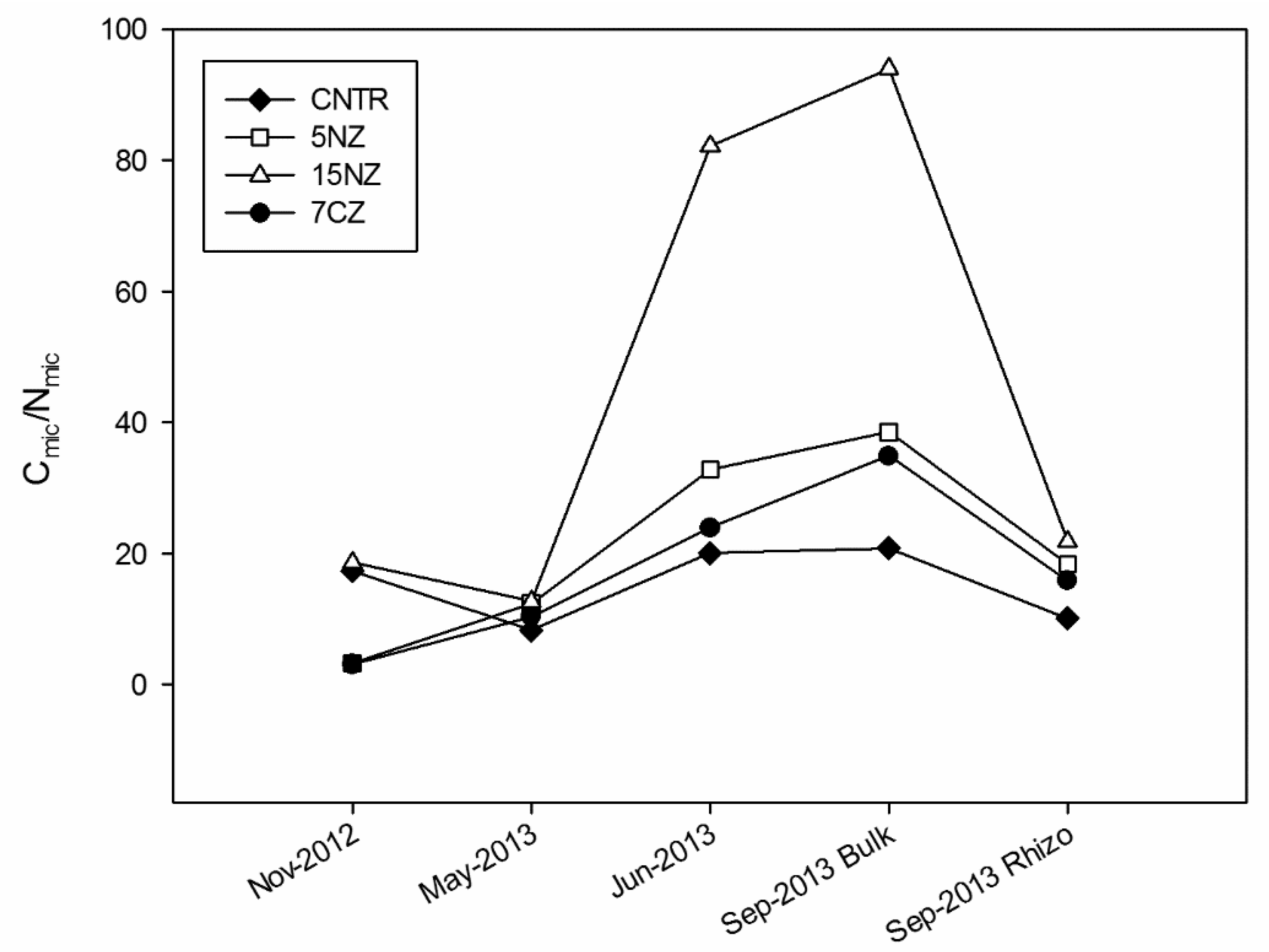
3.1. Nitrogen and Carbon Pools in Soil
3.2. Total N and C Content in Sorghum Plant Components
3.3. δ15N Natural Abundance in Soil and Sorghum Organs
3.4. Sorghum Yield
4. Discussion
4.1. NEZ Influence on Soil N and C Pools.
4.1.1. NEZ Effects on N Pool Size and Isotopic Signature
4.1.2. NEZ Specific Effects on Soil Microbial Biomass
4.1.3. NEZ Specific Effects on Fixed N-NH4+ Pool
4.2. NZ Influence on Soil N and C Pools
4.3. Zeolitite Influence on Rhizosphere
4.4. Plant-Zeolitite Interactions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Conway, G. The Double Green Revolution: Food for All in the 21st Century; Penguin: London, UK, 1997; p. 334. [Google Scholar]
- Smil, V. Nitrogen in crop production: An account of global flows. Glob. Biogeochem. Cycles 1999, 13, 647–662. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D.; Fargione, J.; Wolff, B.; D’Antonio, C.; Dobson, A.; Howarth, R.; Schindler, D.; Schlesinger, W.H.; Simberloff, D.; Swackhamer, D. Forecasting agriculturally driven global environmental change. Science 2001, 292, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Oenema, O.; Kros, H.; Vries, W. Approaches and uncertainties in nutrient budgets: Implications for nutrient management and environmental policies. Eur. J. Agron. 2003, 20, 3–16. [Google Scholar] [CrossRef]
- Kroeze, C.; Aerts, R.; Breemen, N.; Dam, D.; Hoek, K.; Hofschreuder, P.; Hoosbeek, M.; Klein, J.; Kros, H.; Oene, H.; et al. Uncertainties in the fate of nitrogen I: An overview of sources of uncertainty illustrated with a Dutch case study. Nutr. Cycl. Agroecosyst. 2003, 66, 43–69. [Google Scholar] [CrossRef]
- Passaglia, E. Zeoliti Naturali, Zeolititi e Loro Applicazioni; Arvan: Padova, Italy, 2008; p. 104. [Google Scholar]
- Wu, Q.; Chi, D.; Xia, G.; Chen, T.; Sun, Y.; Song, Y. Effects of Zeolite on Drought Resistance and Water—Nitrogen Use Efficiency in Paddy Rice. J. Irrig. Drain. Eng. 2019, 145, 4019024. [Google Scholar] [CrossRef]
- Wu, Q.; Chen, T.; Chi, D.; Xia, G.; Sun, Y.; Song, Y. Increasing nitrogen use efficiency with lower nitrogen application frequencies using zeolite in rice paddy fields. Int. Agrophys. 2019, 33, 263–269. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Li, Y. Zeolite as slow release fertilizer on spinach yields and quality in a greenhouse test. J. Plant Nutr. 2013, 36, 1496–1505. [Google Scholar] [CrossRef]
- Malferrari, D.; Laurora, A.; Brigatti, F.; Coltorti, M.; Di Giuseppe, D.; Faccini, B.; Vezzalini, M. Open-field experimentation of an innovative and integrated zeolitite cycle: Project definition and material characterization. Rend. Lincei-Sci. Fis. 2013, 24, 141–150. [Google Scholar] [CrossRef]
- Colombani, N.; Di Giuseppe, D.; Faccini, B.; Ferretti, G.; Mastrocicco, M.; Coltorti, M. Estimated water saving in an agricultural field amended with natural zeolites. Environ. Process. 2016, 3, 617–628. [Google Scholar] [CrossRef]
- Di Giuseppe, D.; Ibáñez Insa, J.; Melchiorre, M.; Coltorti, M. On the potential effect of micronized zeolites on seed germination: A prospective study. Period. Mineral. 2018, 85, 179–183. [Google Scholar]
- Ferretti, G.; Di Giuseppe, D.; Faccini, B.; Coltorti, M. Mitigation of sodium risk in a sandy agricultural soil by the use of natural zeolites. Environ. Monit. Assess. 2018, 190, 646. [Google Scholar] [CrossRef] [PubMed]
- Reháková, M.; Čuvanová, S.; Dzivák, M.; Rimár, J.; Gaval’ová, Z. Agricultural and agrochemical uses of natural zeolite of the clinoptilolite type. Curr. Opin. Solid State Mater. Sci. 2004, 8, 397–404. [Google Scholar] [CrossRef]
- Ferretti, G.; Keiblinger, K.M.; Di Giuseppe, D.; Faccini, B.; Colombani, N.; Zechmeister-Boltenstern, S.; Coltorti, M.; Mastrocicco, M. Short-term response of soil microbial biomass to different chabazite zeolite amendments. Pedosphere 2018, 28, 277–287. [Google Scholar]
- Bernardi, A.C.C.; Oliviera, P.P.A.; Monte, M.B.M.; Souza-Barros, F. Brazilian sedimentary zeolite use in agriculture. Microporous Mesoporous Mater. 2013, 167, 16–21. [Google Scholar] [CrossRef] [Green Version]
- Colombani, N.; Mastrocicco, M.; Di Giuseppe, D.; Faccini, B.; Coltorti, M. Variation of the hydraulic properties and solute transport mechanisms in a silty-clay soil amended with natural zeolites. Catena 2014, 123, 195–204. [Google Scholar] [CrossRef]
- Ferretti, G.; Keiblinger, K.M.; Zimmermann, M.; Di Giuseppe, D.; Faccini, B.; Colombani, N.; Mastrocicco, M.; Zechmeister-Boltenstern, S.; Mentler, A. High resolution short-term investigation of soil CO2, N2O, NOx and NH3 emissions after different chabazite zeolite amendments. Appl. Soil Ecol. 2017, 119, 138–144. [Google Scholar] [CrossRef]
- Ferretti, G.; Di Giuseppe, D.; Natali, C.; Faccini, B.; Bianchini, G.; Coltorti, M. CN elemental and isotopic investigation in agricultural soils: Insights on the effects of zeolitite amendments. Chem. Der Erde 2017, 77, 45–52. [Google Scholar] [CrossRef]
- Gholamhoseini, M.; Ghalavand, A.; Khodaei-Joghan, A.; Dolatabadian, A.; Zakikhani, H.; Farmanbar, E. Zeolite-amended cattle manure effects on sunflower yield, seed quality, water use efficiency and nutrient leaching. Soil Tillage Res. 2013, 126, 193–202. [Google Scholar] [CrossRef]
- Di Giuseppe, D.; Ferretti, G.; Faccini, B.; Blasi, E.; Passeri, N.; Bianchini, G.; Coltorti, M. Is it possible to cultivate corn in a sustainable way using a quarry waste? Period. Mineral. 2016, 85, 179–183. [Google Scholar]
- Arcara, P.G.; Gamba, C.; Bidini, D.; Marchetti, R. The effect of urea and pig slurry fertilization on denitrification, direct nitrous oxide emission, volatile fatty acids, water-soluble carbon and anthrone-reactive carbon in maize-cropped soil from the Po plain (Modena, Italy). Biol. Fertil. Soils 1999, 29, 270–276. [Google Scholar] [CrossRef]
- Faccini, B.; Di Giuseppe, D.; Malferrari, D.; Coltorti, M.; Abbondanzi, F.; Campisi, T.; Laurora, A.; Passaglia, E. Ammonium-exchanged zeolitite preparation for agricultural uses: From laboratory tests to large-scale application in ZeoLIFE project prototype. Period. Mineral. 2015, 84, 303–321. [Google Scholar]
- Faccini, B.; Di Giuseppe, D.; Ferretti, G.; Coltorti, M.; Colombani, N.; Mastrocicco, M. Natural and NH4+-enriched zeolitite amendment effects on nitrate leaching from a reclaimed agricultural soil (Ferrara Province, Italy). Nutr. Cycl. Agroecosyst. 2018, 110, 327–341. [Google Scholar] [CrossRef]
- Mastrocicco, M.; Colombani, N.; Di Giuseppe, D.; Faccini, B.; Coltorti, M. Contribution of the subsurface drainage system in changing the nitrogen speciation of an agricultural soil located in a complex marsh environment (Ferrara, Italy). Agric. Water Manag. 2013, 119, 144–153. [Google Scholar] [CrossRef]
- Di Giuseppe, D.; Faccini, B.; Mastrocicco, M.; Colombani, N.; Coltorti, M. Reclamation influence and background geochemistry of neutral saline soils in the Po River Delta Plain (Northern Italy). Environ. Earth Sci. 2014, 72, 2457–2473. [Google Scholar] [CrossRef]
- IUSS Working Group. World Reference Base for Soil Resources. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports, No. 106; FAO: Rome, Italy, 2007. [Google Scholar]
- Colombani, N.; Di Giuseppe, D.; Faccini, B.; Ferretti, G.; Mastrocicco, M.; Coltorti, M. Inferring the interconnections between surface water bodies, tile-drains and an unconfined aquifer-aquitard system: A case study. J. Hydrol. 2016, 537, 86–95. [Google Scholar] [CrossRef]
- Orsini, L.; Rèmy, J.C. Utilization du chlorure de cobaltihexammine pour la déterminantion simultanée de la capacité d′èchange et des bases échangeables des sols. Sci. Sol. 1976, 4, 269–275. [Google Scholar]
- Ciesielsky, H.; Sterckeman, T. Determination of cation exchange capacity and exchangeable cations in soils by means of cobalt heamine trichloride. Effects of experimental conditions. Agronomie 1997, 17, 1–7. [Google Scholar]
- Mariotti, A. Natural N-15 abundance measurements and atmospheric nitrogen standards. Nature 1984, 311, 251–252. [Google Scholar] [CrossRef]
- Silva, J.A.; Bremner, J.M. Determination and isotope-ratio analysis of different forms of nitrogen in soils: 5 Fixed ammonium. Soil Sci. Soc. Am. J. 1966, 30, 587–594. [Google Scholar] [CrossRef]
- Ciavatta, C.; Govi, M.; Vittori Antisari, L.; Sequi, P. Characterization of humified compounds by extraction and fractionation on solid polyvinylpyrrolidone. J. Chromatogr. 1990, 509, 41–146. [Google Scholar] [CrossRef]
- Sequi, P.; De Nobili, M.; Leita, L.; Cercignani, G. A new index of humification. Agrochimica 1986, 30, 175–179. [Google Scholar]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Ozbahce, A.; Tari, A.F.; Gönülal, E.; Simsekli, N.; Padem, H. The effect of zeolite applications on yield components and nutrient uptake of common bean under water stress. Arch. Agron. Soil Sci. 2015, 61, 615–626. [Google Scholar] [CrossRef]
- Barbarick, K.A.; Pirela, H.J. Agronomic and horticultural uses of zeolites: A review. In Zeo-Agriculture: Use of Natural Zeolites in Agriculture and Aquaculture; Pond, W.G., Mumpton, F.A., Eds.; Westview Press: Boulder, CO, USA, 1984; pp. 93–103. [Google Scholar]
- Eslami, M.; Khorassani, R.; Coltorti, M.; Malferrari, D.; Faccini, B.; Ferretti, G.; Di Giuseppe, D.; Fotovat, A.; Halajnia, A. Leaching behaviour of a sandy soil amended with natural and NH4+ and K+ saturated clinoptilolite and chabazite. Arch. Agron. Soil Sci. 2018, 64, 1142–1151. [Google Scholar] [CrossRef]
- Dwairi, I.M. Evaluation of Jordanian zeolite tuff as a controlled slow-release fertilizer for NH4+. Environ. Geol. 1998, 34, 1–4. [Google Scholar] [CrossRef]
- Dittert, K.; Georges, T.; Sattelmacher, B. Nitrogen turnover in soil after application of animal manure and slurry as studied by the stable isotope 15N: A review. Z. Pflanzenernähr. Bodenkd. 1998, 161, 453–463. [Google Scholar] [CrossRef]
- Högberg, P. Tansley review No. 95 15N natural abundance in soil-plant systems. N. Phytol. 1997, 137, 179–203. [Google Scholar]
- Schmidt, O.; Ostle, N.J. Tracing nitrogen derived from slurry in earthworms using 15N/14N stable isotope ratios at natural abundances. Appl. Soil Ecol. 1999, 12, 7–13. [Google Scholar] [CrossRef]
- Sang-Sun, L.; Woo-Jung, C.; Jyn-Hyeob, K.; Jae-Woon, J.; Scott, X.C.; Han-Yong, K.; Kwang-Sik, Y.; Soo-Myung, C. Nitrogen and carbon isotope responses of Chinese cabbage and chrysanthemum to the application of liquid pig manure. Plant Soil 2007, 295, 67–77. [Google Scholar]
- Kitayama, K.; Iwamoto, K. Patterns of natural 15N abundance in the leaf-to-soil continuum of tropical rain forests differing in N availability on Mount Kinabalu, Borneo. Plant Soil 2001, 229, 203–212. [Google Scholar] [CrossRef]
- Gualtieri, A.F.; Passaglia, E. Rietveld structure refinement of NH4-exchanged natural chabazite. Eur. J. Mineral. 2006, 18, 351–359. [Google Scholar] [CrossRef]
- Hedström, A. Ion exchange of ammonium in zeolites: A literature review. J. Environ. Eng. 2001, 127, 673–681. [Google Scholar] [CrossRef]
- Widiastuti, N.; Wu, H.; Ang, H.M.; Zhang, D. Removal of ammonium from greywater using natural zeolite. Desalination 2011, 277, 15–23. [Google Scholar] [CrossRef]
- Malekian, R.; Abedi-Koupai, J.; Eslamian, S.S. Influences of clinoptilolite and surfactant-modified clinoptilolite zeolite on nitrate leaching and plant growth. J. Hazard. Mater. 2011, 185, 970–976. [Google Scholar] [CrossRef]
- Azam, F.; Malik, K.A.; Hussain, F. Microbial biomass and mineralization-immobilization of nitrogen in some agricultural soils. Biol. Fertil. Soils 1986, 2, 157–163. [Google Scholar] [CrossRef]
- Ito, O.; Watanabe, I. Immobilization, mineralization and availability to rice plants of nitrogen derived from heterotrophic nitrogen fixation in flooded soil. Soil Sci. Plant Nutr. 1981, 27, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Schulten, H.R.; Schnitzer, M. The chemistry of soil organic nitrogen: A review. Biol. Fertil. Soils 1998, 26, 1–15. [Google Scholar] [CrossRef]
- Meurant, G. Soil Organic Matter and Its Role in Crop Production; Elsevier Science: Amsterdam, The Netherlands, 1973; p. 634. [Google Scholar]
- Scherer, H.W.; Werner, W. Significance of soil microorganisms for the mobilization of nonexchangeable ammonium. Biol. Fertil. Soils 1996, 22, 248–251. [Google Scholar] [CrossRef]
- Martens, D.A.; Bremner, J.M. Urea hydrolysis in soils: Factors influencing the effectiveness of phenylphosphorodiamidate as a retardant. Soil Biol. Biochem. 1984, 16, 515–519. [Google Scholar] [CrossRef]
- Li, C.; Fan, X.; Mengel, K. Turnover of interlayer ammonium in loess-derived soil grown with winter wheat in the Shaanxi Province of China. Biol. Fertil. Soils 1990, 9, 211–214. [Google Scholar] [CrossRef]
- Marzadori, C.; Vittori Antisari, L.; Gioacchini, P.; Sequi, P. Turnover of interlayer ammonium in soil cropped with sugar beet. Biol. Fertil. Soils 1994, 18, 27–31. [Google Scholar] [CrossRef]
- Hodge, A.; Robinson, D.; Fitter, A. Are microorganisms more effective than plants at competing for Nitrogen? Trends Plant Sci. 2000, 5, 304–308. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Vries, F.T.; Hoffland, E.; Eekeren, N.; Brussaard, L.; Bloem, J. Fungal/bacterial ratios in grasslands with contrasting nitrogen management. Soil Biol. Biochem. 2006, 38, 2092–2103. [Google Scholar] [CrossRef] [Green Version]
- Strickland, M.S.; Rousk, J. Considering fungal:bacterial dominance in soils—Methods, controls, and ecosystem implications. Soil Biol. Biochem. 2010, 42, 1385–1395. [Google Scholar] [CrossRef]
- Merckx, R.; Van Ginkel, J.H.; Sinnaeve, J.; Cremers, A. Plant induced changes in the rhizosphere of maize and wheat. I. Production and turnover of root-derived material in the rhizosphere of maize and wheat. Plant Soil 1986, 96, 85–93. [Google Scholar] [CrossRef]
- Liljeroth, E.; Van Veen, J.A.; Miller, H.J. Assimilate translocation to the rhizosphere of two wheat cultivars and subsequent utilization by rhizosphere microorganisms at two soil nitrogen levels. Soil Biol. Biochem. 1990, 22, 1015–1021. [Google Scholar] [CrossRef]
- Capasso, S.; Salvestrini, S.; Coppola, E.; Buondonno, A.; Colella, C. Sorption of humic acid on zeolitic tuff: A preliminary investigation. Appl. Clay Sci. 2005, 28, 159–165. [Google Scholar] [CrossRef]
- Söderström, B.; Hedlund, K.; Jackson, L.E.; Kätterer, T.; Lugato, E.; Thomensen, I.K.; Jørgensen, H.B. What are the effects of agricultural management on soil organic carbon (SOC) stocks? Environ. Evid. 2014, 3, 2. [Google Scholar] [CrossRef] [Green Version]
- Evans, D.R. Physiological mechanism influencing plant nitrogen isotope composition. Trends Plant Sci. 2001, 6, 121–126. [Google Scholar] [CrossRef]
- Evans, D.R.; Bloom, A.J.; Sukrapanna, S.S.; Ehleringer, J.R. Nitrogen isotope composition of tomato (Lycopersicum Esculentum Mill. cv. T-5) grown under ammonium or nitrate nutrition. Plant Cell Environ. 1996, 19, 1317–1323. [Google Scholar] [CrossRef]
- Erskine, P.D.; Bergstrom, D.M.; Schmidt, S.; Stewart, G.R.; Tweedie, C.E.; Shaw, J.D. Subantarctic Macquarie Island ± a model ecosystem for studying animal-derived nitrogen sources using 15N natural abundance. Oecologia 1998, 117, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Hulton, B.Z.; Sigman, D.M.; Schuur, E.A.G.; Edin, L.O. A climate-driven switch in plant nitrogen acquisition within tropical forest communities. Proc. Natl. Acad. Sci. USA 2007, 114, 8902–8906. [Google Scholar] [CrossRef] [PubMed]
- Kahmen, A.; Wanek, W.; Buchmann, N. Foliar d15N values characterize soil N cycling and reflect nitrate or ammonium preference of plants along a temperate grassland gradient. Oecologia 2008, 156, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.J.; Ro, H.M.; Hobbie, E.A. Patterns of natural 15N in soils and plants from chemically and organically fertilized uplands. Soil Biol. Biochem. 2003, 35, 1493–1500. [Google Scholar] [CrossRef]
- Bateman, A.S.; Kelly, S.D.; Jickells, T.D. Nitrogen Isotope Relationships between Crops and Fertilizer: Implications for Using Nitrogen Isotope Analysis as an Indicator of Agricultural Regime. J. Agric. Food Chem. 2005, 53, 5760–5765. [Google Scholar] [CrossRef]
- Yun, S.I.; Ro, H.M.; Choi, W.J.; Chang, S.X. Interactive effects of N fertilizer source and timing of fertilization leave specific N isotopic signatures in Chinese cabbage and soil. Soil Biol. Biochem. 2006, 38, 1682–1689. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
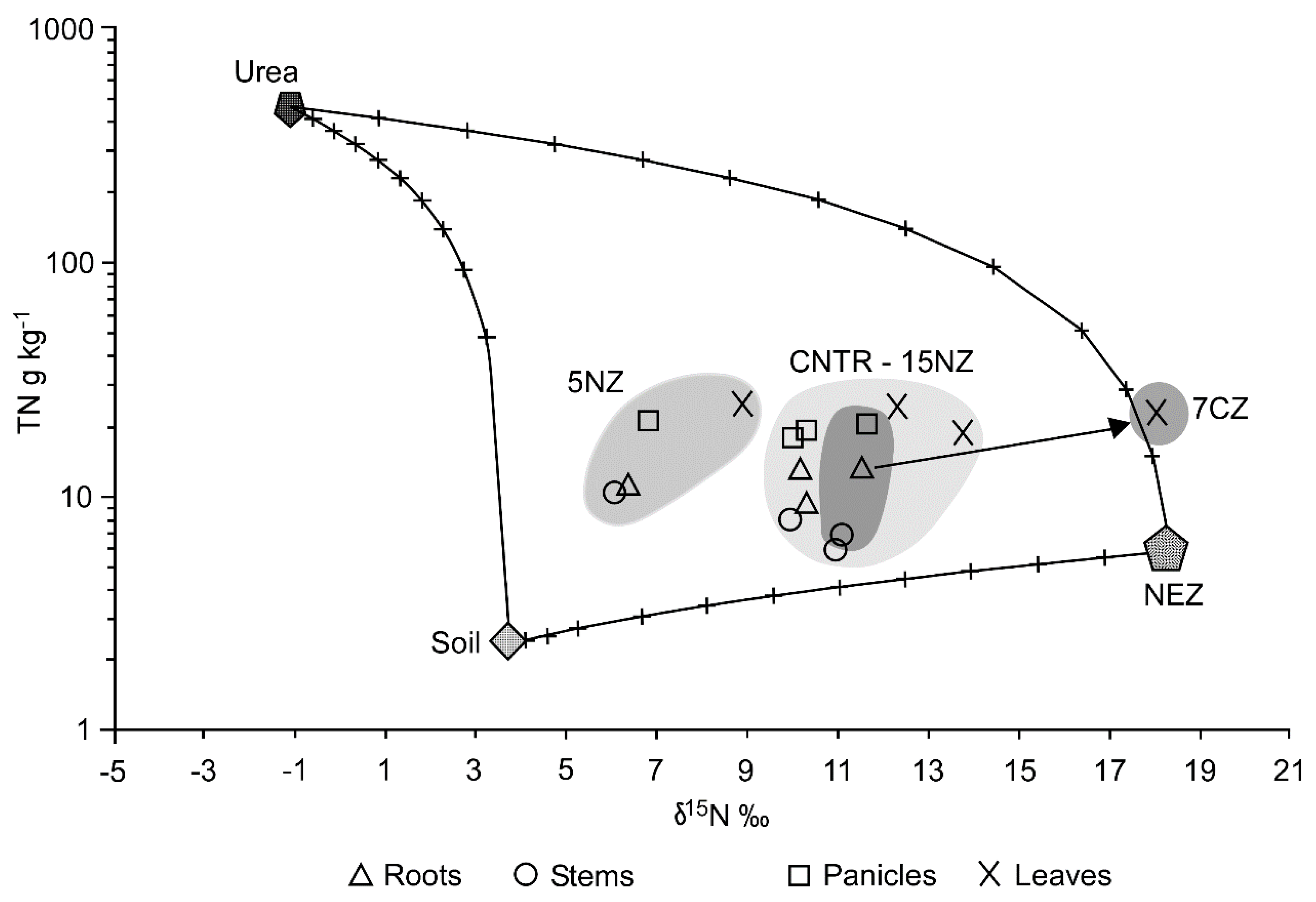{kind=link}
| TN | g·kg−1 | 5.84 (0.90) |
| Fix N-NH4+ | mg·kg−1 | 343 (21) |
| Exch N-NH4+ | mg·kg−1 | 4884 (330) |
| N-NO3− | mg·kg−1 | 148 (7.4) |
| Nmic | mg·kg−1 | 876 (152) |
| TOC | g·kg−1 | 10.3 (0.1) |
| Cmic | mg·kg−1 | 117 (1) |
| C/N | 1.78 | |
| Cmic/Nmic | 0.13 |
| Ca2+ | K+ | Mg2+ | Na+ | CEC | ||
|---|---|---|---|---|---|---|
| Time | Treatment | cmol·kg−1 | cmol·kg−1 | cmol·kg−1 | cmol·kg−1 | cmol·kg−1 |
| Sep-2013 Rhizo | CNTR | 18.3 (0.7) A | 8.3 (1.7) A | 14.4 (1.2) A | 1.8 (0.7) AB | 42.8 (4.0) A |
| 5NZ | 18.6 (0.6) A | 9.9 (0.8) A | 17.2 (0.9) B | 1.3 (0.1) A | 47.1 (1.9) A | |
| 15NZ | 17.7 (0.9) A | 10.4 (0.8) A | 17.9 (1.4) B | 1.6 (0.6) A | 47.5 (2.5) A | |
| 7CZ | 19.4 (1.1) A | 7.7 (3.8) A | 15.9 (0.6) AB | 3.0 (0.4) B | 46.0 (2.7) A | |
| Sep-2013 Bulk | CNTR | 19.9 (1.3) A | 6.1 (3.3) A | 16.8 (1.3) A | 2.5 (1.1) AB | 45.3 (2.3) A |
| 5NZ | 20.3 (0.01) AB | 4.9 (0.1) A | 21.9 (0.1) B | 2.0 (0.02) A | 49.1 (0.04) AB | |
| 15NZ | 23.0 (0.02) B | 5.9 (0.004) A | 19.5 (0.1) AB | 1.7 (0.1) A | 50.2 (0.2) B | |
| 7CZ | 22.0 (0.2) AB | 5.0 (0.1) A | 19.9 (0.2) B | 3.6 (0.04) B | 50.6 (0.6) B |
| HA | FA | TEC/TOC | HI | HD | ||
|---|---|---|---|---|---|---|
| Time | Treatment | g kg−1 | g kg−1 | % | % | |
| Nov-2012 | CNTR | 5.3 | 3.4 | 47.4 | 0.4 | 34.4 |
| 5NZ | 4.3 | 5.0 | 46.2 | 0.2 | 37.7 | |
| 15NZ | 3.9 | 3.7 | 41.4 | 0.5 | 27.8 | |
| 7CZ | 2.7 | 3.9 | 40.1 | 0.3 | 31.9 | |
| May-2013 | CNTR | 7.4 | 3.2 | 63.0 | 0.5 | 43.1 |
| 5NZ | 8.6 | 3.5 | 74.4 | 0.6 | 46.9 | |
| 15NZ | 11.4 | 3.6 | 87.3 | 0.3 | 68.2 | |
| 7CZ | 5.9 | 2.5 | 48.6 | 0.1 | 43.8 | |
| Jun-2013 | CNTR | 4.5 | 3.1 | 52.3 | 0.4 | 38.6 |
| 5NZ | 4.3 | 2.9 | 65.0 | 0.8 | 35.5 | |
| 15NZ | 3.8 | 3.1 | 50.6 | 0.3 | 39.2 | |
| 7CZ | 3.4 | 3.0 | 43.6 | 0.6 | 27.4 | |
| Sep-2013 Bulk | CNTR | 5.9 | 3.2 | 52.9 | 0.2 | 44.2 |
| 5NZ | 3.9 | 2.9 | 54.5 | 0.9 | 29.4 | |
| 15NZ | 8.6 | 3.3 | 70.7 | 0.4 | 51.3 | |
| 7CZ | 5.9 | 3.5 | 57.9 | 0.4 | 41.2 | |
| Sep-2013 Rhizo | CNTR | 8.1 | 3.1 | 58.1 | 0.3 | 43.4 |
| 5NZ | 6.9 | 3.2 | 48.5 | 0.4 | 34.7 | |
| 15NZ | 9.7 | 3.2 | 49.2 | 0.2 | 41.0 | |
| 7CZ | 7.1 | 3.3 | 48.8 | 0.3 | 37.0 |
| TN δ15N vs air | Fix δ15N vs air | ||
|---|---|---|---|
| Time | Treatment | ‰ | ‰ |
| Nov-2012 | CNTR | 3.86 (0.15) A | 6.00 (0.17) B |
| 5NZ | 3.83 (0.22) A | 5.28 (0.45) AB | |
| 15NZ | 3.38 (0.19) A | 4.55 (0.31) A | |
| 7CZ | 7.46 (0.53) B | 8.27 (0.36) C | |
| May-2013 | CNTR | 3.75 (0.20) AB | 3.34 (0.15) A |
| 5NZ | 3.34 (0.06) A | 3.38 (0.30) A | |
| 15NZ | 3.31 (0.62) A | 3.84 (0.04) A | |
| 7CZ | 4.87 (0.17) B | 3.11 (0.12) A | |
| Jun-2013 | CNTR | 3.11 (0.28) A | 3.65 (0.43) A |
| 5NZ | 3.37 (0.10) A | 3.38 (0.17) A | |
| 15NZ | 3.32 (0.05) A | 3.46 (0.13) A | |
| 7CZ | 3.27 (0.17) A | 3.73 (0.19) A | |
| Sep-2013 Bulk | CNTR | 2.68 (0.11) A | 1.37 (0.11) A |
| 5NZ | 2.39 (0.09) A | 1.83 (0.23) AB | |
| 15NZ | 2.73 (0.14) A | 1.79 (0.03) AB | |
| 7CZ | 2.55 (0.29) A | 2.09 (0.16) B | |
| Sep-2013 Rhizo | CNTR | 4.22 (0.38) A | 2.47 (0.26) A |
| 5NZ | 4.17 (0.31) A | 2.81 (0.75) A | |
| 15NZ | 4.55 (1.11) A | 2.40 (0.46) A | |
| 7CZ | 4.45 (0.19) A | 2.52 (0.31) A | |
| NEZ | 18.4 (0.5) | 17.3 (0.4) | |
| Urea | −1.18 (0.06) | - | |
| Diammonium Phosphate | −0.89 (0.09) | - | |
| Treatment | * | * | |
| Time | * | * | |
| Treatment*Time | * | * |
| δ15N vs Air | ||
|---|---|---|
| Plant Organ | Treatment | ‰ |
| Roots | CNTR | 10.2 (1.4) AB |
| 5NZ | 6.33 (1.69) A | |
| 15NZ | 10.3 (2.8) AB | |
| 7CZ | 11.6 (2.7) B | |
| Stems | CNTR | 9.95 (0.81) B |
| 5NZ | 6.08 (1.56) A | |
| 15NZ | 11.1 (1.1) B | |
| 7CZ | 11.0 (1.5) B | |
| Leaves | CNTR | 12.3 (1.6) B |
| 5NZ | 8.88 (0.94) A | |
| 15NZ | 13.8 (2.2) BC | |
| 7CZ | 18.1 (1.7) C | |
| Panicles | CNTR | 10.3 (0.4) B |
| 5NZ | 6.82 (0.97) A | |
| 15NZ | 10.0 (1.8) B | |
| 7CZ | 11.7 (2.0) B |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferretti, G.; Faccini, B.; Vittori Antisari, L.; Di Giuseppe, D.; Coltorti, M. 15N Natural Abundance, Nitrogen and Carbon Pools in Soil-Sorghum System Amended with Natural and NH4+-Enriched Zeolitites. Appl. Sci. 2019, 9, 4524. https://doi.org/10.3390/app9214524
Ferretti G, Faccini B, Vittori Antisari L, Di Giuseppe D, Coltorti M. 15N Natural Abundance, Nitrogen and Carbon Pools in Soil-Sorghum System Amended with Natural and NH4+-Enriched Zeolitites. Applied Sciences. 2019; 9(21):4524. https://doi.org/10.3390/app9214524
Chicago/Turabian StyleFerretti, Giacomo, Barbara Faccini, Livia Vittori Antisari, Dario Di Giuseppe, and Massimo Coltorti. 2019. "15N Natural Abundance, Nitrogen and Carbon Pools in Soil-Sorghum System Amended with Natural and NH4+-Enriched Zeolitites" Applied Sciences 9, no. 21: 4524. https://doi.org/10.3390/app9214524
APA StyleFerretti, G., Faccini, B., Vittori Antisari, L., Di Giuseppe, D., & Coltorti, M. (2019). 15N Natural Abundance, Nitrogen and Carbon Pools in Soil-Sorghum System Amended with Natural and NH4+-Enriched Zeolitites. Applied Sciences, 9(21), 4524. https://doi.org/10.3390/app9214524