1. Introduction
Everyone with normal hearing sometimes experiences, when focusing on visual information, failure to hear surrounding sounds, for example, while reading we ignore background sounds and may not even hear someone talking to us. Conversely, if we focus our attention on auditory information, such as listening to a radio or another person, then, it is difficult to read. The question is whether such switches of attention are performed only at the cortical level or whether peripheral parts, such as the ear, are also involved.
If the ear itself plays an active or “conscious” part in switching attention, it is most likely to be through activity of the medial olivocochlear (MOC) system. MOC neurons descend from the brainstem to the cochleae of both ears, while the brainstem also receives input from auditory cortex. The function of this system is not entirely understood; probably its role relates to bilateral hearing tasks such as localization or detecting speech in noise [
1,
2]. Usefully, the activity of this system can be studied noninvasively by measuring changes in otoacoustic emissions (OAEs) in response to contralateral acoustic stimulation (CAS). With CAS, the amplitude of OAEs decreases, an effect called OAE suppression, OAE inhibition, or the MOC reflex (reviewed in [
3,
4,
5]).
This study concentrates on transiently evoked OAEs (TEOAEs), which are signals recorded up to 20 ms after onset of a stimulus, which is most often a click [
6]. Some early TEOAE papers claimed to find there was an effect of attention on TEOAEs even when contralateral stimulation was not involved (e.g., [
7]). These experiments were repeated by Michie et al. [
8] who found it was not possible to replicate these attentional effects. Some years later, several papers revived interest in the question of whether TEOAEs may in fact depend on attention but this time they involved TEOAE suppression. For example, de Boer and Thornton [
9] showed that performing a task did affect measured levels of suppression. Other studies also shown that there was a greater level of OAE suppression during active listening as compared with passive listening (e.g., [
10]). However, it is important to note that the suppression findings have been far from conclusive and even the authors acknowledge that the changes are very small and border on insignificance.
There are also encouraging results from animal studies. For example, Delano et al. [
11] showed a decrease in cochlear sensitivity during periods when chinchillas were attending to visual stimuli as compared with when they attended to auditory stimuli. Terreros et al. [
12] also showed that the MOC system of mice helped the animals ignore auditory distractors during visual attention. However, it is still unknown whether observations in animals mimic those in humans.
The rationale for the present study was to combine TEOAE measurements with event-related potentials (ERPs) to investigate the effect of switching attention from the visual to the auditory modality. Such an approach has the potential to give insight into whether there is a link between peripheral function (as assessed by TEOAEs) and conscious attention to a particular task (as reflected by the amplitude of the P3 cognitive wave in an ERP measurement). Until now, such an approach has not been used in studies comparing different modalities [
9,
13,
14,
15].
The purpose of this study was to investigate whether there was a difference in TEOAE suppression between tasks involving auditory attention on the one hand and visual attention on the other. ERPs were used as an objective measure of attention, while the OAE measurements were being made. TEOAE suppression was evaluated in different windows, and for its highest energy component. An additional perspective was to check whether task performance was affected by noise levels or number of rejected trials, since earlier studies have reported that, curiously, noise seemed to play an important role (e.g., [
9,
14]).
2. Materials and Methods
2.1. Experimental Design
ERPs were used to objectively gauge the subject’s state of attention, while TEOAEs in response to clicks were used to gauge inner ear function. As a test of descending neural activity, TEOAE levels were evaluated with and without CAS by broadband noise, a paradigm known to activate the MOC pathway.
The experiment was based on two attentional tasks, one auditory and the other visual. A schematic of the paradigm is shown in
Figure 1. There were four parts to the experiment. First, a reference measurement was made of TEOAEs without CAS, and without any tasks. Second, we introduced a task (randomly auditory or visual) while measuring TEOAEs with CAS. Third, we made a second reference measurement of TEOAEs without CAS. Fourth, we introduced a second task (again auditory or visual at random) while measuring TEOAEs with CAS. During both cognitive tasks, the subject received the same auditory and visual stimuli, but was instructed to direct attention to only one task modality and to ignore the other. The performance of tasks was evaluated by ERPs (which were synchronized with the auditory stimuli but not synchronized with the visual stimuli). In this way we expected to see a P3 wave (a marker of cognition) in ERPs during the auditory task and no P3 during the visual task when attention shifted to another modality.
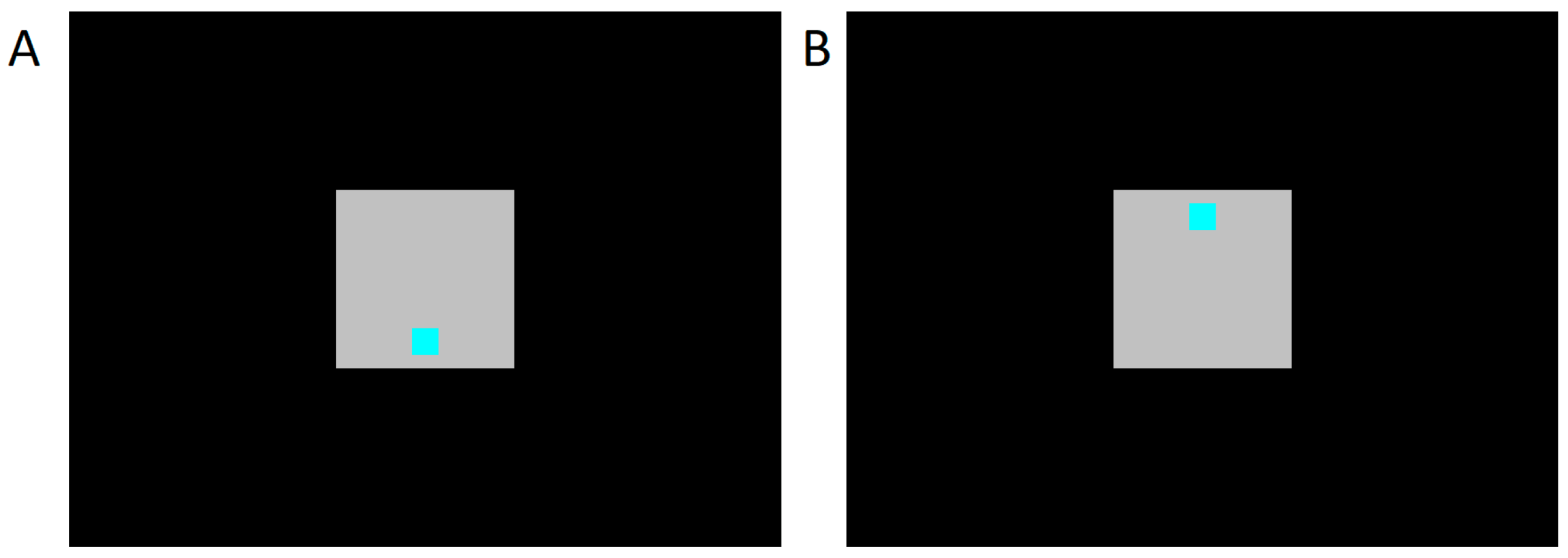
To manipulate attention, we asked subjects to attend to 5 dB Sound Pressure Level (SPL) decreases in the level of noise presented to the contralateral ear. In the auditory condition, the subject’s task was to tap a button when the noise level decreased (duration 1 s, every 4 ± 2 s). At the same time, the subject was presented with visual stimuli on a computer monitor 1.5 m in front of them. Stimuli were delivered according to a visual odd-ball paradigm (a sequence of standard stimuli randomly interrupted by an inverted deviant stimulus). Standard stimuli were squares shown at the bottom of the screen; deviant stimuli were the same squares but shown at the top (
Figure 2). There were 400 (80%) standard stimuli and 100 (20%) deviant stimuli in each recording session. The standard and deviant stimuli were both randomly presented for 100 ms every 2 s. The subjects were seated in such a way that the screen was about 1 m in front of their eyes.
The tasks were specifically designed to be easy in order to get a clear P3. In preliminary recordings when we used more difficult auditory tasks (1–3 dB SPL changes in CAS), we obtained significantly lower P3 magnitudes. This decrease made it harder to distinguish shifts from auditory to visual attention in ERP recordings. Therefore, we used 5 dB changes in CAS, which are quite easy to pick. The rates of correct responses were similar, with 97% for the auditory task and 96% for the visual task.
The two parts of the experiment were delivered to subjects in mixed order (in 11 cases the auditory part came first). The whole procedure lasted about 30 min. The duration of recording varied slightly depending on the number of artifacts rejected.
2.2. Participants
There were 20 normally hearing adults (age 25–40 years, 14 females) who participated in the study. All subjects had pure tone thresholds better than 25 dB HL at 0.5–8 kHz, normal middle ear function verified by 226 Hz tympanometry (tympanometric peak pressure values between −100 and +100 daPa and peak compensated static acoustic admittance values of 0.2–1.0 mmhos), and no known history of otologic disease. In all subjects, ipsilateral and contralateral middle ear acoustic reflex thresholds (ARTs) for broadband noise were above 80 dB SPL.
Measurements were initially collected in 24 subjects; however, 4 were later excluded from the analyses due to problems related to the experimental setup, i.e., OAE probe slippage, electrode slippage, or contamination of measurements by noise related to tiredness or movement during the experiment.
The subjects gave written informed consent prior to participation. The research procedures were approved by the Ethics Committee of the Institute of Physiology and Pathology of Hearing, Poland (approval no. IFPS:KB/09/2015).
2.3. TEOAE Procedures
Many OAE studies related to attention use custom-built systems and custom signal acquisition procedures (e.g., [
15,
16,
17]). It is not easy to replicate such experiments or to relate them to those made using other methods. Therefore, one of the underpinnings of this study was to perform experiments that were easy to replicate and which could be easily interpreted by researchers or clinicians using commercial diagnostic equipment. However, we did use different equipment (HearID, Mimosa Acoustics Inc., Champaign, IL, USA) than that used in a previous study (the ILO system, [
18]), since the HearID provides better signal-to-noise ratios (SNRs) (e.g., [
19]). Higher SNRs facilitate detection of smaller MOC effects (e.g., [
20]).
Using the HearID system, TEOAEs were recorded using 65 dB SPL clicks (linear mode). All recordings were performed in a 20 ms acquisition window. TEOAEs were collected for each part of the experiment (
Figure 1); each subaverage took about 8 min to collect. To minimize stimulus artifacts the initial 2.5 ms of all responses was windowed out automatically by the system. Responses were filtered in a 0.5–4.5 kHz range and broadband response levels were used for analysis. The quality of recordings was evaluated in terms of broadband SNR, calculated by subtracting the noise levels (in dB) from the response levels (in dB). For MOC reflex studies, SNRs need to be higher than for standard TEOAE studies (e.g., [
21]), and therefore, here, all analyzed recordings (with or without CAS) were required to have an SNR of at least 12 dB, as opposed to the usual 3 or 6 dB.
Recordings were made with and without 60 dB SPL white noise delivered to the contralateral ear. Only the right ear of every subject was tested for TEOAEs. Contralateral noise was delivered to the left ear through an Interacoustics AC40 audiometer (Denmark); its level was 60 dB SPL with 5 dB SPL decreases as described earlier. The sequence of decreases in noise amplitude was controlled by Presentation software (version 16.4).
TEOAE suppression was calculated by two methods. First, by subtracting the response levels with contralateral stimulation from the levels without, i.e., the response level of TEOAEs measured while performing a task was subtracted from the average response level measured during reference measurements 1 and 2. The second method takes into account phase effects and is based on the percentage change in the time domain waveform [
22,
23]:
where
N is the number of samples,
aquiet is the amplitude of the TEOAE waveform measured without CAS (average of reference measurements 1 and 2), and
anoise is the amplitude of the TEOAE waveform measured with CAS (during a task).
2.4. Event-Related Potentials (ERP) Procedures
ERPs were used to objectively gauge the subject’s state of attention during TEOAE recordings. ERPs were obtained from EEG signals recorded by a 32-channel EEG system (Brain Products, Gilching, GmBH, Germany). During the EEG measurement, 24 recording electrodes were used, made up of 22 active electrodes, 1 reference electrode, and 1 ground electrode [
24]. Active electrodes were mounted in an EEG cap placed on the head of the subject. The positions of 20 of them accorded with the 10/20 standard, and two electrodes were placed on the mastoids (TP9, TP10). The reference electrode was at FCz and the ground electrode was at AFz. During EEG acquisition, the impedance was monitored and always kept below 10 kΩ for all electrodes. The sampling frequency for each channel was 1000 Hz.
After recording, the EEG signal was analyzed offline using Brain Vision Analyzer 2.2 software (Brain Products, GmBH, Gilching, Germany). In the first step, the signal was re-referenced against the signal recorded from the mastoids. It was also digitally high- and low-pass filtered within the range 0.3–30 Hz. Next, eye-blink artifacts were corrected by zeroing the activation curves of individual independent component analysis (ICA) components corresponding to eye blinks [
25,
26]. The EEG signal was manually inspected and all artifacts related to muscle activity were removed. The artifact-free EEG signals were used to calculate ERPs. The signal was cut to an epoch of 1200 ms, from 200 ms pre-stimulus to 1000 ms post-stimulus. Subsequently, the level of each epoch was corrected relative to the baseline. Each of the epochs processed in this way was, then, assessed again in terms of artifacts.
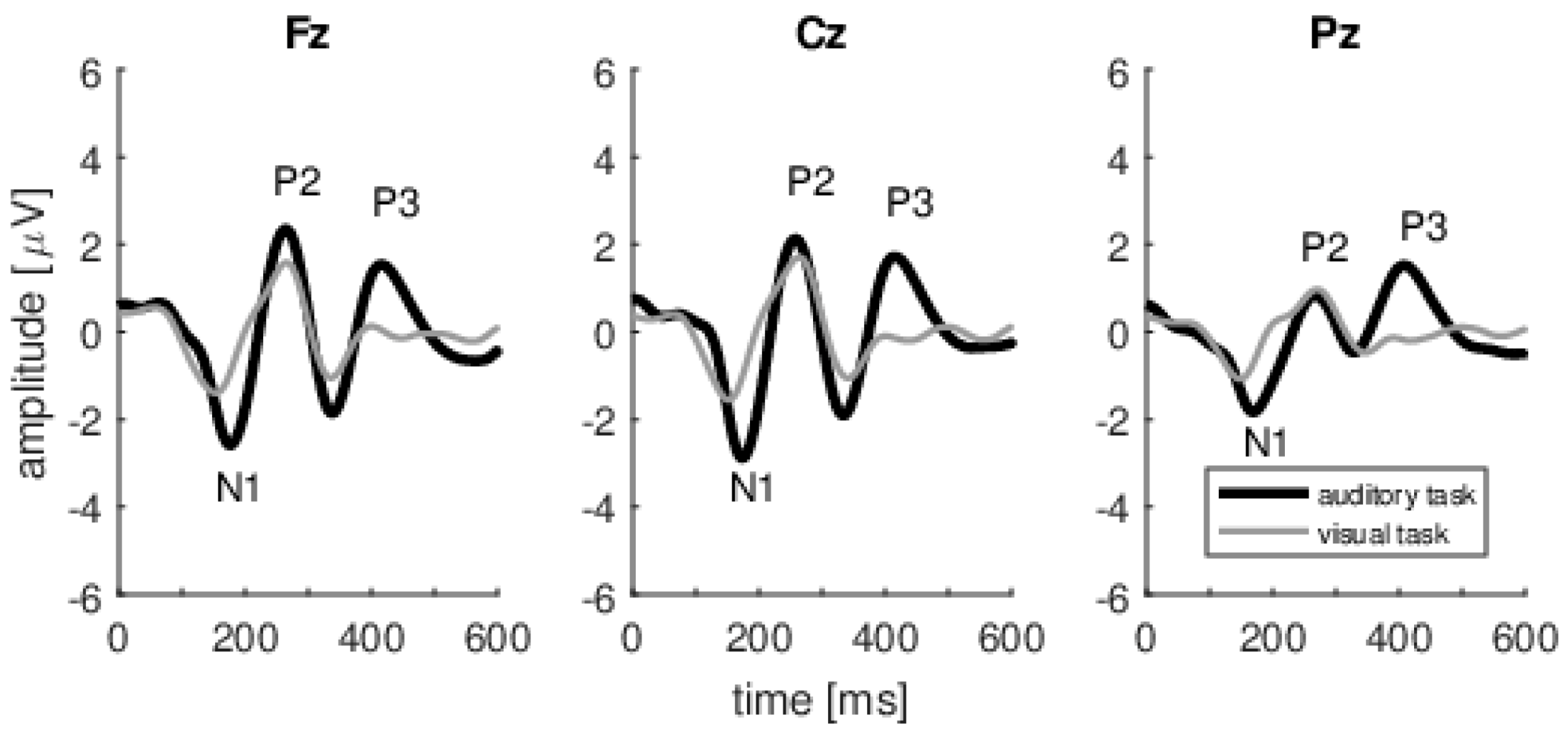
The final step was to obtain average ERPs from the previously prepared and artifact-free epochs. The two ERP averages were calculated for each subject, i.e., one from the EEG signal recorded during the auditory task, and a second during the visual task. Although we recorded the EEG signal from 22 active electrodes, ERPs were analyzed only for the following three positions along the midline: Fz, Cz, and Pz. In the present study, we simply wanted to identify the P3 wave and since P3 has frontal and parietal generators, we chose the midline electrodes.
The P3 wave in signals recorded during an auditory task was defined as the largest positive wave in the 250–600 ms window. The N1 and P2 waves in the auditory and visual tasks were defined as the maximum negative peak in the 90–170 ms and 170–250 ms windows respectively. The semi-automatic algorithm in Brain Analyzer software was used to detect the peaks in each of the listed waves for each subject.
2.5. Data Analysis: Matching Pursuit
For extracting the main components of the signal, a method based on the matching pursuit (MP) algorithm was used [
27]. The MP method allows a TEOAE signal to be decomposed into waveforms of defined frequency, latency, duration, and amplitude. The method fits the characteristics of TEOAE signals well, and is able to pick ”resonance modes” of TEOAEs (i.e., the most prominent waveform within a TEOAE signal), and show its suppression [
28,
29].
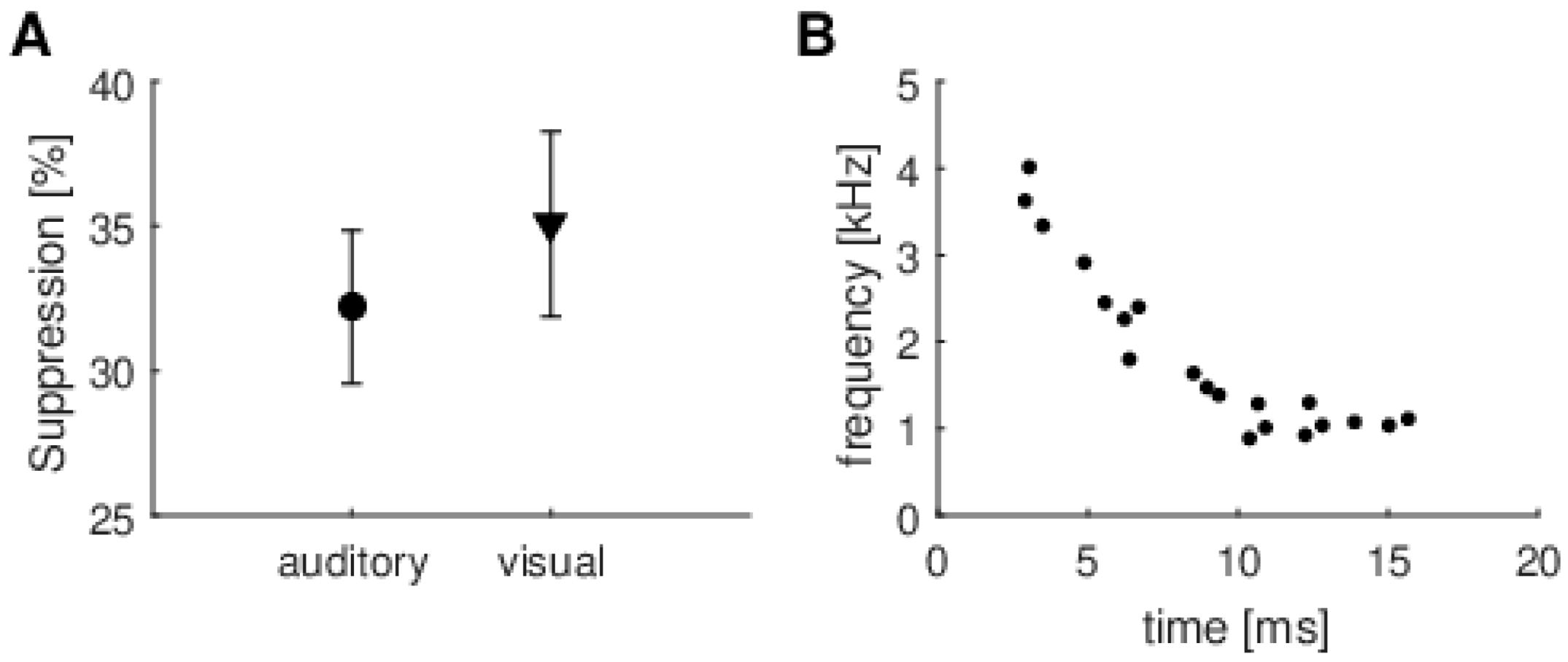
An example of applying the MP algorithm to calculate TEOAE suppression is shown in
Figure 3. In the top panel is a typical TEOAE signal (grey) with the highest energy resonant mode picked up by the MP method superimposed (black). In the middle panel, this resonant mode is shown with and without CAS. The bottom panel shows the time–frequency position of this resonant mode. The whole TEOAE signal is composed from several (usually about 20) resonant modes which span from around 4 kHz and 4 ms to around 1 kHz and 15 ms (e.g., [
30]).
2.6. Statistical Analysis
All analyses were made in Matlab (version 2020a, MathWorks, Natick, MA, USA). All datasets had normal distributions as indicated by a Shapiro–Wilk test. The statistical significance of the mean difference between groups was evaluated for all parameters using a t-test or ANOVA. For some analyses, Pearson correlations were calculated. In all analyses, a 95% confidence level (p < 0.05) was chosen as the criterion of significance.
4. Discussion
This study has investigated whether changing attention from the auditory to the visual modality has an effect on the MOC system (as gauged by the amount of TEOAE suppression produced by CAS). A change of attention was confirmed by a corresponding change in the P3 wave; it was present during auditory stimuli and absent during the visual task (while the same auditory stimuli were being delivered). However, despite a clear change in attention, we were unable to observe any change in the MOC system. At the same time, another interesting and novel observation was made, i.e., that the amplitude of the P3 cognitive potential increases at the same time as the level of TEOAE suppression actually decreases.
This study is a continuation of our previous study which focused only on visual attention [
18]. In that work, it was found that there was no effect of visual attention on TEOAE suppression. Some other studies have concluded that auditory attention may have a stronger influence on OAE suppression than does visual attention (e.g., [
9]). However, we were not able to see such an effect. This might be because of several factors. Here, we tried to control all parameters, and different modality sessions were identical with the only difference being the switch in modality. Therefore, there were no differences between TEOAEs during different modality tasks and there were no differences in noise levels or number of rejected trials. Some previous studies have claimed to have found such differences, but there is the possibility they were related not to attention but to the experimental setup [
9,
14]. In fact, Francis et al. [
16] gave examples of where there were decreases in physiological noise during attentional parts of the experiments, and they were associated with a reduction in subject motion, not by changes in MOC. Our study indirectly backs up this idea, that is, we do not observe changes in physiological noise when switching from a visual task to an auditory task (in fact we deliberately designed the experiment so that there would not be any change in movement between tasks). Our results are also in line with other recent studies, such as that of Dragicevic et al. [
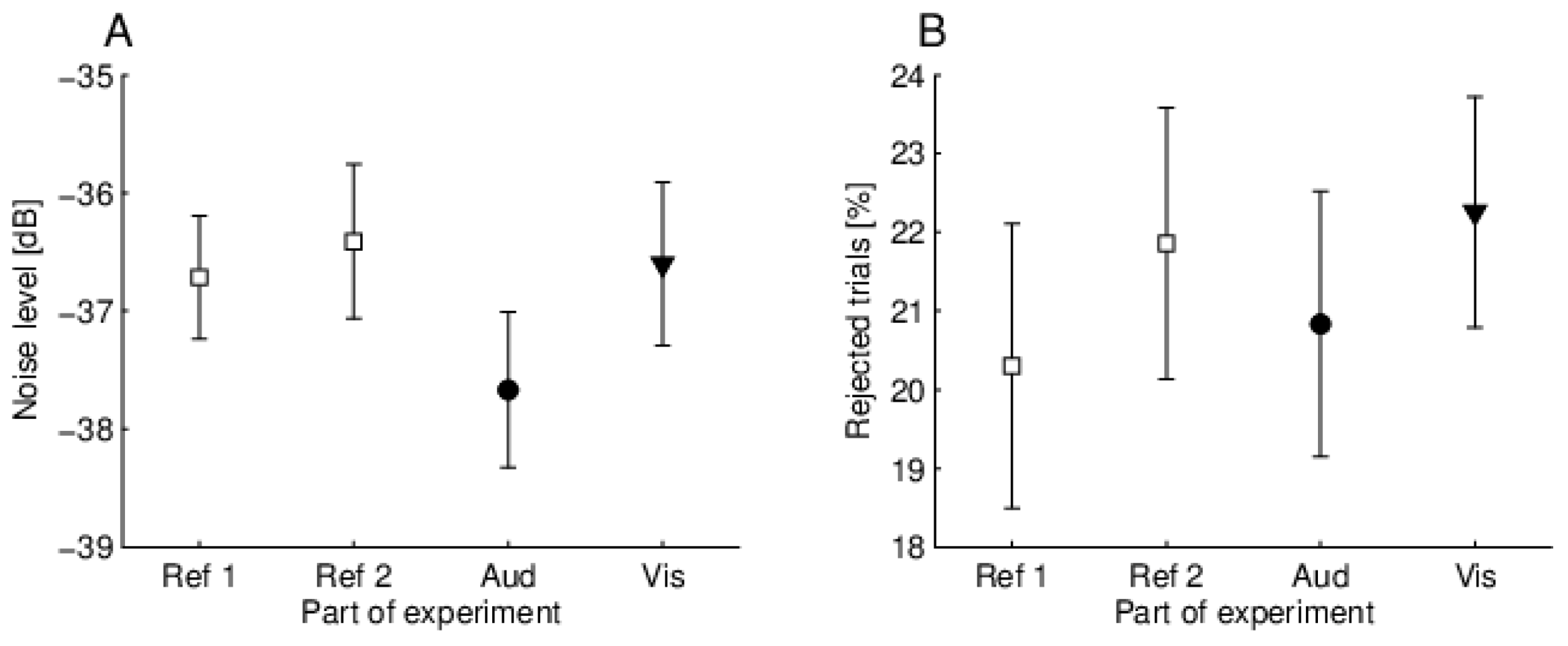
17], who found no change in the amplitude of OAEs during attention tasks, although they did find low frequency oscillations. Indeed, it seems that the attentional effects on OAEs, if they are present, are more subtle than just amplitude or broadband suppression changes. However, our experimental setup did not provide a way to explore effects such as low frequency oscillations, and therefore we were unable to directly confirm this interesting observation. We did try to look for other more subtle changes in the signal, for example, by examining specific components using the MP method, but again we failed to see any systematic changes.
It should be underlined that our study was based on easy tasks. It is possible that MOC effects are switched on only when the difficulty becomes higher. This needs to be resolved by dedicated experiments. Here, our intention was to examine the effect on OAE suppression when switching attention from one modality to the other, and such an approach gives the advantage of easily distinguishing ERP effects. Our starting assumption, based on the literature (e.g., [
9,
14,
15]), was that we would see some effect, at least on averaged data. The final results, which demonstrate no effect, might be a starting point for further experiments involving more difficult tasks.
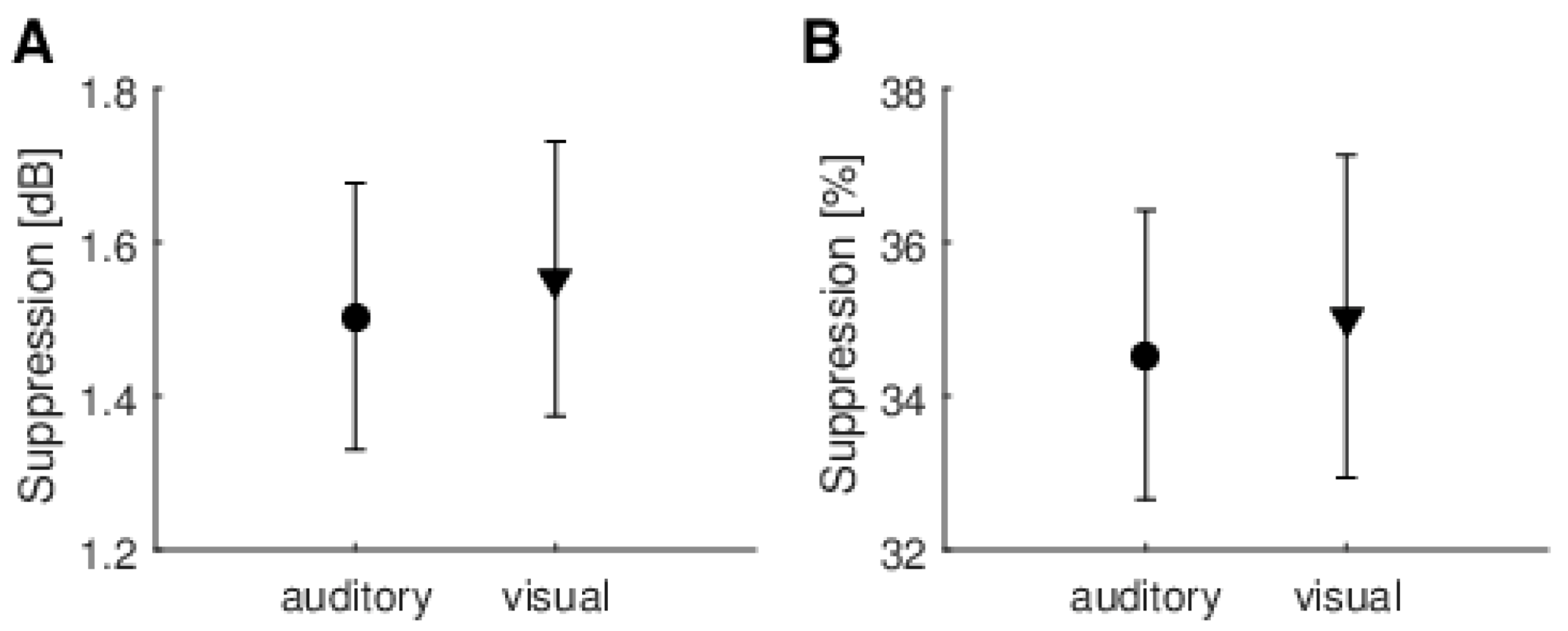
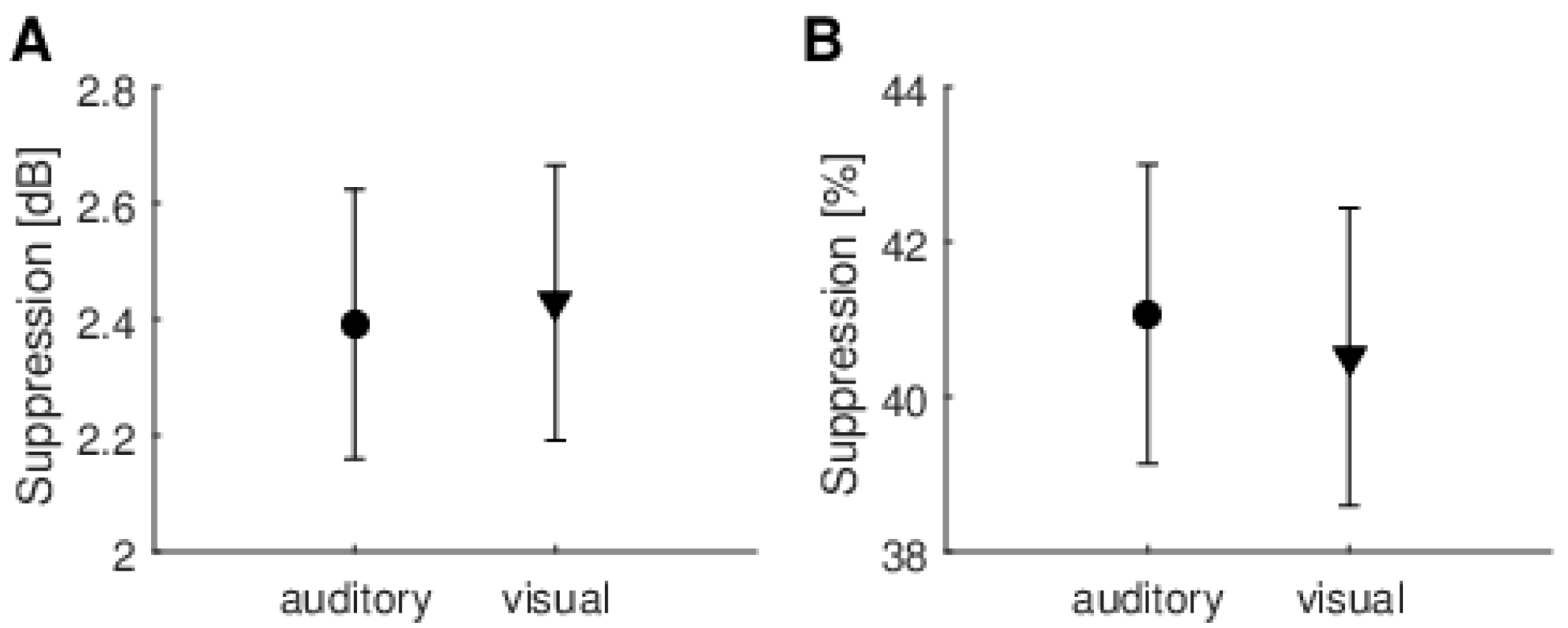
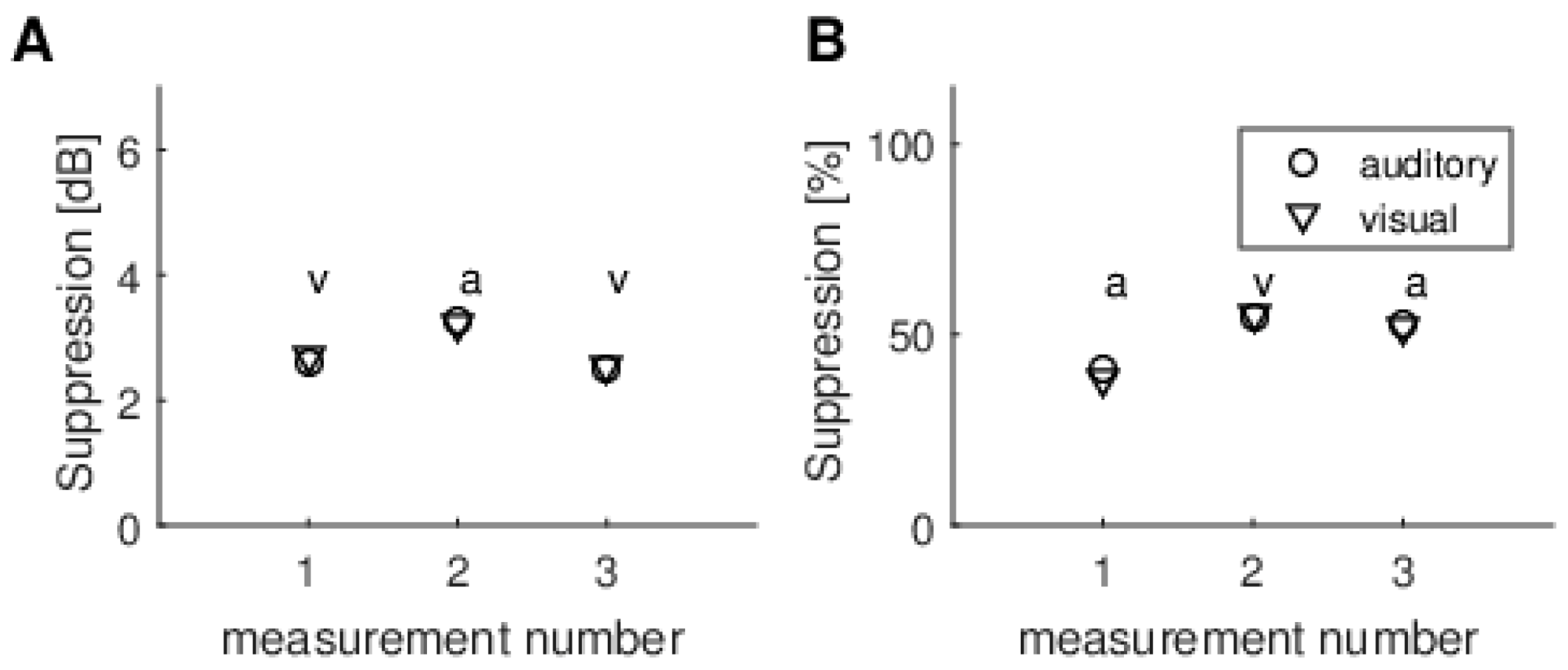
Another issue is measurement reliability. Looking at
Figure 9, which shows multiple recordings from the same subject, it is clear that differences in TEOAE suppression values among modalities are smaller than those between consecutive sessions. It should be underlined that the variability of TEOAE suppression between sessions observed here, i.e., about 1 dB, is not related to this particular measurement setup. Using different equipment, other authors have observed even larger variability (e.g., [
35]).
When different methods of calculating the MOC reflex were used (raw dB vs. normalized %), it was often the case that one method produced higher figures than the other. This was especially clear for data from several repeats of the experiment for a single person (
Figure 9). This is further evidence that the fluctuations are random and not due to some systematic effect.
Unlike some previous studies which used custom-built systems (e.g., [
15,
16,
17]), we used a readily available commercial system that could easily be replicated, especially if the ERP part was omitted. Our results give reassurance to clinicians that the subject’s level of attention is not likely to affect measurements.
Another interesting source of variability was that shown by the time–frequency analysis using the MP method. Resonance modes of greatest amplitude came from different frequency bands (
Figure 7B). These resonance modes were not suppressed in any systematic way, suggesting that broadband suppression values are probably better indicators of overall MOC function. Some previous studies have also shown that broadband suppression provided greater reliability than when a particular frequency band was used [
36,
37,
38].
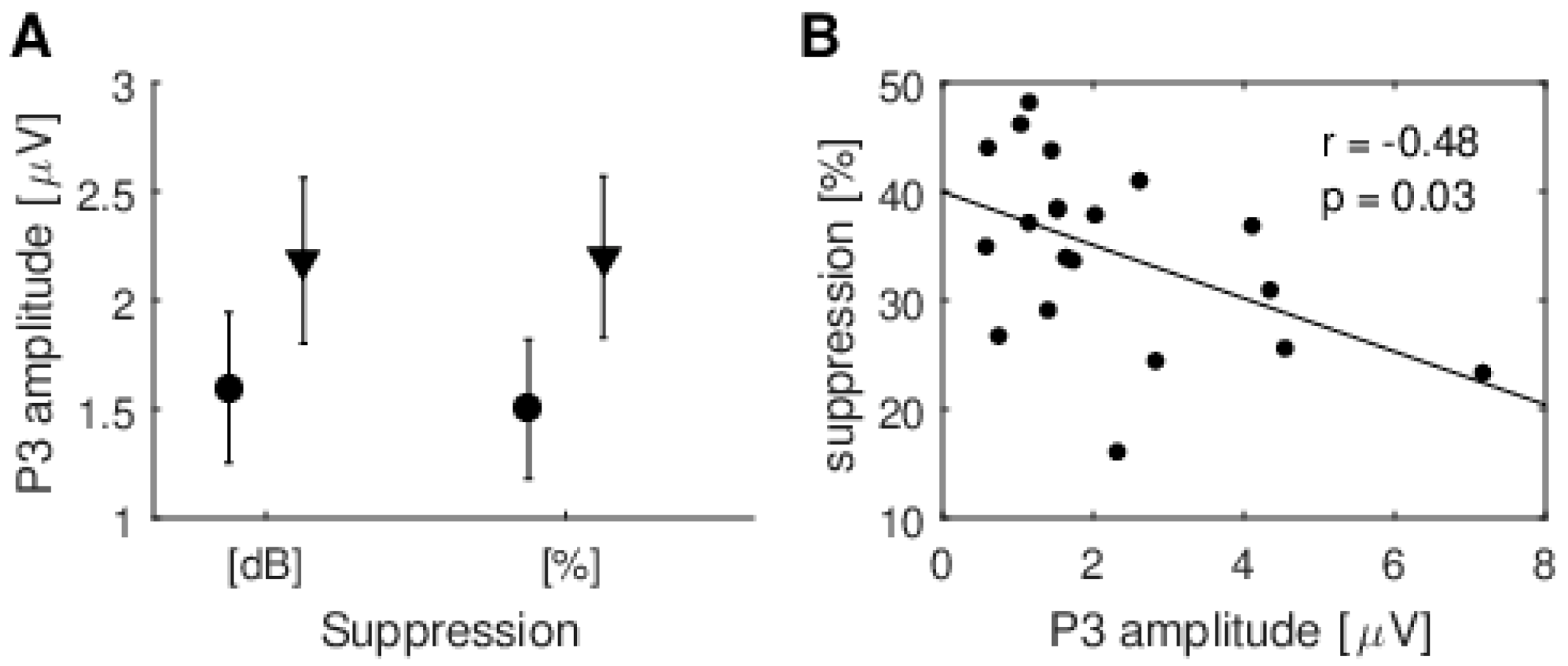
Despite the fact that the study showed no effect of change of modality on TEOAE suppression, it did provide another interesting result, namely that the amplitude of the P3 cognitive potential increased at the same time as normalized % TEOAE suppression decreased. It appears as if cortical processes may be able to compensate for some lack of synchronization at the peripheral level. This might explain the study by Marrufo-Perez et al. [
39] who showed that adaptation to noise by cochlear implant users does not differ statistically from that of listeners with normal hearing. It is known that ERPs, as well as TEOAE suppression by CAS, show huge intersubject variability. In both cases, there are subjects with prominent responses and others in which it is difficult to record any, despite being normal in every other respect. In cases of ERPs, as with TEOAEs, such variability remains unexplained. Perhaps the variability is normal, and one system just compensates for the other. Such a compensation scheme might be generally small and not affect overall perception, but it might be crucial in cases of impairments. For example, the following hypothesis can be made: That cochlear implant users do not adapt to noise in any way different from normal hearing individuals. We suppose that although the MOC does not seem to be involved, it is really the cortex compensating for the MOC.
It is difficult to compare our results with previous studies, as there are few papers combining OAEs and ERPs. Furthermore, in these papers, the procedures vary significantly. For example, the recent study by Rao et al. [
40] showed no connection between P3 and OAE suppression when listening in noise. The present results might be more closely related to a study by Dragicevic et al. [
17] who showed some correlations in the modulation of OAEs and ERPs, although these were of a different kind than in the present study. There are also the interesting results by Riecke et al. [
41] who showed some relationship between OAEs and the N1, although our work failed to detect such a connection.
There are some indications that attentional states may differentially affect MOC reflexes in the right and left ears [
42]. However, in the current study, we decided not to complicate the issue and chose to record TEOAEs only from the right ear. Our reasoning was that TEOAE response levels are known to be higher in the right ear, and therefore SNRs would also be higher [
43], and high SNRs are crucial for acquiring reliable MOC reflexes [
19,
20,
21,
38].
4.1. Limitations
One factor that can always have an effect on MOC measurements is middle ear muscle reflexes (MEMRs). Some researchers who use custom-built equipment have developed procedures that allow MEMR contamination of OAEs to be detected (e.g., [
16,
21]). Here, we used a system in which it was not possible to use such a procedure. Nevertheless, all ears had ART above 80 dB, which is much above the stimulation level of 65 dB used in TEOAE recordings (and the 60 dB for CAS). Together with earlier results which show that only a small percentage of ears are contaminated by MEMRs at such stimulus levels (e.g., [
44,
45]), we assume that MEMRs had little to no influence on the results reported here. In line with this idea, Francis et al. [
16] used a specific test for MEMRs and reported that they found only a small fraction of single responses from each subject which were due to possible MEMR contamination. At the same time, they failed to find any systematic pattern in these responses (which they removed just to be safe) and concluded that they were unsure if they were indeed contaminated by MEMRs. This finding shows how difficult the MEMR problem is. Certainly, if MOC measures are to be used clinically it is necessary that commercial OAE systems provide ways of detecting MEMRs.
In our work, we decided not to do no-task recordings, as subjects became exhausted doing them. Furthermore, in our previous study [
18], we did not find significant differences between OAEs recorded while performing a visual task and OAEs during the no-task condition. We also did not measure ERPs synchronized with visual stimuli. The reason was that we planned the experimental paradigm to demonstrate only that P3 could be evoked during an auditory task and would disappear when subjects were distracted by performing a visual task.
4.2. Implications
There seem to be no difference between the effect of visual and auditory attention on MOC as measured by TEOAEs. It seems that when attention is switched from one modality to another it is done by the cortex alone without the help of the periphery, in this case the ear. The study group was small (20 persons), although comparable with previous studies that showed a difference between auditory and visual tasks (e.g., [
9,
14,
15]).
The ERP recordings were very important here. If they were not recorded, one could argue that there may have been no difference in the periphery (TEOAEs), since there would be no evidence the cortex was in a different state. The ERPs provide proof that there are indeed two different cortical effects of the performed tasks, while it seems there was no effect on TEOAEs. Furthermore, the negative correlation between suppression of TEOAEs and the P3 amplitude during the auditory task seems to suggest that the cortex compensates for lower synchronization at the MOC level.
A secondary aspect of this study, flowing from the primary finding that a change in attentional state does not seem to affect the MOC reflex, is that it is permissible to use distractors during suppression experiments. For example, it might be beneficial to show a movie while making MOC reflex measurements. This may help some patients, especially children, relax and sit still (e.g., [
37,
38]). At the same time, there seems to be no need to control for attention in OAE measurements (as suggested by other recent papers, e.g., [
21]). Furthermore, if OAE suppression is not affected by conscious attention, one might conclude that sleep should not hinder the MOC inhibitory circuit. However, from our experience, when a person sleeps during an OAE recording, the noise levels increase, causing a decrease in SNR. The higher noise levels are mostly related to louder breathing and more uncontrolled movement. As already mentioned, high SNR is crucial for reliable MOC reflex testing, and therefore performing such measurements during sleep could be difficult.
A more general implication of this study is that making a connection between various aspects of sound processing and the MOC reflex needs to be done with caution. Even though we ensured good quality measurements with high SNRs, we did not find a difference between tasks (unlike in some previous experiments). This is in line with some other studies that failed to confirm earlier reported effects on the MOC reflex, for example, there are studies that have failed to show any connection between the MOC reflex and gender or laterality [
46], adaptation to noise and central auditory processing [
39,
40], auditory processing disorders [
47], tinnitus [
48], or sickle cell disease [
49]. Specifically, it has been suggested that previous work on auditory processing disorders probably did not fulfill appropriate SNR criteria in order to ensure reliability [
47]. Nevertheless, despite the abovementioned findings, we do not completely discard the possibility that there is cooperation between ear and cortex when switching between modalities. However, such an effect on OAEs might be very small and below the level of detection with current group sizes, as Beim et al. also suggested [
50].

 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}










