Patterns of Intrahemispheric EEG Asymmetry in Insomnia Sufferers: An Exploratory Study



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Participants
2.3. Research Protocol
2.4. Polysomnographic Montage
2.5. Power Spectral Analysis (PSA)
2.6. Statistical Aanalyses
2.6.1. Sample Size
2.6.2. Socio-Demographic Characteristics and Psychological Measures (Group Effects)
2.6.3. Sleep Measures (Group Effects)
2.6.4. Intrahemispheric Asymmetry (Group Effects)
2.6.5. Asymmetry and Misperception (Exploratory Correlations)
2.6.6. Asymmetry and Psychological Measures (Exploratory Correlations)
3. Results
3.1. Socio-Demographic Characteristics
3.2. Sleep Outcomes
3.2.1. Sleep Continuity
3.2.2. Sleep Macrostructure
3.3. Asymmetry Measures
3.3.1. Fronto-Central Regions
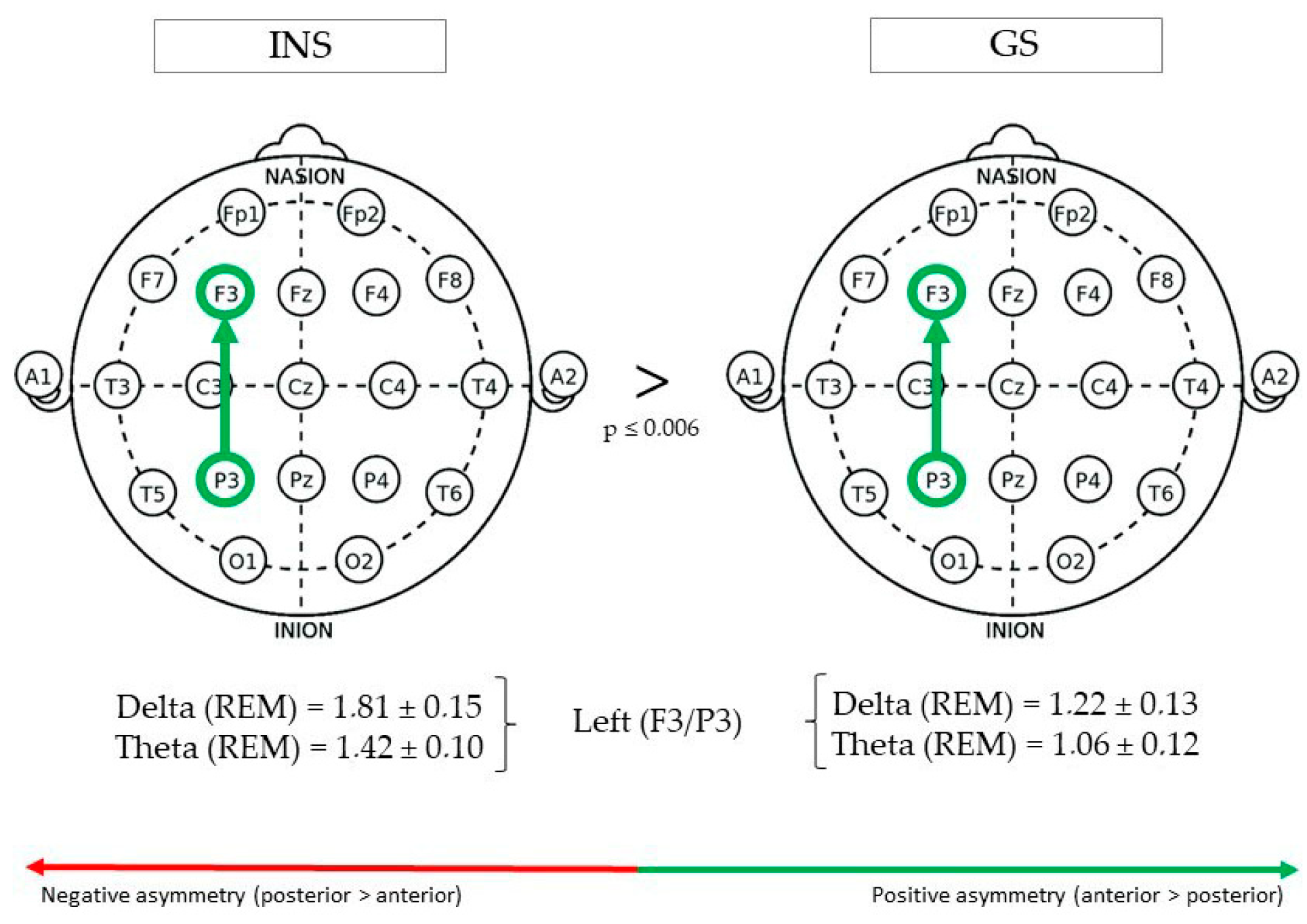
3.3.2. Fronto-Parietal Regions
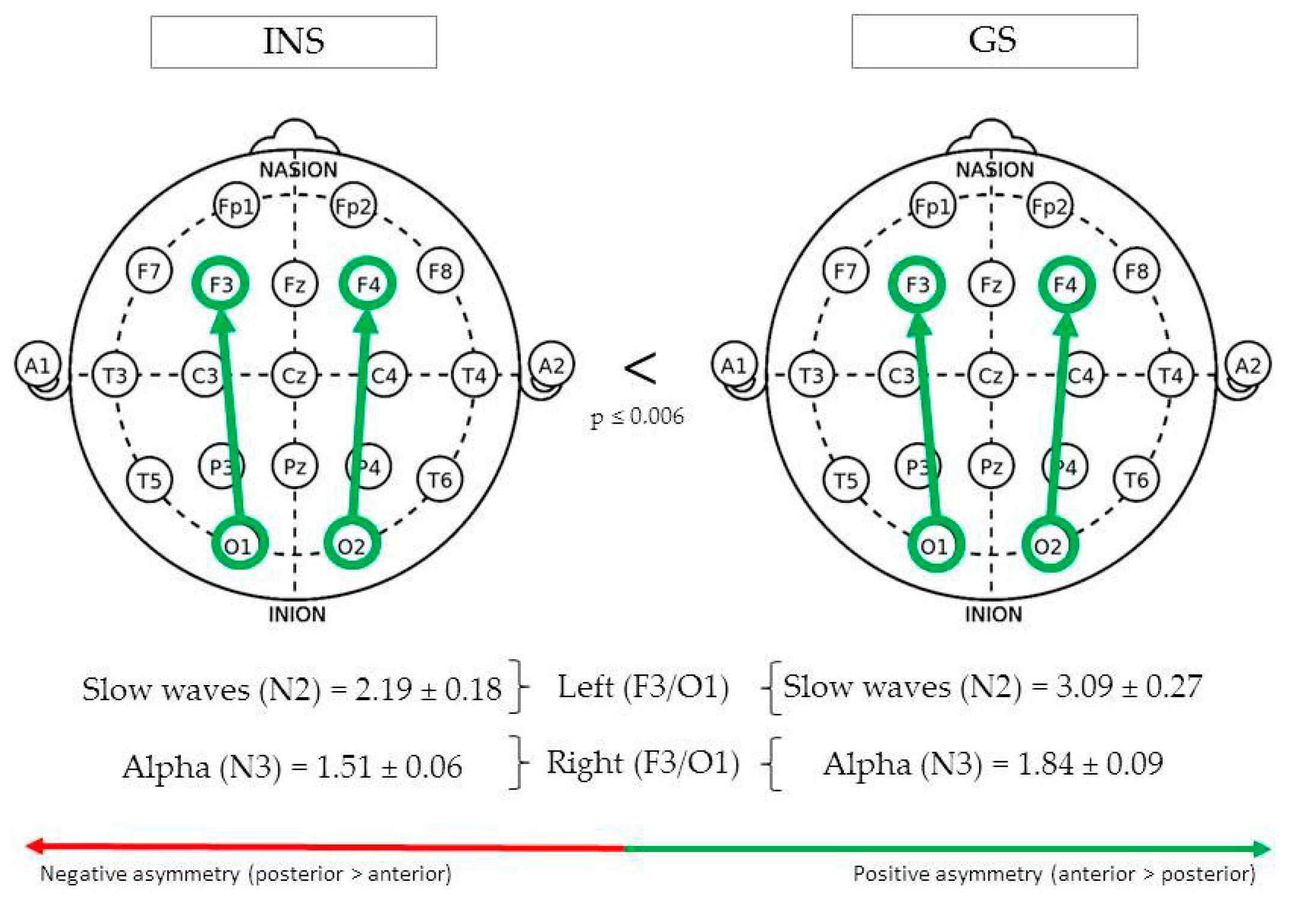
3.3.3. Fronto-Occipital Regions
3.3.4. Centro-Parietal Regions
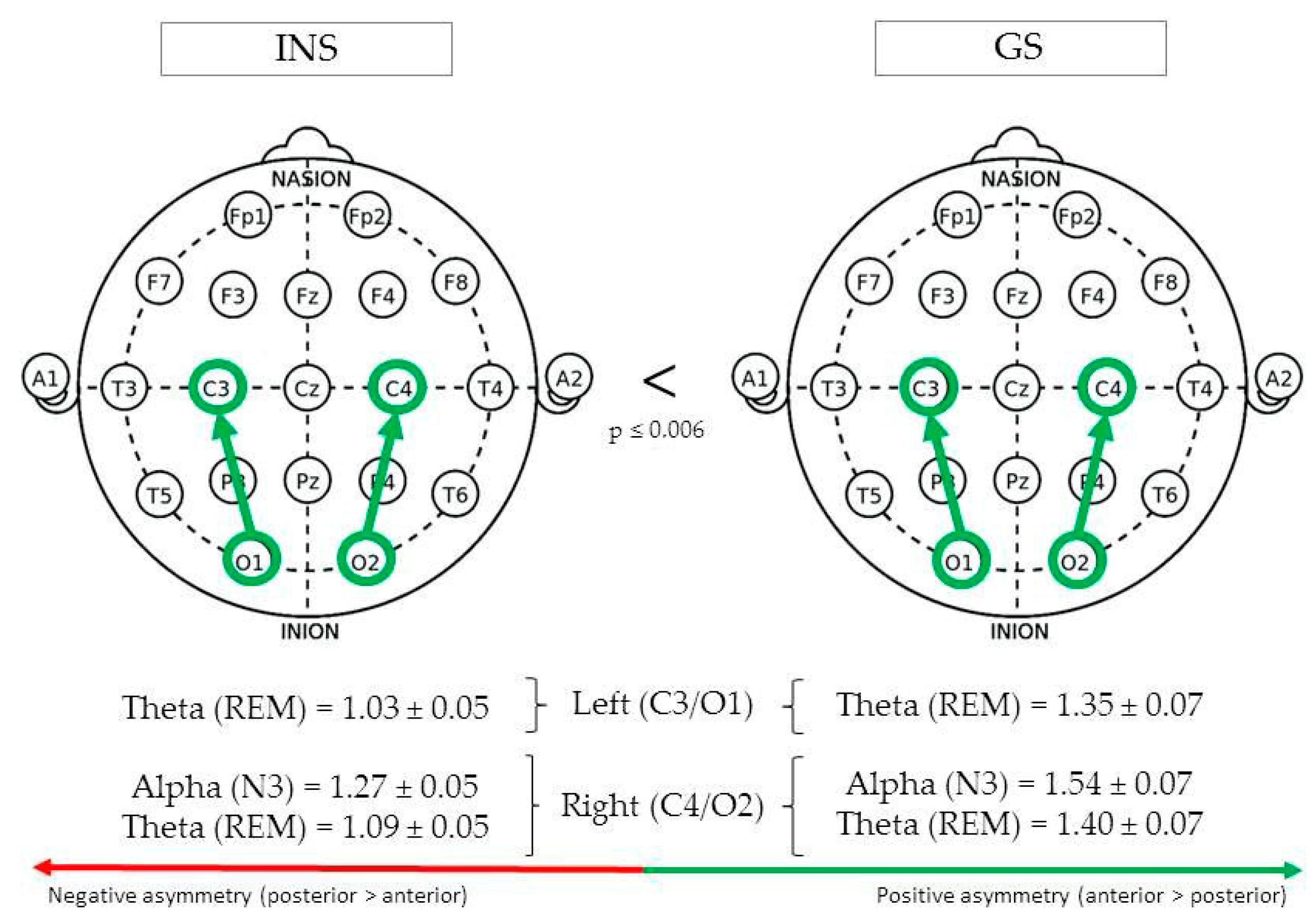
3.3.5. Centro-Occipital Regions
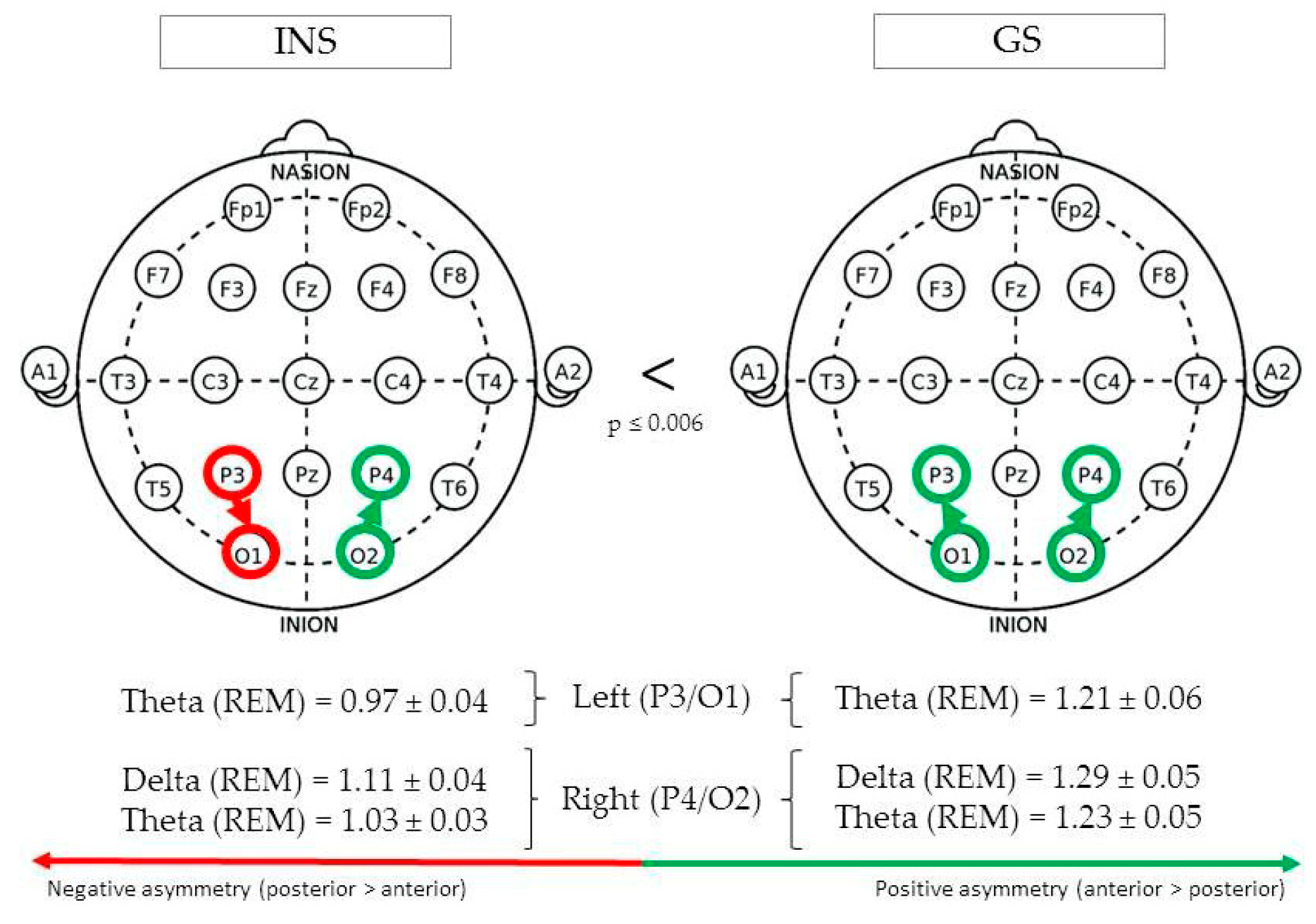
3.3.6. Parieto-Occipital Regions
3.4. Correlations between Asymmetry Measures and Sleep-Wake Misperception
3.4.1. Sleep-Onset Latency (SOL)
3.4.2. Wake after Sleep Onset (WASO)
3.4.3. Total Sleep Time (TST)
3.4.4. Total Wake Time (TWT)
3.5. Correlations between Asymmetry Measures and Clinical Symptoms
3.5.1. Beck Depression Inventory (BDI)
3.5.2. Beck Anxiety Inventory (BAI)
3.5.3. Insomnia Severity Index (ISI)
4. Discussion
4.1. Summary of Main Findings and Comparison With Existing Literature
4.1.1. Fronto-Parietal Derivations
4.1.2. Centro-Occipital and Parieto-Occipital Derivations
4.2. Strengths and Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-5®); American Psychiatric Association: Arlington, VA, USA, 2013. [Google Scholar]
- Freedman, R.R. EEG power spectra in sleep-onset insomnia. Electroencephalogr. Clin. Neurophysiol. 1986, 63, 408–413. [Google Scholar] [CrossRef]
- Merica, H.; Blois, R.; Gaillard, J.-M. Spectral characteristics of sleep EEG in chronic insomnia. Eur. J. Neurosci. 1998, 10, 1826–1834. [Google Scholar] [CrossRef] [PubMed]
- Perlis, M.L.; Smith, M.T.; Andrews, P.J.; Orff, H.; Giles, D.E. Beta/Gamma EEG Activity in Patients with Primary and Secondary Insomnia and Good Sleeper Controls. Sleep 2001, 24, 110–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perlis, M.L.; Mericab, H.; Smith, M.T.; Giles, D.E. Beta EEG activity and insomnia. Sleep Med. Rev. 2001, 5, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Spiegelhalder, K.; Regen, W.; Feige, B.; Holz, J.; Piosczyk, H.; Baglioni, C.; Riemann, D.; Nissen, C. Increased EEG sigma and beta power during NREM sleep in primary insomnia. Biol. Psychol. 2012, 91, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Krystal, A.D.; Edinger, J.D.; Wohlgemuth, W.K.; Marsh, G.R. NREM Sleep EEG Frequency Spectral Correlates of Sleep Complaints in Primary Insomnia Subtypes. Sleep 2002, 25, 626–636. [Google Scholar] [CrossRef] [Green Version]
- Buysse, D.J.; Germain, A.; Hall, M.L.; Moul, D.E.; Nofzinger, E.A.; Begley, A.; Ehlers, C.L.; Thompson, W.; Kupfer, D.J. EEG Spectral Analysis in Primary Insomnia: NREM Period Effects and Sex Differences. Sleep 2008, 31, 1673–1682. [Google Scholar] [CrossRef] [Green Version]
- Bastien, C.H. Insomnia: Neurophysiological and NeuropsychologicalApproaches. Neuropsychol. Rev. 2011, 21, 22–40. [Google Scholar] [CrossRef]
- Riemann, D.; Spiegelhalder, K.; Nissen, C.; Hirscher, V.; Baglioni, C.; Feige, B. REM sleep instability—A new pathway for insomnia? Pharmacopsychiatry 2012, 45, 167–176. [Google Scholar] [CrossRef]
- Pesonen, A.-K.; Gradisar, M.; Kuula, L.; Short, M.; Merikanto, I.; Tark, R.; Räikkönen, K.; Lahti, J. REM sleep fragmentation associated with depressive symptoms and genetic risk for depression in a community-based sample of adolescents. J. Affect. Disord. 2019, 245, 757–763. [Google Scholar] [CrossRef] [Green Version]
- Jansson-Fröjmark, M.; Lindblom, K. A bidirectional relationship between anxiety and depression, and insomnia? A prospective study in the general population. J. Psychosom. Res. 2008, 64, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Hoehn-Saric, R. Faculty Opinions recommendation of Worry, generalized anxiety disorder, and emotion: Evidence from the EEG gamma band. Fac. Opin. Post Publ. Peer Rev. Biomed. Lit. 2009, 79, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Cea-Cañas, B.; Gomez-Pilar, J.; Núñez, P.; Rodríguez-Vázquez, E.; De Uribe, N.; Díez, Á.; Pérez-Escudero, A.; Molina, V. Connectivity strength of the EEG functional network in schizophrenia and bipolar disorder. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2020, 98, 109801. [Google Scholar] [CrossRef]
- Goldschmied, J.R.; Cheng, P.; Armitage, R.; Deldin, P.J. A preliminary investigation of the role of slow-wave activity in modulating waking EEG theta as a marker of sleep propensity in major depressive disorder. J. Affect. Disord. 2019, 257, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Van Der Vinne, N.; Vollebregt, M.A.; Van Putten, M.J.; Arns, M. Frontal alpha asymmetry as a diagnostic marker in depression: Fact or fiction? A meta-analysis. NeuroImage Clin. 2017, 16, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.J.B.; Reznik, S.J. Frontal EEG asymmetry as a promising marker of depression vulnerability: Summary and methodological considerations. Curr. Opin. Psychol. 2015, 4, 93–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diego, M.A.; Field, T.; Hernandez-Reif, M. CES-D depression scores are correlated with frontal EEG alpha asymmetry. Depress. Anxiety 2001, 13, 32–37. [Google Scholar] [CrossRef]
- Grünewald, B.D.; Greimel, E.; Trinkl, M.; Bartling, J.; Großheinrich, N.; Schulte-Körne, G. Resting frontal EEG asymmetry patterns in adolescents with and without major depression. Biol. Psychol. 2018, 132, 212–216. [Google Scholar] [CrossRef]
- Kang, S.-G.; Mariani, S.; Marvin, S.A.; Ko, K.-P.; Redline, S.; Winkelman, J.W. Sleep EEG spectral power is correlated with subjective-objective discrepancy of sleep onset latency in major depressive disorder. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2018, 85, 122–127. [Google Scholar] [CrossRef]
- Thibodeau, R.; Jorgensen, R.S.; Kim, S. Depression, anxiety, and resting frontal EEG asymmetry: A meta-analytic review. J. Abnorm. Psychol. 2006, 115, 715–729. [Google Scholar] [CrossRef] [Green Version]
- Adolph, D.; Margraf, J. The differential relationship between trait anxiety, depression, and resting frontal α-asymmetry. J. Neural Transm. 2016, 124, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.F.; Kan, D.P.X.; Croarkin, P.; Phang, C.K.; Doruk, D. Neurophysiological correlates of depressive symptoms in young adults: A quantitative EEG study. J. Clin. Neurosci. 2018, 47, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Metzger, L.J.; Paige, S.R.; Carson, M.A.; Lasko, N.B.; Paulus, L.A.; Pitman, R.K.; Orr, S.P. PTSD Arousal and Depression Symptoms Associated With Increased Right-Sided Parietal EEG Asymmetry. J. Abnorm. Psychol. 2004, 113, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.L.; Towers, D.N.; Coan, J.A.; Allen, J.J.B. The oft-neglected role of parietal EEG asymmetry and risk for major depressive disorder. Psychophysiology 2011, 48, 82–95. [Google Scholar] [CrossRef] [Green Version]
- Nusslock, R.; Shackman, A.J.; McMenamin, B.W.; Greischar, L.L.; Davidson, R.J.; Kovacs, M. Comorbid anxiety moderates the relationship between depression history and prefrontal EEG asymmetry. Psychophysiology 2018, 55, e12953. [Google Scholar] [CrossRef] [Green Version]
- Ohayon, M.; Caulet, M.; Lemoine, P. Comorbidity of mental and insomnia disorders in the general population. Compr. Psychiatry 1998, 39, 185–197. [Google Scholar] [CrossRef]
- St-Jean, G.; Turcotte, I.; Bastien, C.H. Cerebral Asymmetry in Insomnia Sufferers. Front. Neurol. 2012, 3, 47. [Google Scholar] [CrossRef] [Green Version]
- Davidson, R.J. EEG Measures of Cerebral Asymmetry: Conceptual and Methodological Issues. Int. J. Neurosci. 1988, 39, 71–89. [Google Scholar] [CrossRef]
- Corsi-Cabrera, M.; Guevara, M.A.; Arce, C.; Ramos, J. Inter and intrahemispheric EEG correlation as a function of sleep cycles. Prog. Neuro Psychopharmacol. Biol. Psychiatry 1996, 20, 387–405. [Google Scholar] [CrossRef]
- Benca, R.M.; Obermeyer, W.H.; Larson, C.L.; Yun, B.; Dolski, I.; Kleist, K.D.; Weber, S.M.; Davidson, R.J. EEG alpha power and alpha power asymmetry in sleep and wakefulness. Psychophysiology 1999, 36, 430–436. [Google Scholar]
- Ferrara, M.; De Gennaro, L.; Curcio, G.; Cristiani, R.; Bertini, M. Interhemispheric asymmetry of human sleep EEG in response to selective slow-wave sleep deprivation. Behav. Neurosci. 2002, 116, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Kovrov, G.V.; Posokhov, S.I.; Strygin, K.N. Interhemispheric EEG asymmetry in patients with insomnia during nocturnal sleep. Bull. Exp. Biol. Med. 2006, 141, 197–199. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.; Buysse, D.J.; Nowell, P.D.; Nofzinger, E.A.; Houck, P.; Reynolds, C.F., 3rd; Kupfer, D.J. Symptoms of stress and depression as correlates of sleep in primary insomnia. Psychosom. Med. 2000, 62, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.A.; Wei, Y.; Ramautar, J.R.; Linkenkaer-Hansen, K.; Tagliazucchi, E.; Van Someren, E.J.W. More Severe Insomnia Complaints in People with Stronger Long-Range Temporal Correlations in Wake Resting-State EEG. Front. Physiol. 2016, 7, 576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mennella, R.; Patron, E.; Palomba, D. Frontal alpha asymmetry neurofeedback for the reduction of negative affect and anxiety. Behav. Res. Ther. 2017, 92, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Frase, L.; Piosczyk, H.; Zittel, S.; Jahn, F.; Selhausen, P.; Krone, L.; Feige, B.; Mainberger, F.; Maier, J.G.; Kuhn, M.; et al. Modulation of Total Sleep Time by Transcranial Direct Current Stimulation (tDCS). Neuropsychopharmacology 2016, 41, 2577–2586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frase, L.; Selhausen, P.; Krone, L.B.; Tsodor, S.; Jahn, F.; Feige, B.; Maier, J.G.; Mainberger, F.; Piosczyk, H.; Kuhn, M.; et al. Differential effects of bifrontal tDCS on arousal and sleep duration in insomnia patients and healthy controls. Brain Stimul. 2019, 12, 674–683. [Google Scholar] [CrossRef]
- Saebipour, M.R.; Joghataei, M.T.; Yoonessi, A.; Sadeghniiat-Haghighi, K.; Khalighinejad, N.; Khademi, S. Slow oscillating transcranial direct current stimulation during sleep has a sleep-stabilizing effect in chronic insomnia: A pilot study. J. Sleep Res. 2015, 24, 518–525. [Google Scholar] [CrossRef]
- Cervena, K.; Dauvilliers, Y.; Espa, F.; Touchon, J.; Matousek, M.; Billiard, M.; Besset, A. Effect of cognitive behavioural therapy for insomnia on sleep architecture and sleep EEG power spectra in psychophysiological insomnia. J. Sleep Res. 2004, 13, 385–393. [Google Scholar] [CrossRef]
- St-Jean, G.; Turcotte, I.; Pérusse, A.D.; Bastien, C.H. REM and NREM power spectral analysis on two consecutive nights in psychophysiological and paradoxical insomnia sufferers. Int. J. Psychophysiol. 2013, 89, 181–194. [Google Scholar] [CrossRef]
- Poyares, D.; Guilleminault, C.; Ohayon, M.M.; Tufik, S. Chronic benzodiazepine usage and withdrawal in insomnia patients. J. Psychiatr. Res. 2004, 38, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Bastien, C.H. Validation of the Insomnia Severity Index as an outcome measure for insomnia research. Sleep Med. 2001, 2, 297–307. [Google Scholar] [CrossRef]
- Beck, A.T.; Epstein, N.; Brown, G.; Steer, R.A. An inventory for measuring clinical anxiety: Psychometric properties. J. Consult. Clin. Psychol. 1988, 56, 893–897. [Google Scholar] [CrossRef] [PubMed]
- Beck, A.T.; Steer, R.A. Internal consistencies of the original and revised beck depression inventory. J. Clin. Psychol. 1984, 40, 1365–1367. [Google Scholar] [CrossRef]
- Morin, C.M. Insomnia: Psychological Assessment and Management; Guilford Press: New York, NY, USA, 1993. [Google Scholar]
- Spitzer, R.L.; Williams, J.B.; Gibbon, M.; First, M.B. The Structured Clinical Interview for DSM-III-R (SCID). I: History, rationale, and description. Arch. Gen. Psychiatry 1992, 49, 624–629. [Google Scholar] [CrossRef] [PubMed]
- Pivik, R.T.; Broughton, R.J.; Coppola, R.; Davidson, R.J.; Fox, N.; Nuwer, M.R. Guidelines for the recording and quantitative analysis of electroencephalographic activity in research contexts. Psychophysiology 1993, 30, 547–558. [Google Scholar] [CrossRef]
- Rechtschaffen, A.; Kales, A. A Manual of Standardized Terminology, Techniques and Scoring System of Sleep Stages in Human Subjects; Brain Information Service/Brain Research Institute, University of California: Los Angeles, CA, USA, 1968. [Google Scholar]
- Brunner, D.P.; Vasko, R.C.; Detka, C.S.; Monahan, J.P.; Reynolds, C.F., 3rd; Kupfer, D.J. Muscle artifacts in the sleep EEG: Automated detection and effect on all-night EEG power spectra. J. Sleep Res. 1996, 5, 155–164. [Google Scholar]
- Campbell, I.G. EEG Recording and Analysis for Sleep Research. Curr. Protoc. Neurosci. 2009, 49, 10.2.1–10.2.19. [Google Scholar] [CrossRef] [Green Version]
- Corsi-Cabrera, M.; Arce, C.; Ramos, J.; Guevara, M.A. Effect of spatial ability and sex on inter- and intrahemispheric correlation of EEG activity. Electroencephalogr. Clin. Neurophysiol. 1997, 102, 5–11. [Google Scholar] [CrossRef]
- Corsi-Cabrera, M.; Figueredo-Rodríguez, P.; Del Río-Portilla, Y.; Sánchez-Romero, J.; Galán, L.; Bosch-Bayard, J. Enhanced Frontoparietal Synchronized Activation During the Wake-Sleep Transition in Patients with Primary Insomnia. Sleep 2012, 35, 501–511. [Google Scholar] [CrossRef] [Green Version]
- Corsi-Cabrera, M.; Miró, E.; Del-Río-Portilla, Y.; Pérez-Garci, E.; Villanueva, Y.; Guevara, M.A. Rapid eye movement sleep dreaming is characterized by uncoupled EEG activity between frontal and perceptual cortical regions. Brain Cogn. 2003, 51, 337–345. [Google Scholar] [CrossRef]
- Corsi-Cabrera, M.; Sifuentes-Ortega, R.; Rosales-Lagarde, A.; Rojas-Ramos, O.A.; Del-Rı#xo-Portilla, Y. Enhanced synchronization of gamma activity between frontal lobes during REM sleep as a function of REM sleep deprivation in man. Exp. Brain Res. 2014, 232, 1497–1508. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, D.; Jung, S.; Saadat, M.; Sirohi, R.; Crewson, K. How to interpret the results of a sleep study. J. Community Hosp. Intern. Med. Perspect. 2014, 4, 24983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzbani, H.; Marateb, H.R.; Mansourian, M. Methodological Note: Neurofeedback: A Comprehensive Review on System Design, Methodology and Clinical Applications. Basic Clin. Neurosci. J. 2015, 7, 143–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Provencher, T.; De Psychologie, U.L.É.; Charest, J.; Bastien, C.H. Non-Invasive Brain Stimulation for Insomnia—A Review of Current Data and Future Implications. OBM Integr. Complement. Med. 2019, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Merica, H.; Gaillard, J.-M. The EEG of the sleep onset period in insomnia: A discriminant analysis. Physiol. Behav. 1992, 52, 199–204. [Google Scholar] [CrossRef]
- Picchioni, D.; Fukunaga, M.; Carr, W.S.; Braun, A.R.; Balkin, T.J.; Duyn, J.H.; Horovitz, S.G. fMRI differences between early and late stage-1 sleep. Neurosci. Lett. 2008, 441, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Untergehrer, G.; Jordan, D.; Kochs, E.F.; Ilg, R.; Schneider, G. Fronto-Parietal Connectivity Is a Non-Static Phenomenon with Characteristic Changes during Unconsciousness. PLoS ONE 2014, 9, e87498. [Google Scholar] [CrossRef] [Green Version]
- Grimshaw, G.; Foster, J.J.; Corballis, P.M. Frontal and parietal EEG asymmetries interact to predict attentional bias to threat. Brain Cogn. 2014, 90, 76–86. [Google Scholar] [CrossRef]
- Marchetti, L.M.; Biello, S.; Broomfield, N.M.; MacMahon, K.M.A.; Espie, C.A. Who is pre-occupied with sleep? A comparison of attention bias in people with psychophysiological insomnia, delayed sleep phase syndrome and good sleepers using the induced change blindness paradigm. J. Sleep Res. 2006, 15, 212–221. [Google Scholar] [CrossRef]
- Jones, B.T.; Macphee, L.M.; Broomfield, N.M.; Jones, B.C.; Espie, C.A. Sleep-Related Attentional Bias in Good, Moderate, and Poor (Primary Insomnia) Sleepers. J. Abnorm. Psychol. 2005, 114, 249–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simor, P.; Gombos, F.; Blaskovich, B.; Bódizs, R. Long-range alpha and beta and short-range gamma EEG synchronization distinguishes phasic and tonic REM periods. Sleep 2017, 41. [Google Scholar] [CrossRef] [PubMed]
- Hasan, J. Automatic analysis of sleep recordings: A critical review. Ann. Clin. Res. 1985, 17, 280–287. [Google Scholar] [PubMed]
- Waterman, D.; Woestenburg, J.C.; Elton, M.; Hofman, W.; Kok, A. Removal of Ocular Artifacts From the REM Sleep EEG. Sleep 1992, 15, 371–375. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Zhang, Y.; Chen, J.; Xie, C.; Gan, R.; Wang, L.; Wang, L. Changes in theta activities in the left posterior temporal region, left occipital region and right frontal region related to mild cognitive impairment in Parkinson’s disease patients. Int. J. Neurosci. 2017, 127, 66–72. [Google Scholar]
- Coben, L.A.; Chi, D.; Snyder, A.Z.; Storandt, M. Replication of a study of frequency analysis of the resting awake EEG in mild probable Alzheimer’s disease. Electroencephalogr. Clin. Neurophysiol. 1990, 75, 148–154. [Google Scholar] [CrossRef]
- Coben, L.A.; Danziger, W.; Storandt, M. A longitudinal EEG study of mild senile dementia of Alzheimer type: Changes at 1 year and at 2.5 years. Electroencephalogr. Clin. Neurophysiol. 1985, 61, 101–112. [Google Scholar] [CrossRef]
- Penttila, M.; Partanen, J.V.; Soininen, H.; Riekkinen, P.J. Quantitative analysis of occipital EEG in different stages of Alzheimer’s disease. Electroencephalogr. Clin. Neurophysiol. 1985, 60, 1–6. [Google Scholar] [CrossRef]
- Hot, P.; Rauchs, G.; Bertran, F.; Denise, P.; Desgranges, B.; Clochon, P.; Eustache, F. Changes in sleep theta rhythm are related to episodic memory impairment in early Alzheimer’s disease. Biol Psychol. 2011, 87, 334–339. [Google Scholar] [CrossRef]
- Hankins, T.C.; Wilson, G.F. A comparison of heart rate, eye activity, EEG and subjective measures of pilot mental workload during flight. Aviat. Space Environ. Med. 1998, 69, 360–367. [Google Scholar]
- Schreiner, T.; Göldi, M.; Rasch, B. Cueing vocabulary during sleep increases theta activity during later recognition testing. Psychophysiology 2015, 52, 1538–1543. [Google Scholar] [CrossRef] [PubMed]
- Schreiner, T.; Rasch, B. Boosting Vocabulary Learning by Verbal Cueing During Sleep. Cereb. Cortex 2015, 25, 4169–4179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauseng, P.; Klimesch, W.; Doppelmayr, M.; Pecherstorfer, T.; Freunberger, R.; Hanslmayr, S. EEG alpha synchronization and functional coupling during top-down processing in a working memory task. Hum. Brain Mapp. 2005, 26, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, J.; Hwang, G.; Curran, T.; Kahana, M.J. EEG oscillations and recognition memory: Theta correlates of memory retrieval and decision making. NeuroImage 2006, 32, 978–987. [Google Scholar] [CrossRef] [PubMed]
- Reiner, M.; Rozengurt, R.; Barnea, A. Better than sleep: Theta neurofeedback training accelerates memory consolidation. Biol. Psychol. 2014, 95, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Fortier-Brochu, É.; Beaulieu-Bonneau, S.; Ivers, H.; Morin, C.M. Insomnia and daytime cognitive performance: A meta-analysis. Sleep Med. Rev. 2012, 16, 83–94. [Google Scholar] [CrossRef]
- Iber, C.; Ancoli-Israel, S.; Chesson, A.L.; Quan, S.F. The AASM Manual for the Scoring of Sleep and Associated Events: Rules, Terminology and Technical Specifications; American Academy of Sleep Medicine: Westchester, IL, USA, 2007. [Google Scholar]
- Arns, M.; Bruder, G.; Hegerl, U.; Spooner, C.; Palmer, D.M.; Etkin, A.; Gordon, E. EEG alpha asymmetry as a gender-specific predictor of outcome to acute treatment with different antidepressant medications in the randomized iSPOT-D study. Clin. Neurophysiol. 2016, 127, 509–519. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GS (n = 19) Mean (SD) | INS (n = 43) Mean (SD) | |
|---|---|---|
| Sex | ||
| Male | 9 | 16 |
| Female | 10 | 27 |
| Age (years) | 37.47 (9.75) | 41.05 (8.79) |
| Education (years) | 15.47 (4.01) | 15.37 (3.36) |
| Questionnaires | ||
| ISI | 2.00 (3.04) * | 16.98 (3.43) |
| BDI | 2.11 (3.43) * | 7.31 (4.99) |
| BAI | 1.22 (2.13) * | 7.33 (6.20) |
| GS | INS | Group Effect p | Hedge’s g | |
|---|---|---|---|---|
| SOL | ||||
| Objective | 10.82 ± 3.39 | 14.68 ± 2.27 | 0.3477 | 1.45 |
| Subjective | 17.13 ± 5.97 | 35.00 ± 4.02 | 0.0159 * | 3.81 |
| MI (subj./obj.) | 2.15 ± 0.50 | 5.22 ± 1.10∆ | 0.0135 * | 3.20 |
| WASO | ||||
| Objective | 26.84 ± 7.91 | 49.98 ± 5.30 | 0.0182 * | 3.73 |
| Subjective | 9.54 ± 9.81 | 59.03 ± 6.71 | 0.0001 * | 6.37 |
| MI (subj./obj.) | 0.51 ± 0.18 | 2.46 ± 0.59 | 0.0025 * | 3.87 |
| TST | ||||
| Objective | 408.33 ± 9.72 | 399.68 ± 6.54 | 0.4628 | 1.13 |
| Subjective | 432.51 ± 13.85 | 353.58 ± 9.34 | <0.0001 * | 7.25 |
| MI (subj./obj.) | 1.07 ± 0.03 | 0.89 ± 0.02∆ | <0.0001 * | 7.68 |
| TWT | ||||
| Objective | 23.53 ± 6.28 | 41.93 ± 4.24 | 0.0183 * | 3.72 |
| Subjective | 33.09 ± 12.48 | 118.04 ± 8.51 | <0.0001 * | 8.61 |
| MI (subj./obj.) | 2.76 ± 0.59 | 6.27 ± 0.96∆ | 0.0027 * | 4.05 |
| GS | INS | Multivariate Group Effect | ||
|---|---|---|---|---|
| F | p | |||
| Total Time (min) | ||||
| N1 | 13.89 ± 2.08 | 12.60 ± 1.39 | 0.23 | 0.875 |
| N2 | 251.91 ± 9.96 | 247.33 ± 6.67 | ||
| N3 | 38.99 ± 6.00 | 41.45 ± 4.01 | ||
| REM | 103.68 ± 5.99 | 98.14 ± 4.01 | ||
| Proportion (%) | ||||
| N1 | 3.56 ± 0.58 | 3.20 ± 0.39 | 0.28 | 0.888 |
| N2 | 61.72 ± 1.65 | 61.79 ± 1.11 | ||
| N3 | 9.46 ± 1.54 | 10.38 ± 1.03 | ||
| REM | 25.26 ± 1.30 | 24.63 ± 0.87 | ||
| Region | Hemisphere | Stage | Frequency | GS | INS | p | Hedges’ g |
|---|---|---|---|---|---|---|---|
| Fronto-central | Left (F3/C3) | ns | |||||
| Right (F4/C4) | ns | ||||||
| Mid (Fz/Cz) | ns | ||||||
| Fronto-parietal | Left (F3/P3) | REM | Delta | 1.22 ± 0.13 | 1.81 ± 0.15 | 0.005 | 4.08 |
| REM | Theta | 1.06 ± 0.12 | 1.42 ± 0.10 | 0.006 | 3.38 | ||
| Right (F4/P4) | ns | ||||||
| Mid (Fz/Pz) | ns | ||||||
| Fronto-occipital | Left (F3/O1) | N2 | Slow waves | 3.09 ± 0.27 | 2.19 ± 0.18 | 0.006 | 4.26 |
| Right (F4/O2) | N3 | Alpha | 1.84 ± 0.09 | 1.51 ± 0.06 | 0.005 | 4.69 | |
| Centro-parietal | Left (C3/P3) | ns | |||||
| Right (C4/P4) | ns | ||||||
| Mid (Cz/Pz) | ns | ||||||
| Centro-occipital | Left (C3/O1) | REM | Theta | 1.35 ± 0.07 | 1.03 ± 0.05 | <0.001 | 5.64 |
| Right (C4/O2) | N3 | Alpha | 1.54 ± 0.07 | 1.27 ± 0.05 | 0.003 | 4.76 | |
| REM | Theta | 1.40 ± 0.07 | 1.09 ± 0.05 | <0.001 | 5.46 | ||
| Parieto-occipital | Left (P3/O1) | REM | Theta | 1.21 ± 0.06 | 0.97 ± 0.04 | 0.001 | 5.12 |
| Right (P4/O2) | REM | Delta | 1.29 ± 0.05 | 1.11 ± 0.04 | 0.006 | 4.16 | |
| REM | Theta | 1.23 ± 0.05 | 1.03 ± 0.03 | 0.001 | 5.38 |
| Region | Stage | Frequency | SOL | WASO | TST | TWT | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| GS | INS | GS | INS | GS | INS | GS | INS | |||
| F3/P3 | REM | Delta | ns | ns | ns | ns | −0.324 * | ns | ns | ns |
| Theta | ns | ns | ns | ns | ns | ns | ns | ns | ||
| F3/O1 | N2 | Slow | ns | ns | ns | ns | ns | ns | ns | ns |
| F4/O2 | N3 | Alpha | 0.400 * | ns | ns | ns | ns | ns | ns | ns |
| C3/O1 | REM | Theta | 0.430 ** | ns | ns | ns | ns | ns | ns | ns |
| C4/O2 | N3 | Alpha | 0.410 * | ns | ns | ns | ns | ns | ns | ns |
| REM | Theta | 0.343 * | ns | ns | ns | ns | ns | ns | ns | |
| P3/O1 | REM | Theta | 0.325 * | ns | ns | ns | ns | ns | ns | ns |
| P4/O2 | REM | Delta | ns | −0.259 * | ns | ns | ns | 0.232 * | ns | ns |
| Theta | 0.401 * | ns | ns | ns | ns | ns | ns | ns | ||
| Region | Stage | Frequency | BDI | BAI | ISI | |||
|---|---|---|---|---|---|---|---|---|
| GS | INS | GS | GS | INS | GS | |||
| F3/P3 | REM | Delta | ns | 0.214 * | ns | ns | ns | ns |
| Theta | −0.559 ** | 0.205 * | ns | ns | −0.406 * | 0.182 * | ||
| F3/O1 | N2 | Slow | −0.488 ** | ns | ns | ns | ns | ns |
| F4/O2 | N3 | Alpha | −0.362 * | ns | ns | ns | ns | ns |
| C3/O1 | REM | Theta | −0.331 * | ns | ns | ns | ns | ns |
| C4/O2 | N3 | Alpha | ns | ns | ns | ns | ns | ns |
| REM | Theta | ns | ns | ns | ns | ns | ns | |
| P3/O1 | REM | Theta | ns | ns | ns | ns | ns | ns |
| P4/O2 | REM | Delta | ns | 0.263 * | ns | ns | ns | ns |
| Theta | ns | ns | ns | ns | ns | ns | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Provencher, T.; Fecteau, S.; Bastien, C.H. Patterns of Intrahemispheric EEG Asymmetry in Insomnia Sufferers: An Exploratory Study. Brain Sci. 2020, 10, 1014. https://doi.org/10.3390/brainsci10121014
Provencher T, Fecteau S, Bastien CH. Patterns of Intrahemispheric EEG Asymmetry in Insomnia Sufferers: An Exploratory Study. Brain Sciences. 2020; 10(12):1014. https://doi.org/10.3390/brainsci10121014
Chicago/Turabian StyleProvencher, Thierry, Shirley Fecteau, and Célyne H. Bastien. 2020. "Patterns of Intrahemispheric EEG Asymmetry in Insomnia Sufferers: An Exploratory Study" Brain Sciences 10, no. 12: 1014. https://doi.org/10.3390/brainsci10121014
APA StyleProvencher, T., Fecteau, S., & Bastien, C. H. (2020). Patterns of Intrahemispheric EEG Asymmetry in Insomnia Sufferers: An Exploratory Study. Brain Sciences, 10(12), 1014. https://doi.org/10.3390/brainsci10121014