The Effect of Aging on the ERP Correlates of Feedback Processing in the Probabilistic Selection Task


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Materials
2.3. Procedure
2.4. EEG Recording and Analysis
2.5. Data Availability
3. Results
3.1. Behavioral Data
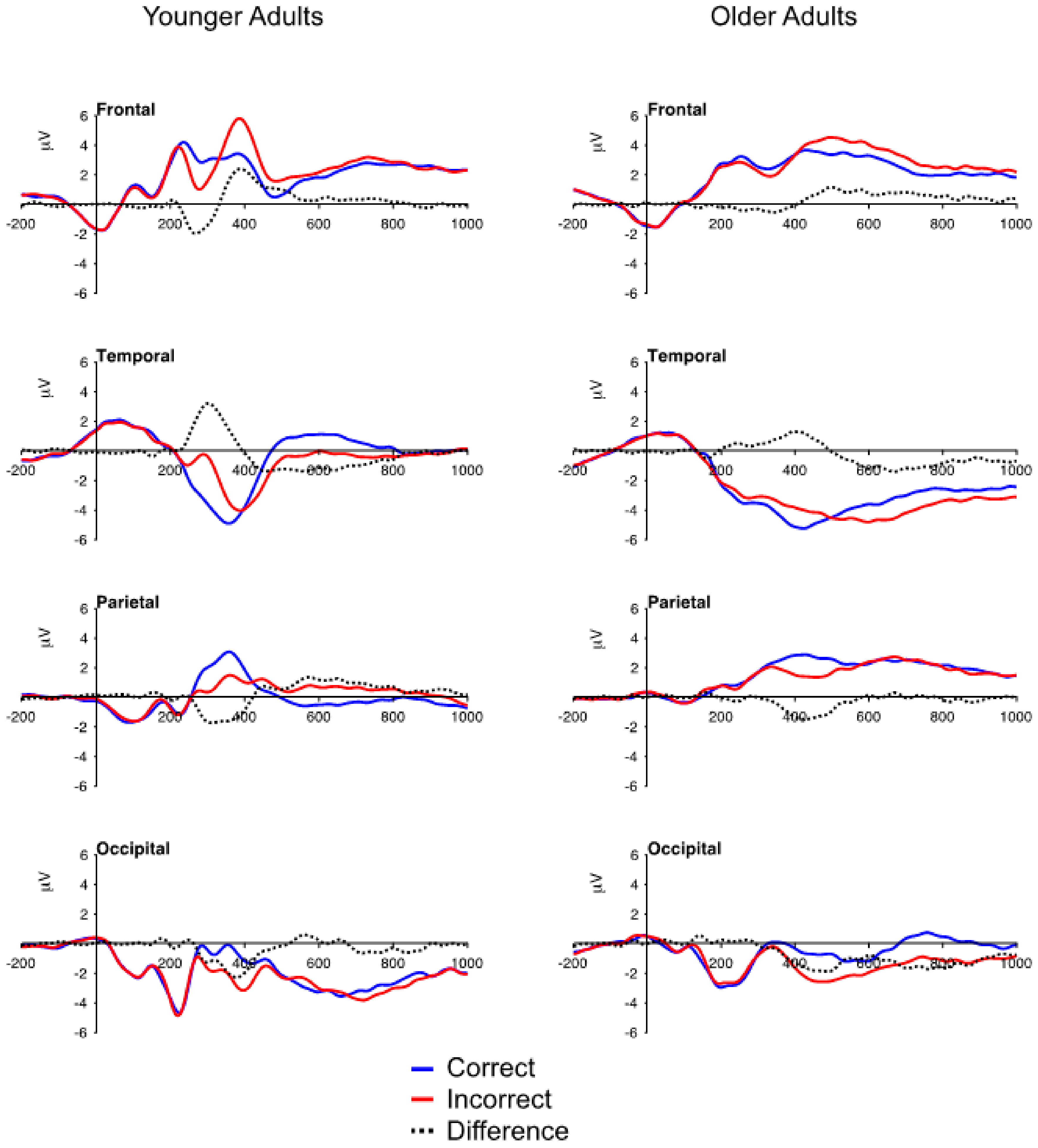
3.2. ERP Data: Mean Voltage
3.2.1. FN and Temporal Positivity
3.2.2. Frontal P3 and Occipital Negativity
3.2.3. Parietal P3
3.2.4. Aging and Correct Feedback
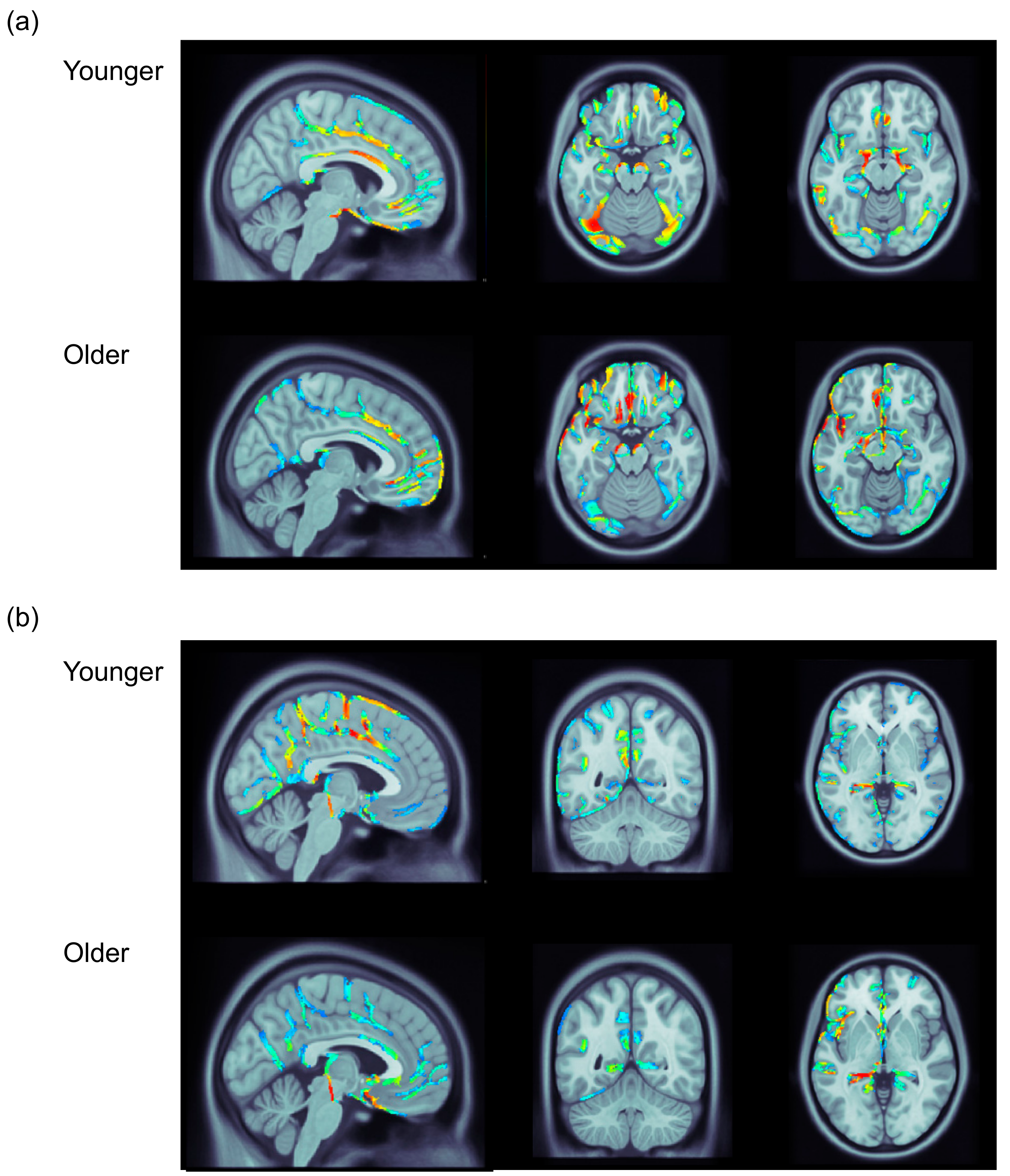
3.3. ERP Data: Distributed Source Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Frank, M.J.; Seeberger, L.C.; O’Reilly, R.C. By carrot or by stick: Cognitive reinforcement learning in parkinsonism. Science 2004, 306, 1940–1943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klucharav, V.; Hytönen, K.; Rijpkema, M.; Smidts, A.; Fernández, G. Reinforcement learning signal predicts social conformity. Neuron 2009, 61, 140–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knutson, B.; Samanez-Larkin, G.R.; Kuhnen, C.M. Gain and loss learning differentially contribute to life financial outcomes. PLoS ONE 2011, 6, e24390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gehring, W.J.; Willoughby, A.R. The medial frontal cortex and the rapid processing of monetary gains and losses. Science 2002, 295, 2279–2282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, M.W.; Anderson, J.R. Learning from experience: Event-related potential correlates of reward processing, neural adaptation, and behavioral choice. Neurosci. Biobeh. Rev. 2012, 36, 1870–1884. [Google Scholar] [CrossRef] [Green Version]
- West, R.; Tiernan, B.N.; Kieffaber, P.D.; Bailey, K.; Anderson, S. The effect of age on the neural correlates of feedback processing in a naturalistic gambling game. Psychophysiology 2012, 51, 734–745. [Google Scholar] [CrossRef] [PubMed]
- Yeung, N.; Sanfey, A.G. Independent coding of reward magnitude and valence in the human brain. J. Neurosci. 2004, 24, 6258–6264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellebaum, C.; Polezzi, D.; Daum, I. It is less than you expected: The feedback-related negativity reflects violations of reward magnitude expectations. Neuropsychologia 2010, 48, 3343–3350. [Google Scholar] [CrossRef] [PubMed]
- Goyer, J.P.; Woldorff, M.G.; Huettel, S.A. Rapid electrophysiological brain responses are influenced by both valence and magnitude of monetary rewards. J. Cogn. Neurosci. 2008, 20, 2058–2069. [Google Scholar] [CrossRef] [Green Version]
- West, R.; Bailey, K.; Tiernan, B.N.; Boonsuk, W.; Gilbert, S. The temporal dynamics of medial and lateral frontal neural activity related to proactive cognitive control. Neuropsychologia 2012, 50, 3450–3460. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhou, X. The P300 and reward valence, magnitude, and expectancy in outcome evaluation. Brain Res. 2009, 1286, 114–122. [Google Scholar] [CrossRef]
- Holroyd, C.B.; Pakzad-Vaezi, K.L.; Krigolson, O.E. The feedback correct-related positivity: Sensitivity of the event-related brain potential to unexpected positive feedback. Psychophysiology 2008, 45, 688–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, R.; Bailey, K.; Anderson, S.; Kieffaber, P.D. Beyond the FN: The neural correlates of feedback processing in a virtual blackjack game. Brain Cogn. 2014, 86, 104–115. [Google Scholar] [CrossRef] [PubMed]
- West, R.; Bailey, K.; Anderson, S. Transient and sustained ERP activity related to feedback processing in the probabilistic selection task. Int. J. Psychophysiol. 2018, 126, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hajcak, G.; Moser, J.S.; Holroyd, C.B.; Simons, R.F. It’s worse than you thought: The feedback negativity and violations of reward prediction in gambling tasks. Psychophysiology 2007, 44, 905–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van de Vijver, I.; van Driel, J.; Hillebrand, A.; Cohen, M.X. Interactions between frontal and posterior oscillatory dynamics support adjustments of stimulus processing during reinforcement learning. NeuroImage 2018, 18, 170–181. [Google Scholar] [CrossRef] [PubMed]
- Hewig, J.; Straube, T.; Trippe, R.H.; Kretschmer, N.; Hecht, H.; Coles, M.G.H.; Miltner, W.H.R. Decision-making under risk: An fMRI study. J. Cogn. Neurosci. 2008, 21, 1642–1652. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hairston, J.; Schrier, M.; Fan, J. Common and distinct networks underlying reward valence and processing stages: A meta-analysis of functional neuroimaging studies. Neurosci. Biobehav. Rev. 2011, 35, 1219–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eppinger, B.; Kray, J.; Mock, B.; Mecklinger, A. Better or worse than expected? Aging, learning, and the ERN. Neuropsychologia 2008, 46, 521–539. [Google Scholar] [CrossRef]
- Hämmerer, D.; Li, S.; Müller, V.; Lindenberger, U. Life span differences in electrophysiological correlates of monitoring gains and losses during probabilistic reinforcement learning. J. Cogn. Neurosci. 2010, 23, 579–592. [Google Scholar] [CrossRef] [Green Version]
- Nieuwenhuis, S.; Ridderinkhof, K.R.; Talsma, D.; Coles, M.G.H.; Holroyd, C.B.; Kok, A.; van der Molen, M.W. A computational account of altered error processing in older age: Dopamine and the error-related negativity. Cogn. Affect. Behav. Neurosci. 2002, 2, 19–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathewson, K.J.; Dyan, J.; Snyder, P.J.; Tays, W.J.; Segalowitz, S.J. Aging and electrocortical response to error feedback during a spatial learning task. Psychophysiology 2008, 45, 936–948. [Google Scholar] [CrossRef]
- Oldfield, R.C. The assessment and analysis of handedness. The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Wechsler, D. Wechsler Adult Intelligence Scale-Revised; The Psychological Corporation, Hartcourt Brace Janovich: San Antonio, TX, USA, 1981. [Google Scholar]
- Raven, J.C. Mill Hill Vocabulary Scale; H.K. Lewis: London, UK, 1965. [Google Scholar]
- Tadel, F.; Baillet, S.; Mosher, J.C.; Pantazis, D.; Leahy, R.M. Brainstorm: A user-friendly application for MEG/EEG analysis. Comput. Intell. Neurosci. 2011. [Google Scholar] [CrossRef]
- Salthouse, T.A. The processing-speed theory of adult age differences in cognition. Psychol. Rev. 1996, 103, 403–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, M.X.; Wilmes, K.A.; van de Vijver, I. Cortical electrophysiological network dynamics of feedback learning. Trends Cogn. Sci. 2011, 15, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.W.; Dennis, N.A.; Daseleer, S.M.; Fleck, M.S.; Cabeza, R. Qué PASA? The posterior-anterior shift in aging. Cereb. Cortex. 2008, 18, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Mann, S.L.; Hazlett, E.A.; Byne, W.; Hof, P.R.; Buchsbaum, M.S.; Cohen, B.H.; Goldstein, K.E.; Haznedar, M.M.; Mitsis, E.M.; Siever, L.J.; et al. Anterior and posterior cingulate cortex volume in healthy adults: Effects of aging and gender differences. Brain Res. 2011, 1401, 18–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carstensen, L.L.; Isaacowitz, D.M.; Charles, S.T. Taking time seriously: A theory of socioemotional selectivity. Am. Psychol. 1999, 54, 165–181. [Google Scholar] [CrossRef] [PubMed]
- Mather, M.; Carstensen, L.L. Aging and motivated cognition: The positivity effect in attention and memory. Trends Cogn. Sci. 2005, 9, 496–502. [Google Scholar] [CrossRef]
- Weniger, G.; Boucsein, K.; Irle, E. Impaired associative memory in temporal lobe epilepsy subjects after lesions of hippocampus, parahippocampal gyrus, and amygdala. Hippocampus 2004, 14, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Burgmans, S.; van Boxtel, M.P.J.; van den Berg, K.E.M.; Gronenschild, E.H.B.M.; Jabobs, H.I.L.; Jolles, J.; Uylings, H.B.M. The posterior parahippocampal gyrus is preferentially affected in age-related memory decline. Neurobiol. Aging 2011, 32, 1572–1578. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R. A triarchic model of P300 amplitude. Psychophysiology 1986, 23, 367–384. [Google Scholar] [CrossRef] [PubMed]
- Kok, A. On the utility of P3 amplitude as a measure of processing capacity. Psychophysiology 2001, 38, 557–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, D.; Kazmerski, V.; Fabiani, M. An overview of age-related changes in the scalp distribution of P3b. Electroencephalogr. Clin. Neurophysiol. 1997, 104, 98–513. [Google Scholar] [CrossRef]
- Polich, J. Meta-analysis of P300 normative aging studies. Psychophysiology 1996, 33, 334–353. [Google Scholar] [CrossRef] [PubMed]
- West, R.; Schwarb, H.; Johnson, B.N. The influence of age and individual differences in executive function on stimulus processing in the oddball task. Cortex 2010, 46, 550–563. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Younger | Older | ||
|---|---|---|---|
| Age | M | 19.56 | 71.42 |
| SD | 1.56 | 5.79 | |
| Education | M | 13.94 | 17.36 |
| SD | 0.95 | 2.76 | |
| Vocabulary | M | 14.62 | 23.14 |
| SD | 3.60 | 4.80 | |
| Digit symbol | M | 43.08 | 32.06 |
| SD | 6.01 | 7.30 | |
| Choose A | M | 0.66 | 0.60 |
| SD | 0.22 | 0.23 | |
| Avoid B | M | 0.63 | 0.54 |
| SD | 0.19 | 0.21 | |
| A | B | C | |||||
|---|---|---|---|---|---|---|---|
| Younger | Older | Younger | Older | Younger | Older | ||
| Block 1 | M | 0.70 | 0.66 | 0.65 | 0.57 | 0.51 | 0.57 |
| SD | 0.23 | 0.21 | 0.18 | 0.18 | 0.19 | 0.20 | |
| Block 2 | M | 0.72 | 0.72 | 0.68 | 0.65 | 0.55 | 0.60 |
| SD | 0.21 | 0.25 | 0.19 | 0.19 | 0.18 | 0.21 | |
| Block 3 | M | 0.78 | 0.78 | 0.68 | 0.61 | 0.55 | 0.60 |
| SD | 0.17 | 0.19 | 0.21 | 0.22 | 0.18 | 0.17 | |
| FN | Frontal P3 | Temp. pos. | Occ. neg. | Parietal P3 | ||
|---|---|---|---|---|---|---|
| Younger Adults | ||||||
| Correct feedback | M | 3.18 | 3.33 | −2.88 | −0.57 | 2.87 |
| SD | 2.52 | 2.44 | 2.57 | 2.56 | 2.31 | |
| Incorrect feedback | M | 1.40 | 5.24 | −0.54 | −2.43 | 1.29 |
| SD | 2.43 | 3.21 | 1.71 | 2.97 | 2.75 | |
| Older Adults | ||||||
| Correct feedback | M | 2.44 | 3.43 | −3.73 | −0.87 | 2.85 |
| SD | 3.38 | 3.43 | 3.68 | 3.80 | 2.76 | |
| Incorrect feedback | M | 2.02 | 4.43 | −3.16 | −2.39 | 1.43 |
| SD | 3.55 | 3.68 | 2.99 | 4.24 | 3.22 | |
| F | p | BF10 | |
|---|---|---|---|
| FN | 1.09 | 0.30 | 0.60 |
| Temporal positivity | 1.61 | 0.21 | 0.57 |
| Frontal | 0.05 | 0.83 | 0.38 |
| Occipital negativity | 0.23 | 0.64 | 0.37 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
West, R.; Huet, A. The Effect of Aging on the ERP Correlates of Feedback Processing in the Probabilistic Selection Task. Brain Sci. 2020, 10, 40. https://doi.org/10.3390/brainsci10010040
West R, Huet A. The Effect of Aging on the ERP Correlates of Feedback Processing in the Probabilistic Selection Task. Brain Sciences. 2020; 10(1):40. https://doi.org/10.3390/brainsci10010040
Chicago/Turabian StyleWest, Robert, and AnnMarie Huet. 2020. "The Effect of Aging on the ERP Correlates of Feedback Processing in the Probabilistic Selection Task" Brain Sciences 10, no. 1: 40. https://doi.org/10.3390/brainsci10010040
APA StyleWest, R., & Huet, A. (2020). The Effect of Aging on the ERP Correlates of Feedback Processing in the Probabilistic Selection Task. Brain Sciences, 10(1), 40. https://doi.org/10.3390/brainsci10010040