Features of Action Potentials from Identified Thalamic Nuclei in Anesthetized Patients



Abstract
:1. Introduction
2. Methods
2.1. Patients
2.2. Surgical Procedures
2.3. Reconstruction of the Trajectory
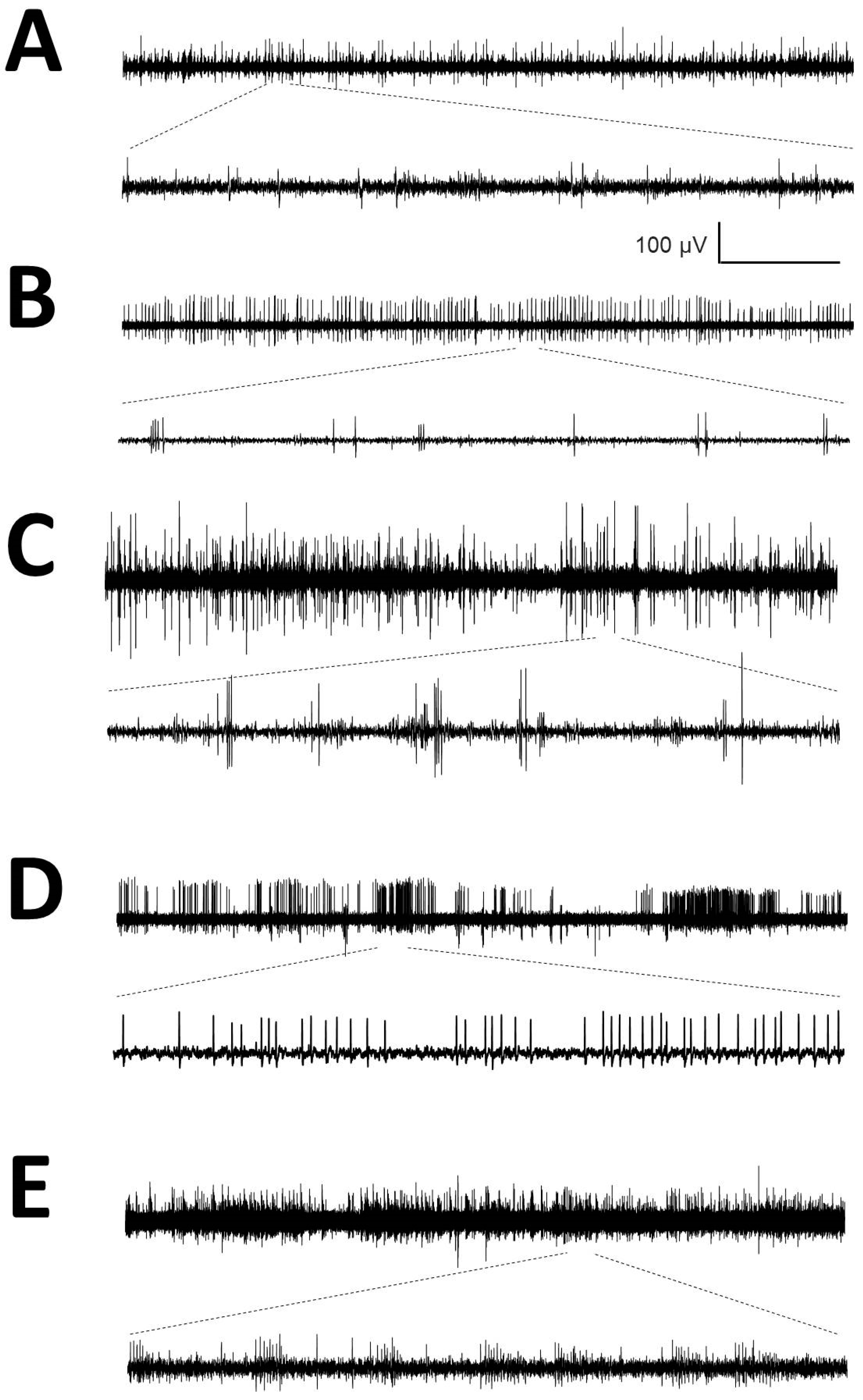
2.4. Sorting Spikes and Analysis of Action Potentials
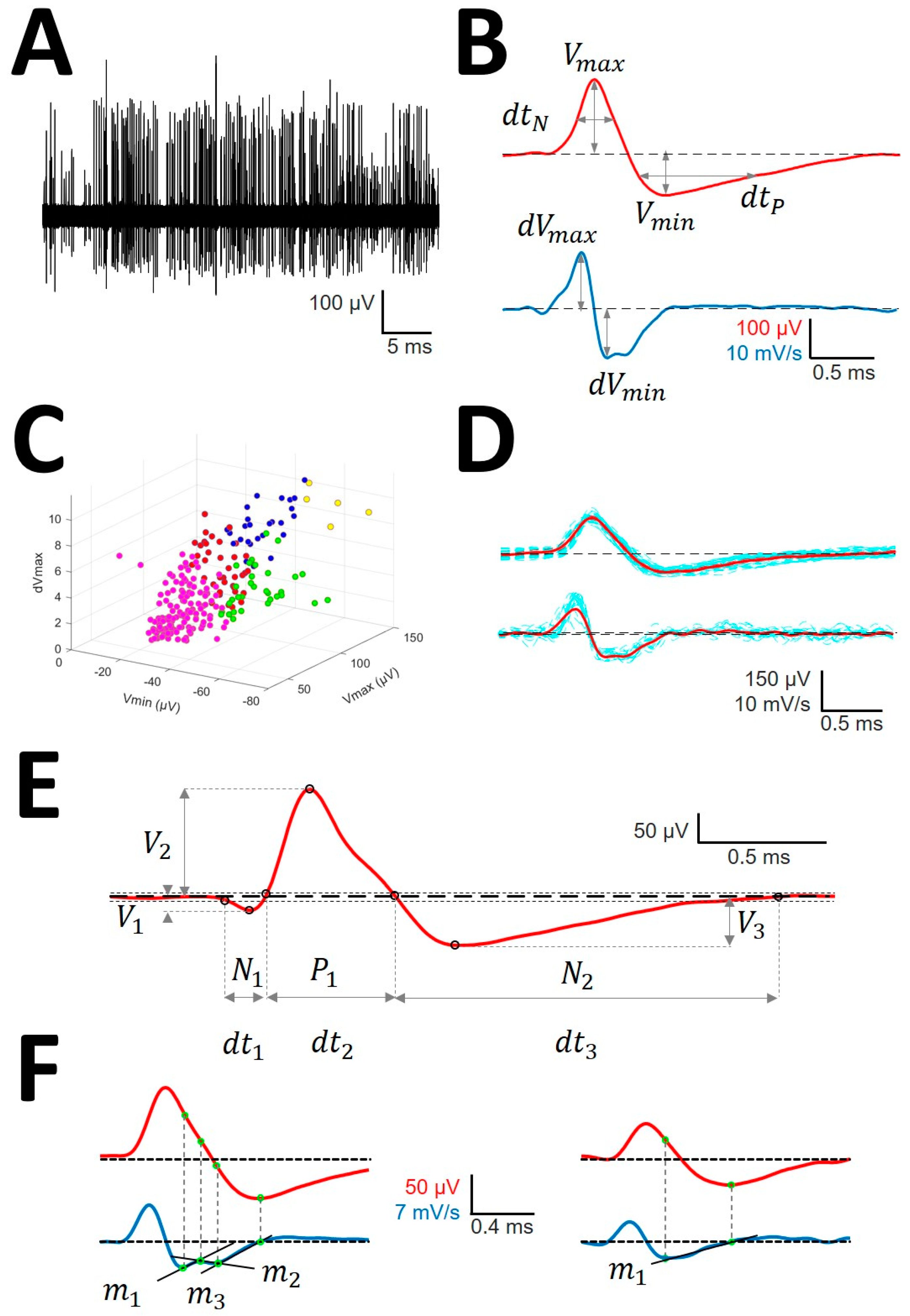
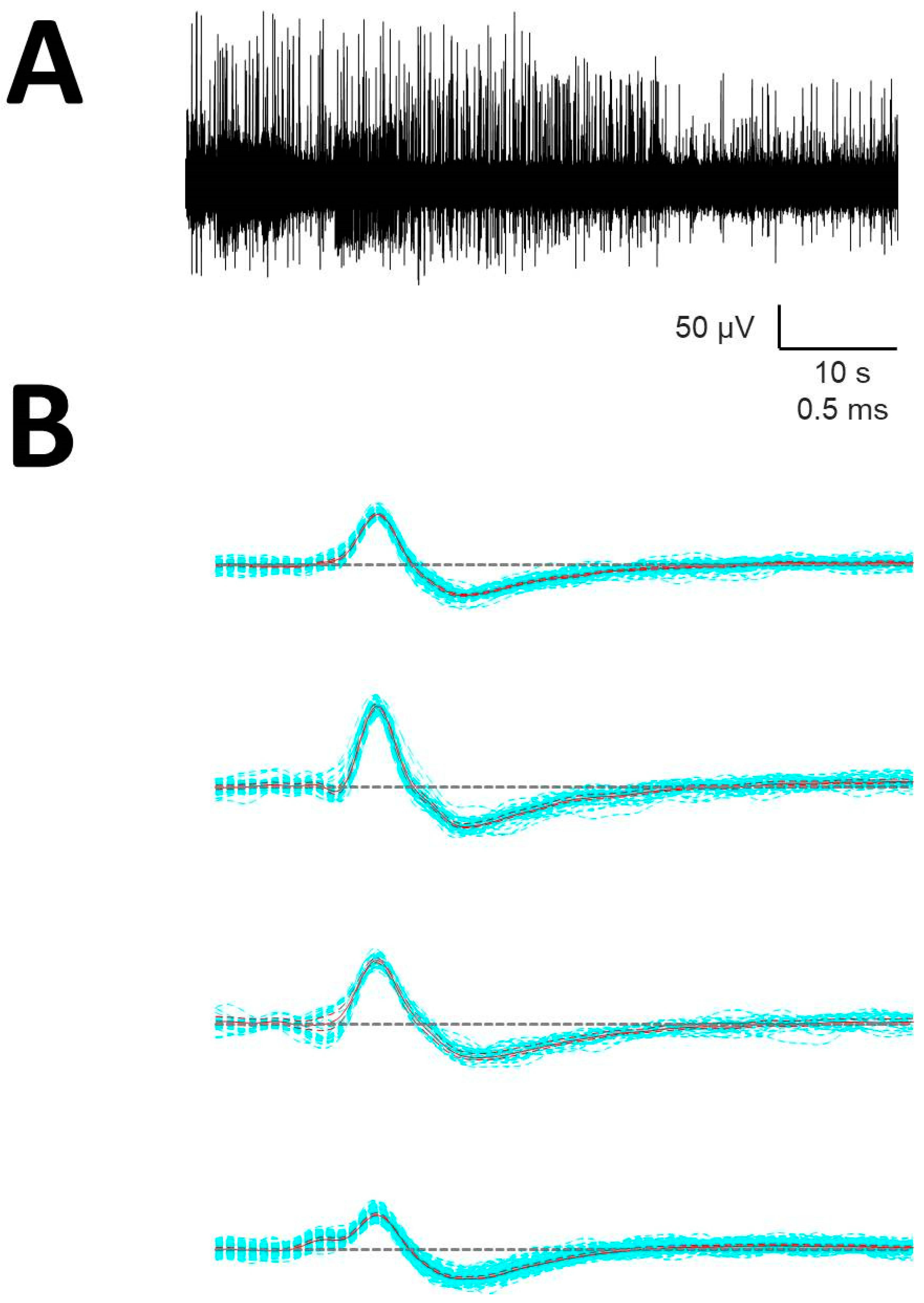
- Identification of APs. For every trace (Figure 1A), we computed a maximum (V+) and minimum (V−) voltage threshold (in µV), defined as , where is the mean and is the standard deviation. APs must have two phases (depolarization and repolarization); therefore, we identified a tentative AP when a positive/negative (P/N) phase was followed by a negative/positive (N/P) phase in a period of 0.3–0.6 ms. APs were defined as positive or negative according to the highest component identified.
- Clustering was performed by an agglomerative hierarchical method, with distance between groups computed by farthest procedure. APs sharing similar morphologies were ascribed to the same neuron. For every AP, we measured the maximum (Vmax) and minimum voltages (Vmin, in µV), durations of negative (dtN) and positive phases at half-amplitude (dtP in ms), and maximum (dVmax) and minimum values of the first derivative (dVmin, in mV/s). These measures can be considered as a 6-dimension vector for every k-AP, (Figure 1B). Then, we clustered the APs with similar properties using the standardized Euclidean distances (see below) (dE) (Figure 1C) [25].
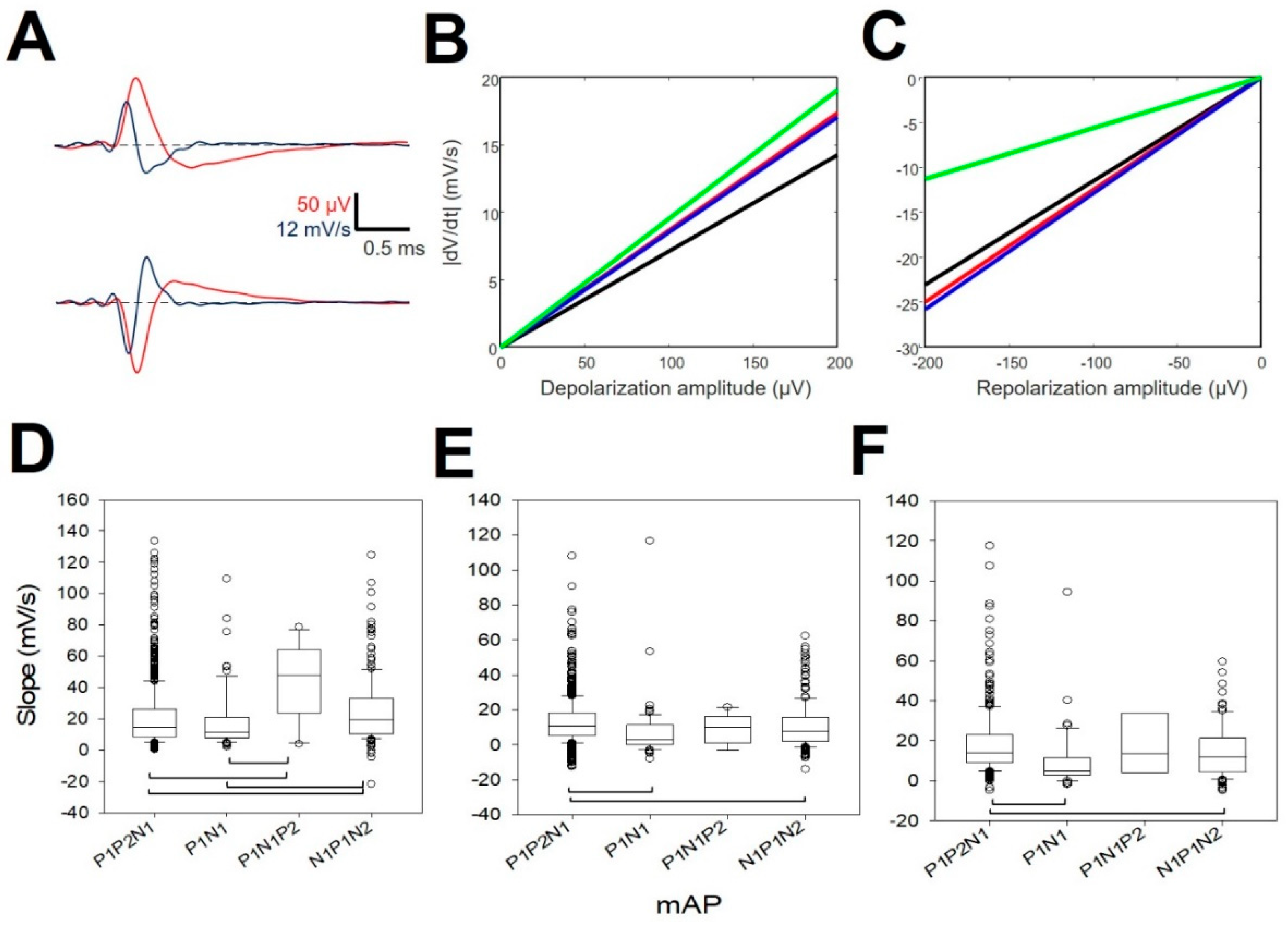
- Construction of the mean action potential (mAP). All of the APs from the same cluster were averaged to obtain a canonical waveform (Figure 1D, upper row), as were the derivatives to obtain the mean derivative (mDAP, 1D, lower row). A minimum of 10 APs were averaged. The first 300 µs (72 points) of baseline were used to compute the maximum (VAP+) and minimum (VAP−) voltage thresholds (in µV), defined as , where is the mean and the standard deviation. We used these thresholds to identify hallmark points in mAPs (Figure 1E). Every phase can be characterized by its polarity (P/N), duration (dti), and amplitude (Vi, i = 1, 2, 3).
- Rectification of the repolarizing phase. We analyzed the number of phases and their slopes. The local maxima and minima of the mDAP between the lowest value and the zero crossing were taken to define uniform dynamics in the mAP (see Figure 1F). We used two consecutive points, i,j, in mDAP () to find the slope (m) following the formula:
2.5. Evaluation of Global Similarity
2.6. Classification of mAP According to Morphology
2.7. Statistics
3. Results
3.1. Reconstruction of Trajectories
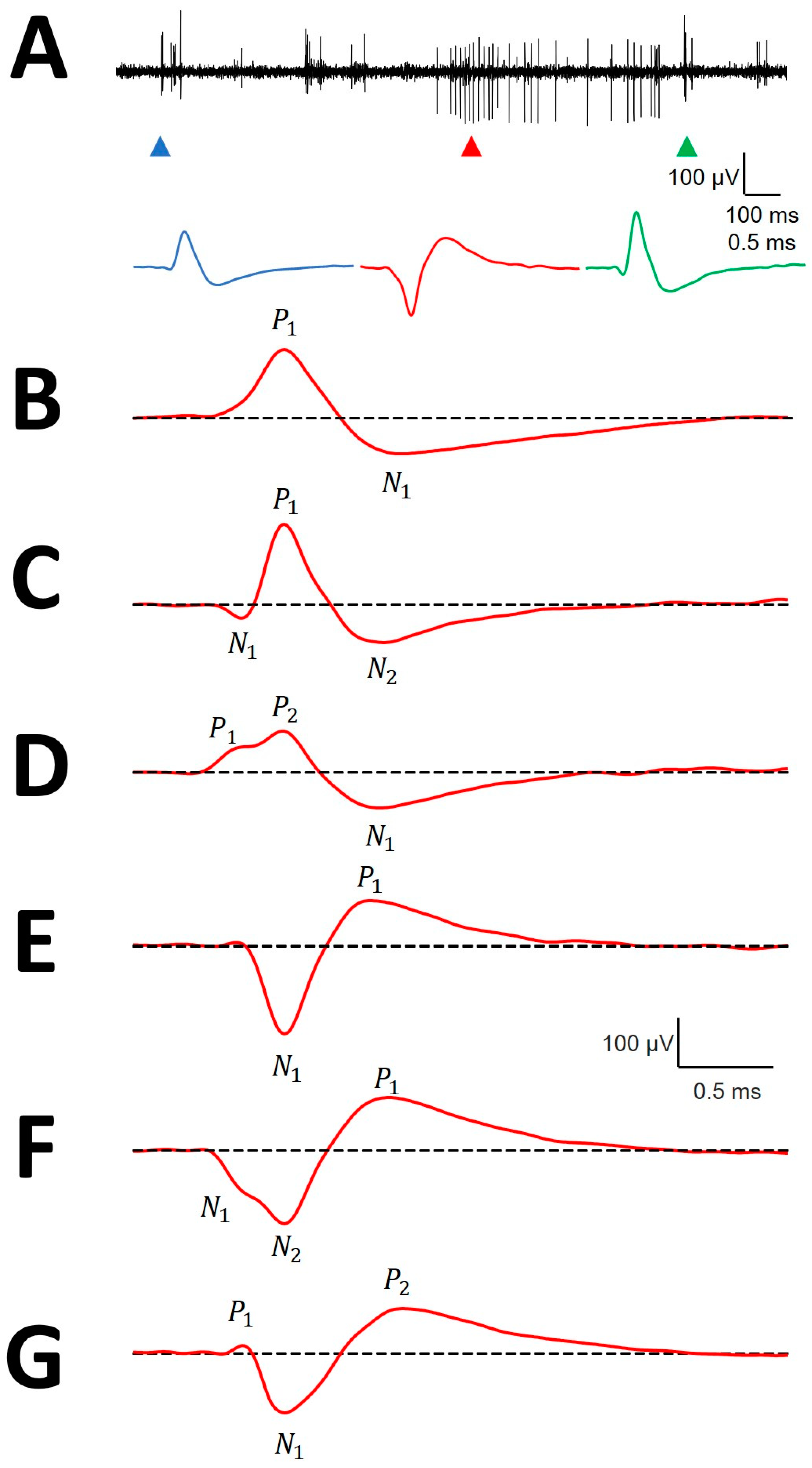
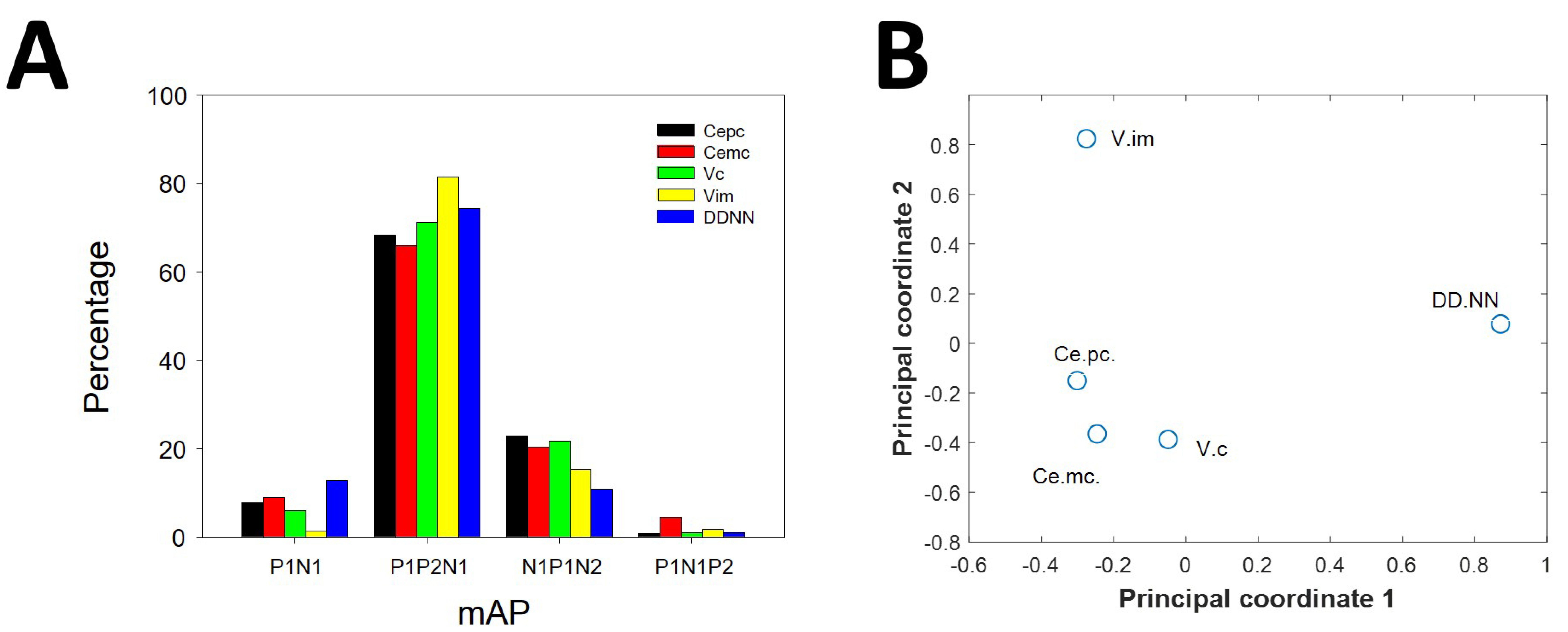
3.2. Types of mAP According to Structure
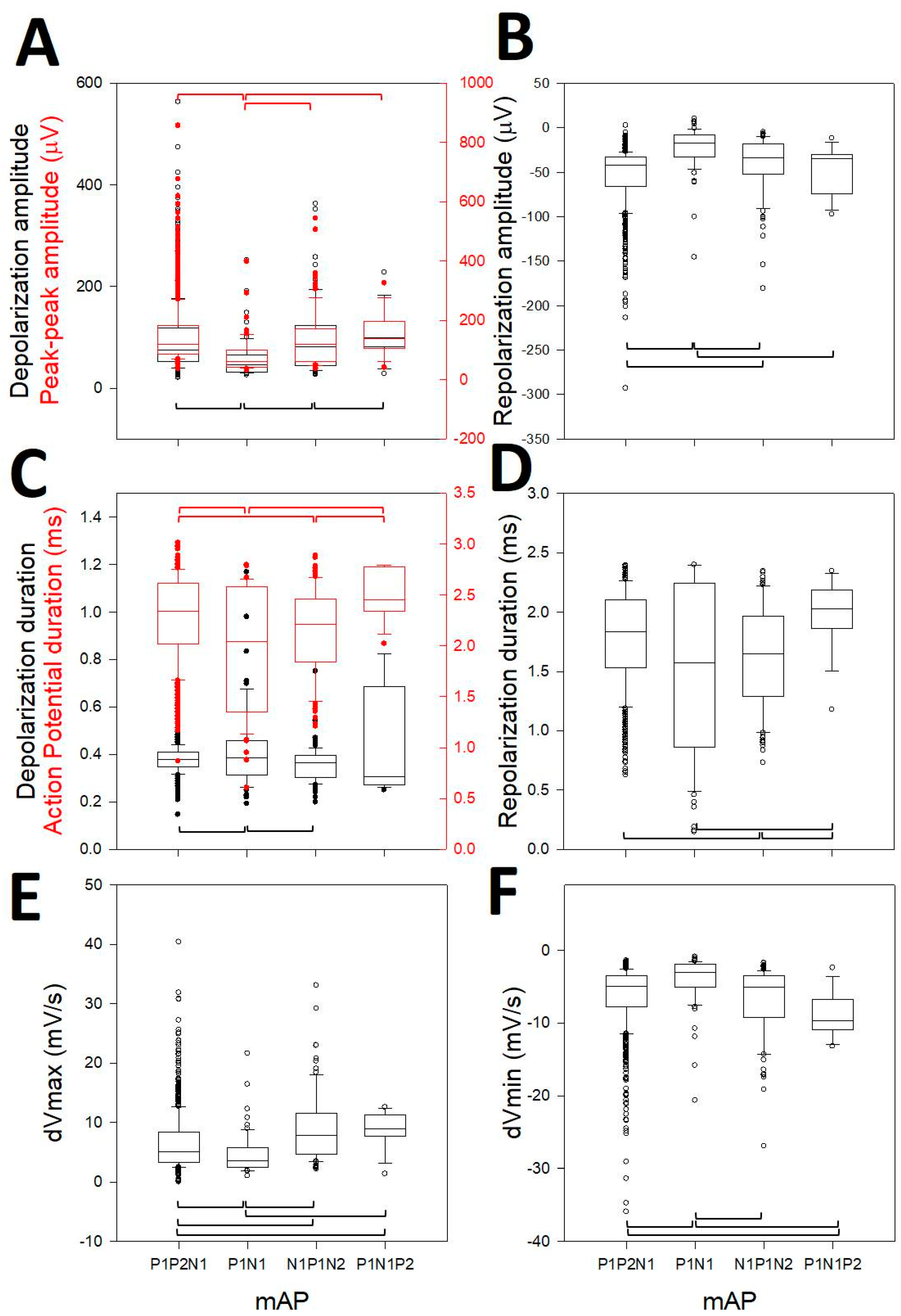
3.3. Canonical Description of mAP
3.4. Properties of the First Derivative
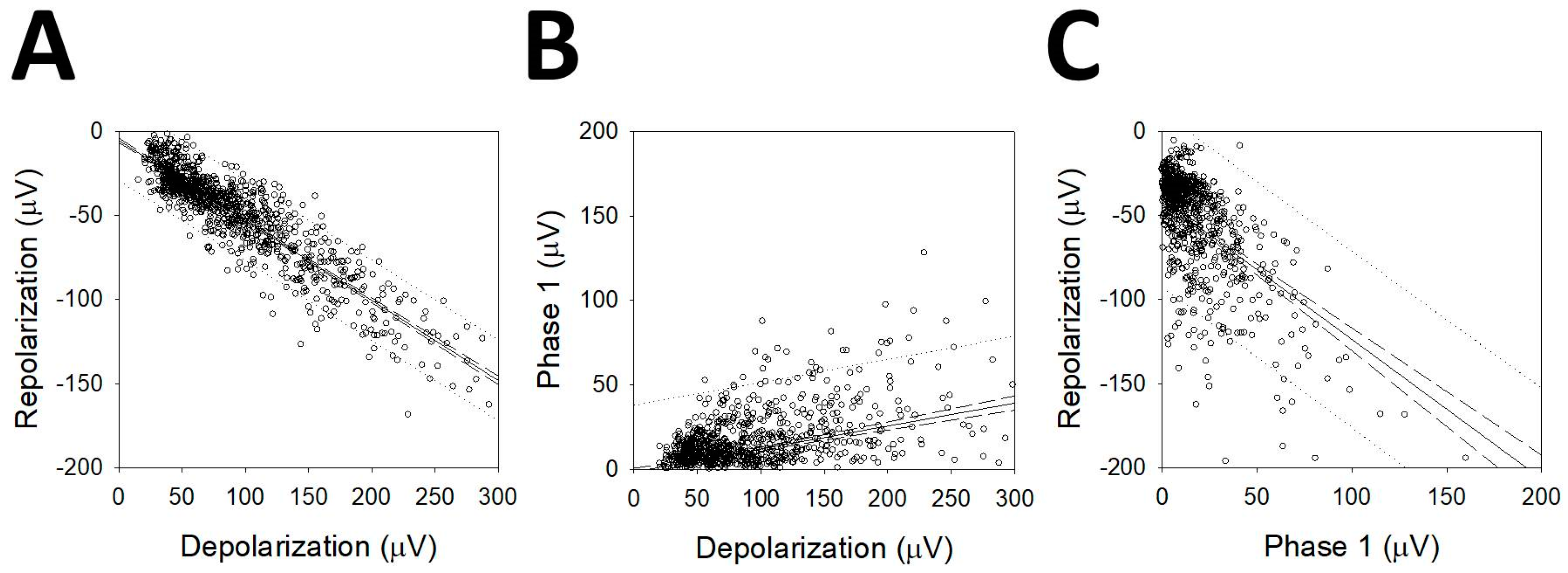
3.5. Analysis of the First Phase
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Ce.pc | Ce.mc | V.c | V.im | DD.NN |
|---|---|---|---|---|---|
| N | 11 | 10 | 32 | 3 | 14 |
| N1 (µV) | 47.6 ± 7.7 | 40.0 ± 4.1 | 51.0 ± 4.3 | 48.9 ± 12.3 | 86.2 ± 18.0 |
| P1 (µV) | −15.5 ± 5.7 | −18.4 ± 3.2 | −16.1 ± 3.1 | −17.2 ± 9.7 | −43.7 ± 10.2 |
| durN1 (ms) | 0.38 ± 0.03 | 0.41 ± 0.05 | 0.49 ± 0.04 | 0.34 ± 0.04 | 0.37 ± 0.03 |
| durP1 (ms) | 1.59 ± 0.04 | 1.41 ± 0.23 | 1.49 ± 0.13 | 1.53 ± 0.43 | 1.64 ± 0.19 |
| Peak-peak (µV) | 63.7 ± 12.3 | 58.4 ± 6.8 | 69.0 ± 6.6 | 67.5 ± 20.1 | 129.8 ± 27.8 |
| durPA (ms) | 1.97 ± 0.24 | 1.82 ± 0.19 | 2.00 ± 0.11 | 1.87 ± 0.40 | 2.13 ± 0.17 |
| dVmax (mV/s) | 4.3 ± 0.8 | 3.5 ± 0.4 | 3.9 ± 0.4 | 4.1 ± 1.2 | 7.2 ± 1.6 |
| dVmin (mV/s) | −3.5 ± 0.74 | −3.1 ± 0.4 | −3.4 ± 0.4 | −2.8 ± 0.9 | −6.5 ± 15 |
| Properties | Ce.pc | Ce.mc | V.c | V.im | DD.NN |
|---|---|---|---|---|---|
| N | 95 | 74 | 376 | 180 | 81 |
| N1 (µV) | 19.5 ± 2.1 | 13.9 ± 1.6 | 15.2 ± 0.7 | 24.7 ± 1.9 | 19.4 ± 1.4 |
| N2 (µV) | 89.6 ± 6.4 | 95.5 ± 7.5 | 89.8 ± 3.0 | 111.4 ± 5.8 | 91.3 ± 5.3 |
| P1 (µV) | −50.1 ± 3.3 | −49.6 ± 1.5 | −51.6 ± 1.6 | −62.9 ± 2.9 | −54.8 ± 2.9 |
| durN1 (ms) | 0.11 ± 0.01 | 0.11 ± 0.01 | 0.12 ± 0.00 | 0.12 ± 0.01 | 0.12 ± 0.01 |
| durN2 (ms) | 0.37 ± 0.01 | 0.38 ± 0.01 | 0.38 ± 0.00 | 0.40 ± 0.00 | 0.39 ± 0.01 |
| durP1 (ms) | 1.68 ± 0.05 | 1.80 ± 0.02 | 1.80 ± 0.02 | 1.85 ± 0.02 | 1.71 ± 0.05 |
| Peak-peak (µV) | 139.4 ± 9.6 | 139.3 ± 4.4 | 144.5 ± 4.6 | 174.2 ± 8.6 | 146.1 ± 8.0 |
| durPA (ms) | 2.15 ± 0.05 | 2.28 ± 0.02 | 2.29 ± 0.02 | 2.37 ± 0.03 | 2.23 ± 0.05 |
| dVmax (mV/s) | 6.0 ± 0.5 | 6.5 ± 0.3 | 6.6 ± 0.03 | 7.4 ± 0.4 | 6.2 ± 0.4 |
| dVmin (mV/s) | −6.6 ± 0.5 | −6.0 ± 0.2 | −6.2 ± 0.2 | −6.9 ± 0.4 | −6.0 ± 0.4 |
| Properties | Ce.pc | Ce.mc | V.c | V.im | DD.NN |
|---|---|---|---|---|---|
| N | 32 | 23 | 115 | 34 | 12 |
| P1 (µV) | −12.5 ± 2.0 | −15.1 ± 2.3 | −15.8 ± 0.9 | −17.8 ± 2.2 | −17.1 ± 2.4 |
| N1 (µV) | 60.3 ± 6.1 | 80.5 ± 14.8 | 79.1 ± 4.6 | 119.2 ± 12.9 | 122.8 ± 19.8 |
| P2 (µV) | −23.8 ± 2.7 | −34.9 ± 7.0 | −34.5 ± 2.1 | −51.5 ± 5.6 | −56.9 ± 10.8 |
| durP1 (ms) | 0.13 ± 0.01 | 0.17 ± 0.02 | 0.16 ± 0.01 | 0.15 ± 0.01 | 0.17 ± 0.03 |
| durN1 (ms) | 0.34 ± 0.01 | 0.32 ± 0.01 | 0.37 ± 0.01 | 0.37 ± 0.01 | 0.40 ± 0.04 |
| durP2 (ms) | 1.51 ± 0.08 | 1.70 ± 0.09 | 1.69 ± 0.04 | 1.72 ± 0.07 | 1.54 ± 0.11 |
| Peak-peak (µV) | 84.1 ± 8.5 | 115.4 ± 21.4 | 113.6 ± 6.6 | 170.6 ± 18.4 | 179.7 ± 30.3 |
| durPA (ms) | 1.98 ± 0.08 | 2.03 ± 0.09 | 2.16 ± 0.04 | 2.25 ± 0.06 | 2.11 ± 0.12 |
| dVmax (mV/s) | 5.4 ± 0.6 | 8.2 ± 1.3 | 7.7 ± 0.4 | 11.4 ± 1.2 | 11.5 ± 1.7 |
| dVmin (mV/s) | −4.8 ± 0.5 | −6.3 ± 0.8 | −5.6 ± 0.3 | −7.9 ± 0.9 | −9.2 ± 1.6 |
| Properties | Ce.pc | Ce.mc | V.c | V.im | DD.NN |
|---|---|---|---|---|---|
| N | 1 | 5 | 5 | 4 | 2 |
| N1 (µV) | - | 8.8 ± 0.6 | 7.2 ± 2.2 | 5.3 ± 3.2 | - |
| P1 (µV) | - | −106.1 ± 6.8 | −88.8 ± 22.6 | −123.8 ± 36.4 | - |
| N2 (µV) | - | 32.1 ± 3.2 | 50.2 ± 15.8 | 62.3 ± 13.8 | - |
| durN1 (ms) | - | 0.10 ± 0.00 | 0.14 ± 0.03 | 0.01 ± 0.04 | - |
| durP1 (ms) | - | 0.27 ± 0.00 | 0.57 ± 0.12 | 0.48 ± 0.10 | - |
| durN2 (ms) | - | 2.11 ± 0.09 | 1.85 ± 0.19 | 1.98 ± 0.07 | - |
| Peak–peak (µV) | - | 138.2 ± 9.9 | 138.9 ± 37.8 | 186.1 ± 50.0 | - |
| durPA (ms) | - | 2.39 ± 0.09 | 2.56 ± 0.15 | 2.47 ± 0.12 | - |
| dVmax (mV/s) | - | 10.3 ± 0.7 | 8.4 ± 2.0 | 7.6 ± 0.9 | - |
| dVmin (mV/s) | - | −11.0 ± 0.7 | −7.4 ± 1.6 | −8.7 ± 1.6 | - |
| Phases During Repolarization (mV/s) | ||||
|---|---|---|---|---|
| Properties | N | m1 | m2 | m3 |
| P1P2N1 | 768 | 21.10 ± 0.72 | 13.08 ± 0.48 | 18.08 ± 0.90 |
| P1N2 | 69 | 18.54 ± 2.36 | 7.26 ± 1.99 | 10.08 ± 2.55 |
| N1P1N2 | 212 | 24.78 ± 1.44 | 11.03 ± 0.91 | 14.33 ± 1.31 |
| P1N1P2 | 15 | −43.84 ± 6.21 | −9.66 ± 2.14 | −17.75 ± 7.70 |
References
- Nieuwenhuys, R.; Voogd, J.; Van Huijzen, C. Diencephalo: Dorsal Thalamus. In The Human Central Nervous System; Locked, P., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 253–279. [Google Scholar]
- Baxter, D.A.J.H.; Byrne, J.H. Ionic conductance mechanisms contributing to the electrophysiological properties of neurons. Curr. Opin. Neurobiol. 1991, 1, 105–112. [Google Scholar] [CrossRef]
- Chandler, S.H.; Hsaio, C.F.; Inoue, T.; Goldberg, L.J. Electrophysiological properties of guinea pig trigeminal motoneurons recorded in vitro. J. Neurophysiol. 1994, 71, 129–145. [Google Scholar] [CrossRef] [PubMed]
- Samios, V.N.; Inoue, T. Interleukin-1β and interleukin-6 affect electrophysiological properties of thalamic relay cells. Neurosci. Res. 2014, 87, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Traub, R.D.; Wong, R.K.; Miles, R.; Michelson, H. A model of a CA3 hippocampal pyramidal neuron incorporating voltage-clamp data on intrinsic conductances. J. Neurophysiol. 1991, 66, 635–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormick, D.A. Action Potential. In Fundamental Neuroscience, 2nd ed.; Squire, L.R., Bloom, F.E., McConnell, S.K., Roberts, J.L., Spitzer, N.C., Zigmon, M.J., Eds.; Academic Press: New York, NY, USA, 2003; pp. 139–162. [Google Scholar]
- Koester, J.; Siegelbaum, S.A. Membrane Potential. In Principles of Neural Science, 5th ed.; Kandel, E.R., Schwartz, J.H., Jessell, T., Siegelbaum, S.A., Hudspeth, A.J., Eds.; Elsevier: New York, NY, USA, 2013; pp. 148–171. [Google Scholar]
- Mandge, D.; Manchanda, R. A biophysically detailed computational model of urinary bladder small DRG neuron soma. PLoS Comput. Biol. 2018, 14, e1006293. [Google Scholar] [CrossRef] [PubMed]
- Heinricher, M.M. Principles of extracellular single-unit recording. In Microelectrode Recording in Movement Disorder Surgery, 1st ed.; Israel, Z., Burchiel, K.J., Eds.; Thieme: New York, NY, USA, 2004; pp. 8–13. [Google Scholar]
- Obwegeser, A.A.; Uitti, R.J.; Turk, M.F.; Strongosky, A.J.; Wharen, R.E. Thalamic stimulation for the treatment of midline tremors in essential tremor patients. Neurology 2000, 54, 2342–2344. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Wang, S.; Stein, J.F.; Aziz, T.Z.; Green, A. Reciprocal interactions between the human thalamus and periaqueductal gray may be important for pain perception. Exp. Brain Res. 2013, 232, 527–534. [Google Scholar] [CrossRef]
- Vega-Zelaya, L.; Torres, C.V.; Navas, M.; Pastor, J. Neurophysiological characterization of thalamic nuclei in anaesthetized humans. Brain Sci. 2019, 9, 312. [Google Scholar] [CrossRef] [Green Version]
- Shimamoto, S.A.; Larson, P.S.; Ostrem, J.L.; Glass, G.A.; Turner, R.S.; Starr, P.A. Physiological identification of the human pedunculopontine nucleus. J. Neurol. Neurosurg. Psychiatry 2010, 81, 80–86. [Google Scholar] [CrossRef]
- Alam, M.; Sanghera, M.K.; Schwabe, K.; Lütjens, G.; Jin, X.; Song, J.; Von Wrangel, C.; Stewart, R.M.; Jankovic, J.; Grossman, R.G.; et al. Globus pallidus internus neuronal activity: A comparative study of linear and non-linear features in patients with dystonia or Parkinson’s disease. J. Neural Transm. (Vienna) 2015, 123, 231–240. [Google Scholar] [CrossRef]
- Li, X.; Zhuang, P.; Hallett, M.; Zhang, Y.; Li, J.; Li, Y. Subthalamic oscillatory activity in parkinsonian patients with off-period dystonia. Acta Neurol. Scand. 2016, 134, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Micieli, R.; Lopez Rios, A.L.; Plata Aguilar, R.; Posada, L.F.B.; Hutchison, W.D. Single-unit analysis of the human posterior hypothalamus and red nucleus during deep brain stimulation for aggressivity. J. Neurosurg. 2017, 126, 1158–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gold, C.; Henze, D.A.; Koch, C.; Buzsáki, G. On the Origin of the Extracellular Action Potential Waveform: A Modeling Study. J. Neurophysiol. 2006, 95, 3113–3128. [Google Scholar] [CrossRef] [PubMed]
- Vega-Zelaya, L.; Torres, C.; Sola, R.G.; Pastor, J. Characterization of thalamic nuclei and somatosensory evoked potentials in anesthetized humans. Clin. Neurophysiol. 2016, 127, e203–e205. [Google Scholar] [CrossRef]
- Hassler, R. Anatomy of the thalamus. In Introduction to Stereotaxis with an Atlas of the Human Brain; Schaltenbrand, G., Bailey, P., Eds.; Thieme: Stuttgart, Germany, 1959; pp. 230–290. [Google Scholar]
- Pastor, J.; Hernando-Requejo, V.; Domínguez-Gadea, L.; De Llano, I.; Meilán-Paz, M.L.; Martínez-Chacón, J.L.; Sola, R.G. Impact of experience on improving the surgical outcome in temporal lobe epilepsy. Rev. Neurol. 2005, 41, 709–716. [Google Scholar]
- Sola, R.G.; Hernando-Requejo, V.; Pastor, J.; García-Navarrete, E.; DeFelipe, J.; Alijarde, M.T.; Sánchez, A.; Domínguez-Gadea, L.; Martín-Plasencia, P.; Maestú, F.; et al. Pharmacoresistant temporal-lobe epilepsy. Exploration with foramen ovale electrodes and surgical outcomes. Rev. Neurol. 2005, 41, 4–16. [Google Scholar]
- Pastor, J.; Vega-Zelaya, L. A new potential specifically marks the sensory thalamus in anaesthetized patients. Clin. Neurophysiol. 2019, 130, 1926–1936. [Google Scholar] [CrossRef]
- Van Drongelen, W. (Ed.) Filters: Digital Filters. In Signal Processing for Neuroscientists; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Rey, H.G.; Pedreira, C.; Quian Quiroga, R. Past, present and future of spike sorting techniques. Brain Res. Bull. 2015, 119, 106–117. [Google Scholar] [CrossRef] [Green Version]
- Peña, D. Análisis de Datos Multivariantes; McGraw-Hill/Interamericana de España: Madrid, Spain, 2002. [Google Scholar]
- Van Hemert, F.; Jebbink, M.; Van Der Ark, A.; Scholer, F.; Berkhout, B. Euclidean Distance Analysis Enables Nucleotide Skew Analysis in Viral Genomes. Comput. Math. Methods Med. 2018, 2018, 6490647. [Google Scholar] [CrossRef] [Green Version]
- Peña, D.; Prieto, F.J. The kurtosis coefficient and the linear discriminant function. Stat. Probab. Lett. 2000, 49, 257–261. [Google Scholar] [CrossRef] [Green Version]
- Spiegel, M.R.; Schiller, J.; Srinivasan, R.A. Probabilidad y Estadística; McGraw-Hill: Bogotá, Colombia, 2003. [Google Scholar]
- Henze, D.A.; Borhegyi, Z.; Csicsvari, J.; Mamiya, A.; Harris, K.D.; Buzsáki, L.R.B.H.R.E.E.S.G. Intracellular Features Predicted by Extracellular Recordings in the Hippocampus In Vivo. J. Neurophysiol. 2000, 84, 390–400. [Google Scholar] [CrossRef] [PubMed]
- Vega-Zelaya, L.; Sola, R.G.; Pulido, P.; Pastor, J. Do we Need to Wake Patients up during Cortical Surgery? J. Cancer Res. Updat. 2018, 7, 84–96. [Google Scholar] [CrossRef]
- Pastor, J.; Vega-Zelaya, L. Can We Put Aside Microelectrode Recordings in Deep Brain Stimulation Surgery? Brain Sci. 2020, 10, 571. [Google Scholar] [CrossRef] [PubMed]
- Laboy-Juárez, K.J.; Ahn, S.; Feldman, D.E. Normalized template matching method for improving spike detection in extracellular voltage recordings. Sci. Rep. 2019, 9, 12087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, J.E.; Magland, J.F.; Barnett, A.H.; Tolosa, V.M.; Tooker, A.C.; Lee, K.Y.; Shah, K.G.; Felix, S.H.; Frank, L.M.; Greengard, L.F. A Fully Automated Approach to Spike Sorting. Neuron 2017, 95, 1381–1394.e6. [Google Scholar] [CrossRef] [Green Version]
- Rossant, C.; Kadir, S.N.; Goodman, D.F.M.; Schulman, J.; Hunter, M.L.D.; Saleem, A.B.; Grosmark, A.; Belluscio, M.; Denfield, G.H.; Ecker, A.S.; et al. Spike sorting for large, dense electrode arrays. Nat. Neurosci. 2016, 19, 634–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franke, F.; Quian Quiroga, R.; Hierlemann, A.; Obermayer, K. Bayes optimal template matching for spike sorting—Combining fisher discriminant analysis with optimal filtering. J. Comput. Neurosci. 2015, 38, 439–459. [Google Scholar] [CrossRef]
- Su, C.-K.; Chiang, C.-H.; Lee, C.-M.; Fan, Y.-P.; Ho, C.-M.; Shyu, L.-Y. Computational solution of spike overlapping using data-based subtraction algorithms to resolve synchronous sympathetic nerve discharge. Front. Comput. Neurosci. 2013, 7, 149. [Google Scholar] [CrossRef] [Green Version]
- Ghahari, A.; Kumar, S.R.; Badea, T.C. Identification of Retinal Ganglion Cell Firing Patterns Using Clustering Analysis Supplied with Failure Diagnosis. Int. J. Neural Syst. 2018, 28, 1850008. [Google Scholar] [CrossRef]
- Mahallati, S.; Bezdek, J.C.; Popovic, M.R.; Valiante, T.A. Cluster tendency assessment in neuronal spike data. PLoS ONE 2019, 14, e0224547. [Google Scholar] [CrossRef] [Green Version]
- Weinberger, M.; Hamani, C.; Hutchison, W.D.; Moro, E.; Lozano, A.M.; Dostrovsky, J.O. Pedunculopontine nucleus microelectrode recordings in movement disorder patients. Exp. Brain Res. 2008, 188, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Galazky, I.; Kaufmann, J.; Voges, J.; Hinrichs, H.; Heinze, H.-J.; Sweeney-Reed, C.M. Neuronal spiking in the pedunculopontine nucleus in progressive supranuclear palsy and in idiopathic Parkinson’s disease. J. Neurol. 2019, 266, 2244–2251. [Google Scholar] [CrossRef] [PubMed]
- De La Prida, L.M.; Benavides-Piccione, R.; Sola, R.; Pozo, M.A. Electrophysiological properties of interneurons from intraoperative spiking areas of epileptic human temporal neocortex. NeuroReport 2002, 13, 1421–1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anastassiou, C.A.; Perin, R.; Buzsáki, G.; Markram, H.; Koch, C. Cell type- and activity-dependent extracellular correlates of intracellular spiking. J. Neurophysiol. 2015, 114, 608–623. [Google Scholar] [CrossRef] [PubMed]
- Holt, G.R.; Koch, C. Electrical Interactions via the Extracellular Potential near Cell Bodies. J. Comput. Neurosci. 1999, 6, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Bakkum, D.J.; Obien, M.E.J.; Radivojevic, M.; Jäckel, D.; Frey, U.; Takahashi, H.; Hierlemannb, A. The Axon Initial Segment is the Dominant Contributor to the Neuron’s Extracellular Electrical Potential Landscape. Adv. Biosyst. 2019, 3, e1800308. [Google Scholar] [CrossRef] [PubMed]
- Gold, C.; Girardin, C.C.; Martin, K.A.C.; Koch, K. High-amplitude positive spikes recorded extracellularly in cat visual cortex. J. Neurophysiol. 2009, 102, 3340–3351. [Google Scholar] [CrossRef] [Green Version]






| Patient | Gender | Age (Years) | History (Years) | Etiology | v-EEG | MRi | VNS |
|---|---|---|---|---|---|---|---|
| #1 | F | 37 | 31 | Genetic 1 | GE | Normal | Yes |
| #2 | F | 18 | 12 | LGS | GE | Dysplasia LF | No |
| #3 | M | 30 | 23 | Structural | GE/EE | Dysplasia biFT | Yes |
| #4 | M | 34 | 27 | Genetic 2 | EG/EE | Normal | Yes |
| #5 | M | 27 | 27 | LGS | GE | Normal | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pastor, J.; Vega-Zelaya, L. Features of Action Potentials from Identified Thalamic Nuclei in Anesthetized Patients. Brain Sci. 2020, 10, 1002. https://doi.org/10.3390/brainsci10121002
Pastor J, Vega-Zelaya L. Features of Action Potentials from Identified Thalamic Nuclei in Anesthetized Patients. Brain Sciences. 2020; 10(12):1002. https://doi.org/10.3390/brainsci10121002
Chicago/Turabian StylePastor, Jesús, and Lorena Vega-Zelaya. 2020. "Features of Action Potentials from Identified Thalamic Nuclei in Anesthetized Patients" Brain Sciences 10, no. 12: 1002. https://doi.org/10.3390/brainsci10121002
APA StylePastor, J., & Vega-Zelaya, L. (2020). Features of Action Potentials from Identified Thalamic Nuclei in Anesthetized Patients. Brain Sciences, 10(12), 1002. https://doi.org/10.3390/brainsci10121002