
Chronic Mild Unpredictable Stress and High-Fat Diet Given during Adolescence Impact Both Cognitive and Noncognitive Behaviors in Young Adult Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Methods
2.1.1. Animals
2.1.2. Diets
2.1.3. Chronic Mild Unpredictable Stress (CMUS) Paradigm
2.1.4. Behavioral Tests
Open Field Test (OFT)
Elevated-Zero Maze (EZM)
Morris Water Maze (MWM)
Activities of Daily Living (ADLs): Burrowing
Activities of Daily Living (ADLs): Nesting
2.2. Statistical Analyses
3. Results
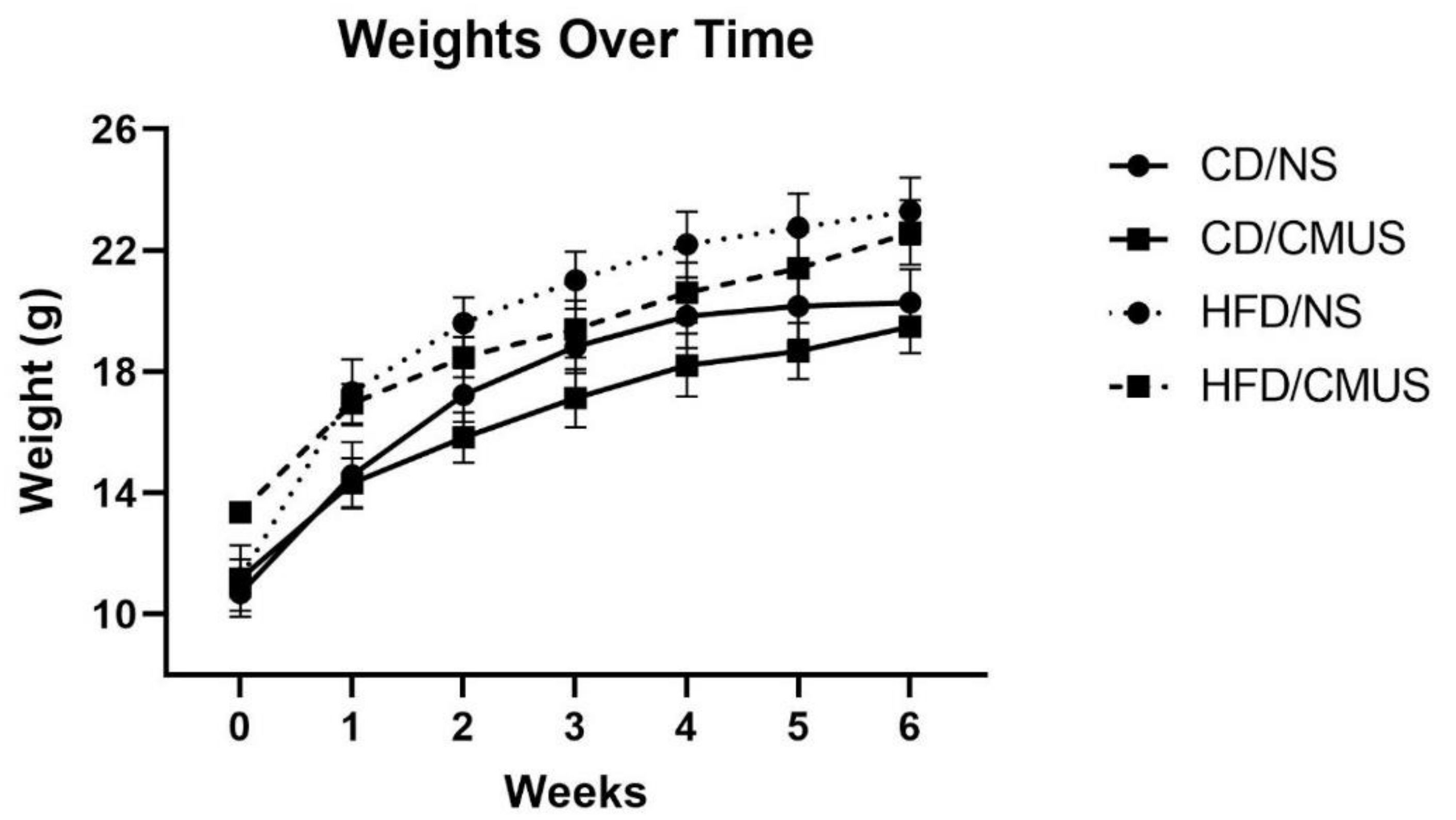
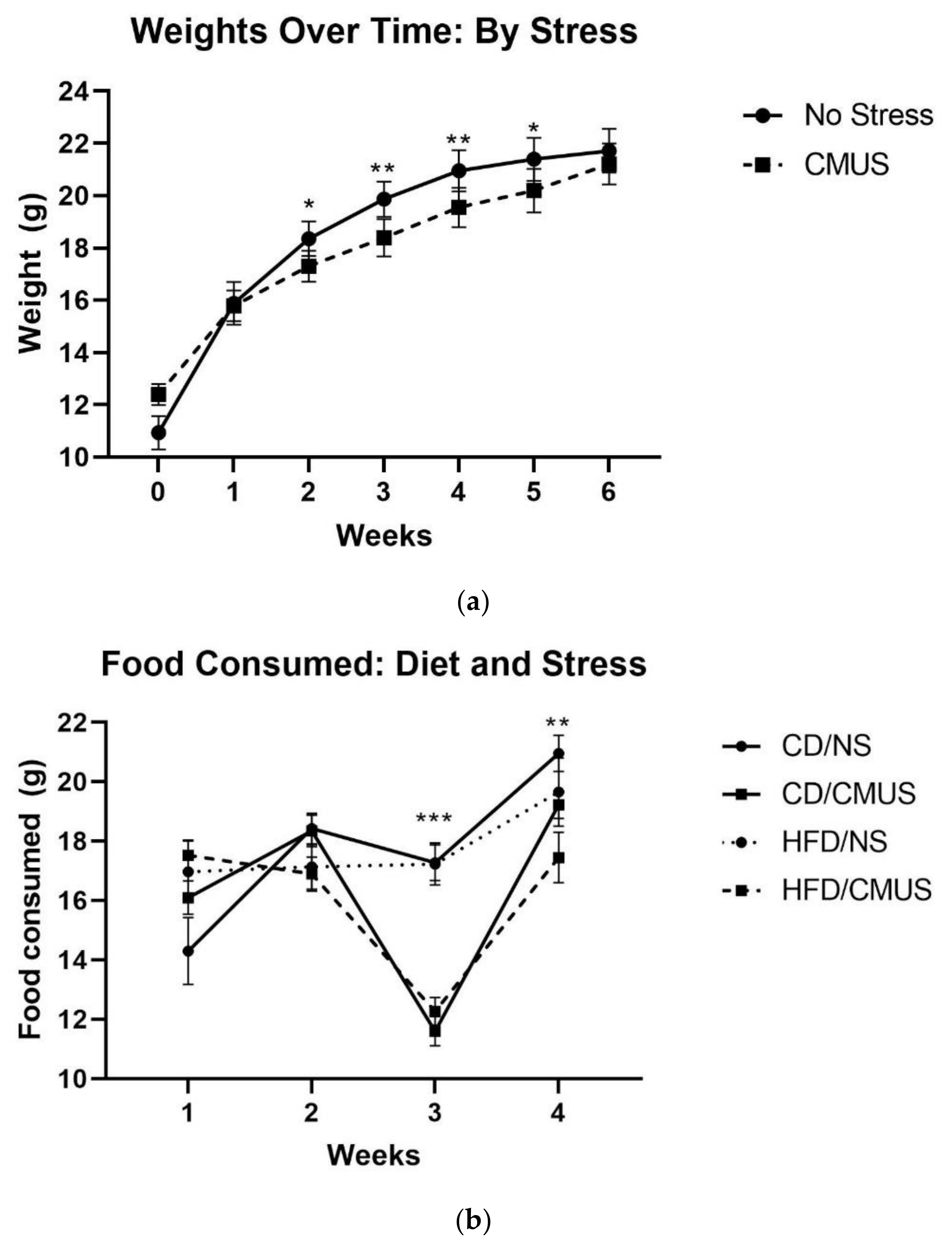
3.1. Animal Weights and Food Consumed
3.2. Open Field Test
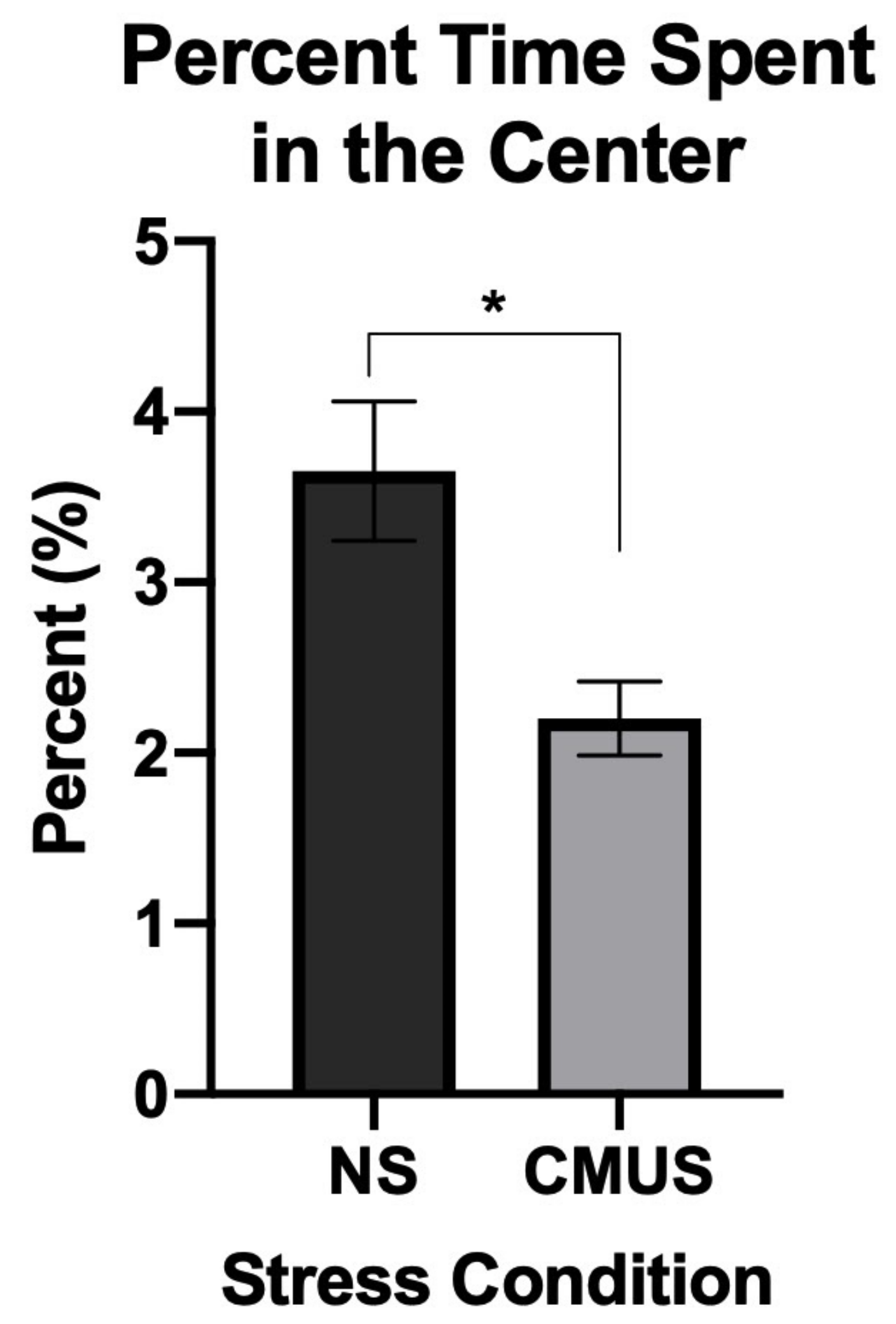
3.2.1. Percent Time in the Center
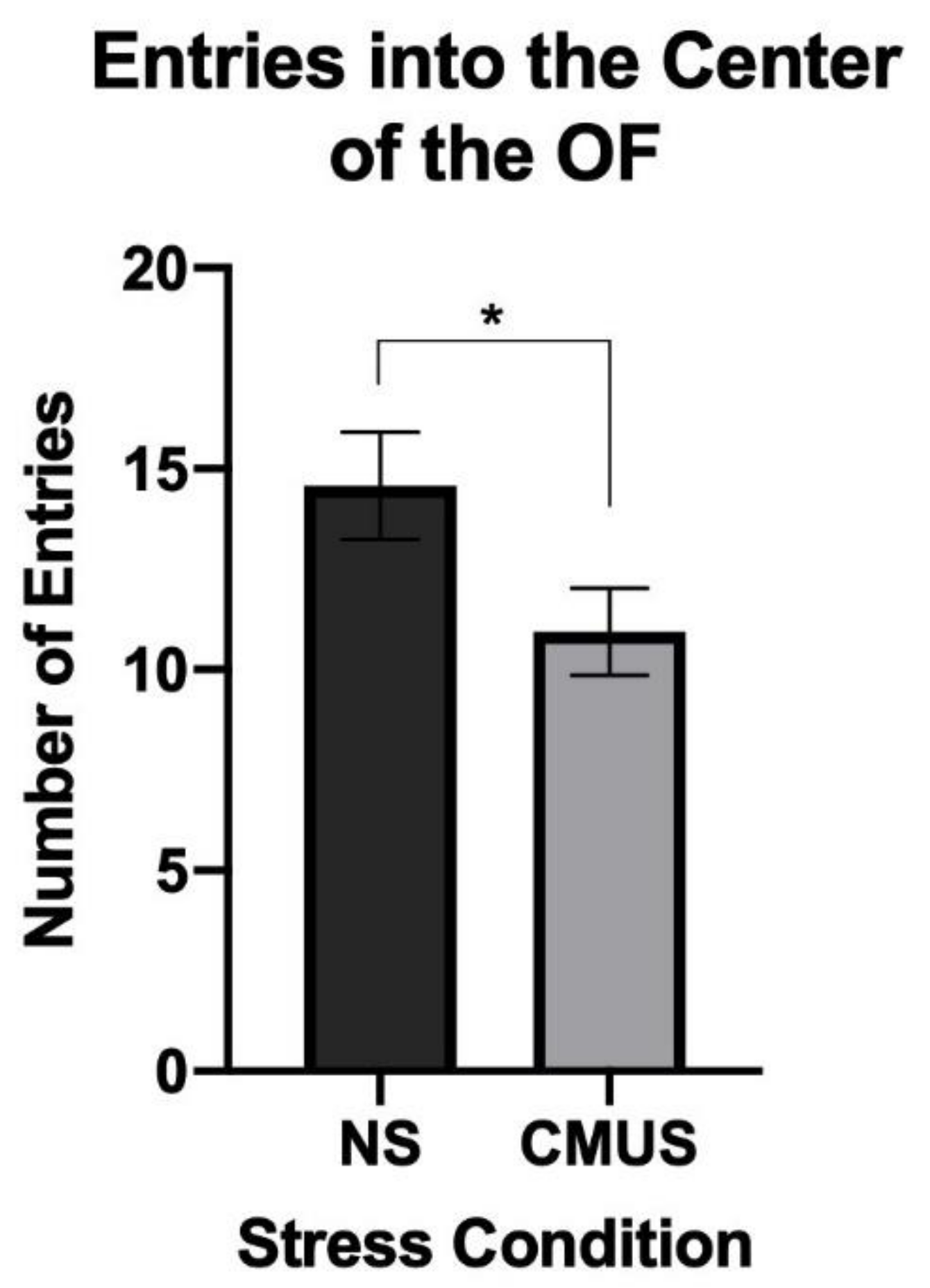
3.2.2. Center Entries
3.2.3. Latency to Enter the Center
3.2.4. Total Distance
3.3. Elevated Zero Maze (EZM)
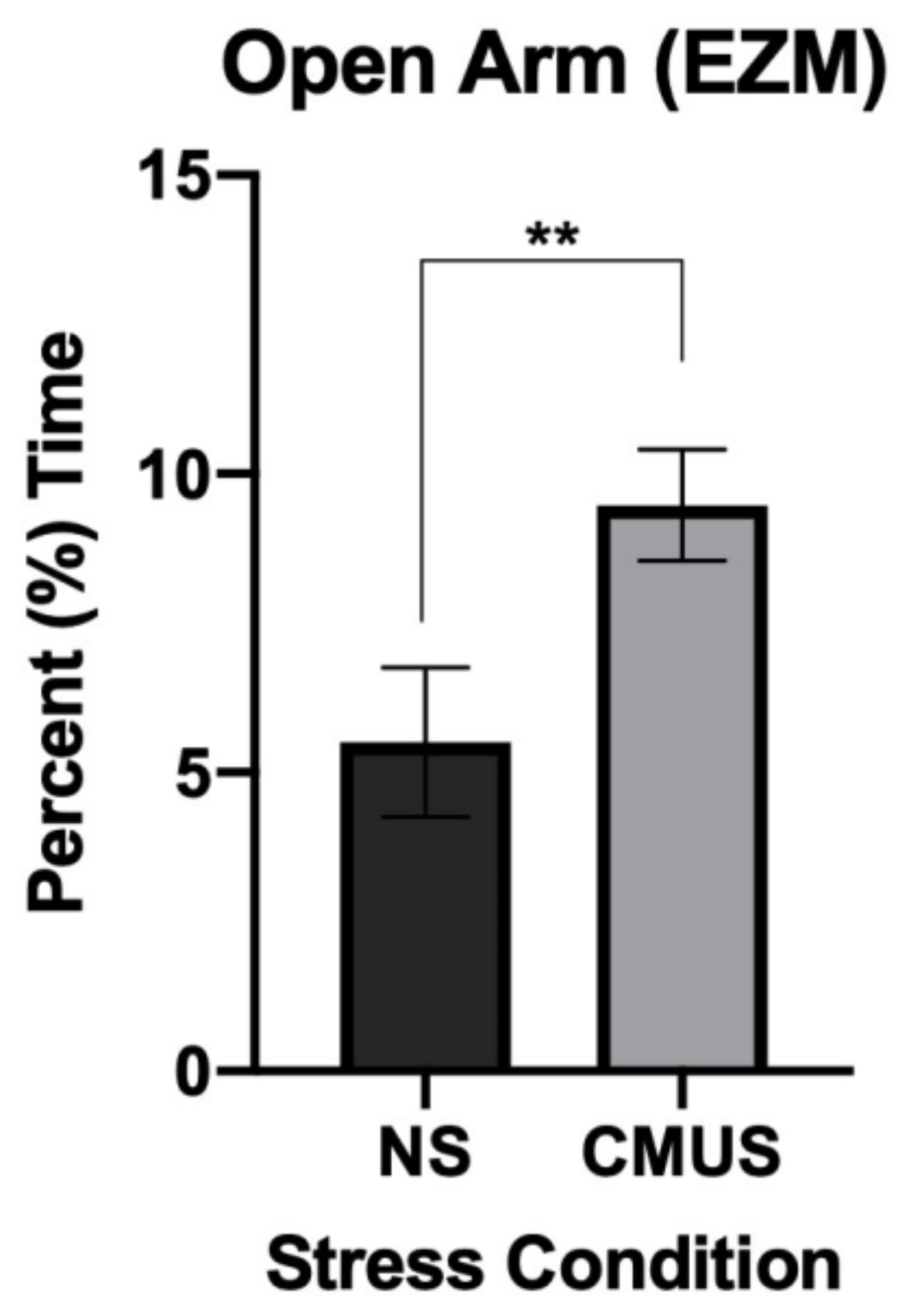
3.3.1. Percent Time in the Open Arm
3.3.2. Latency to Enter into the Open Arm
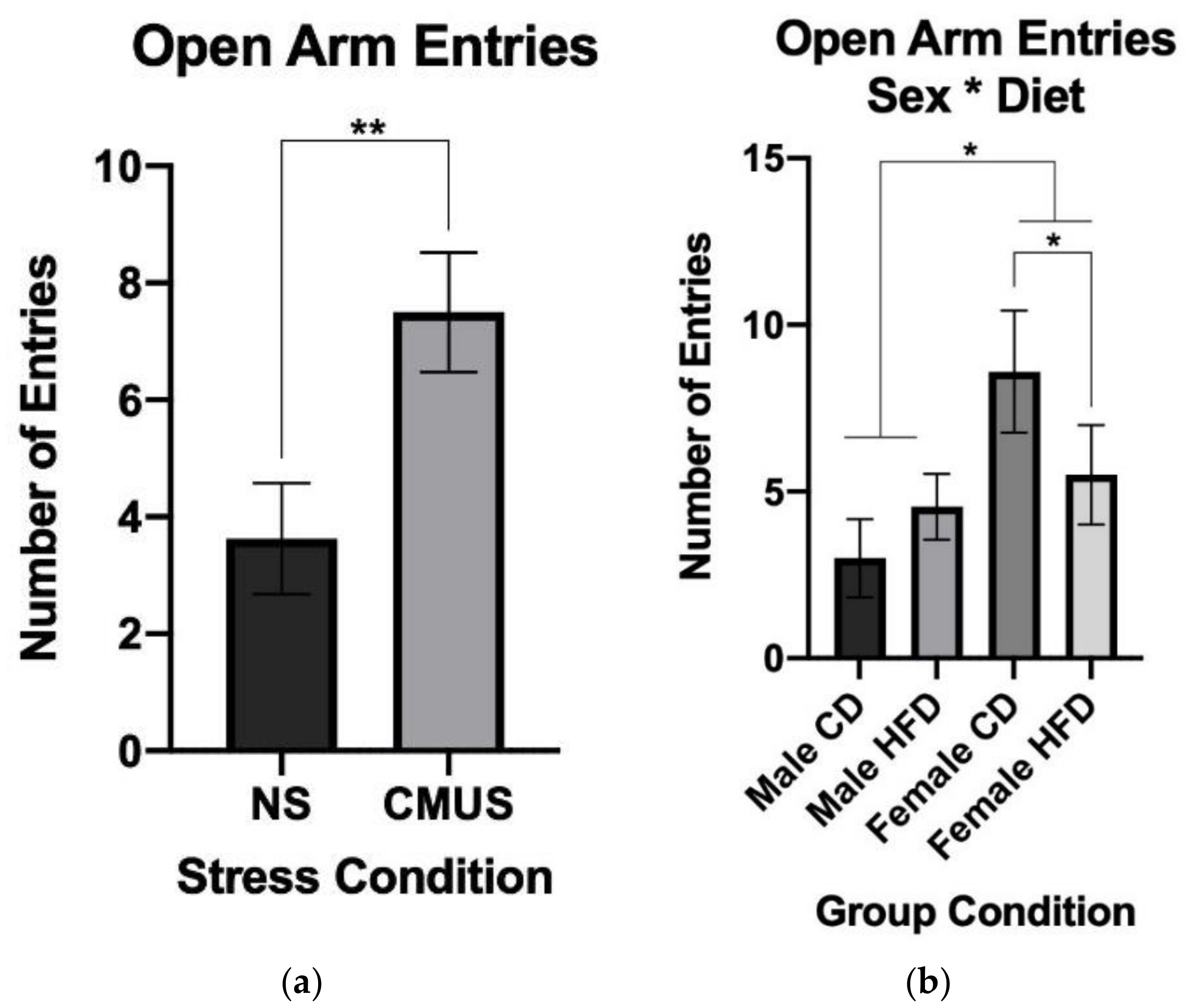
3.3.3. Open Arm Entries
3.4. Morris Water Maze (MWM)
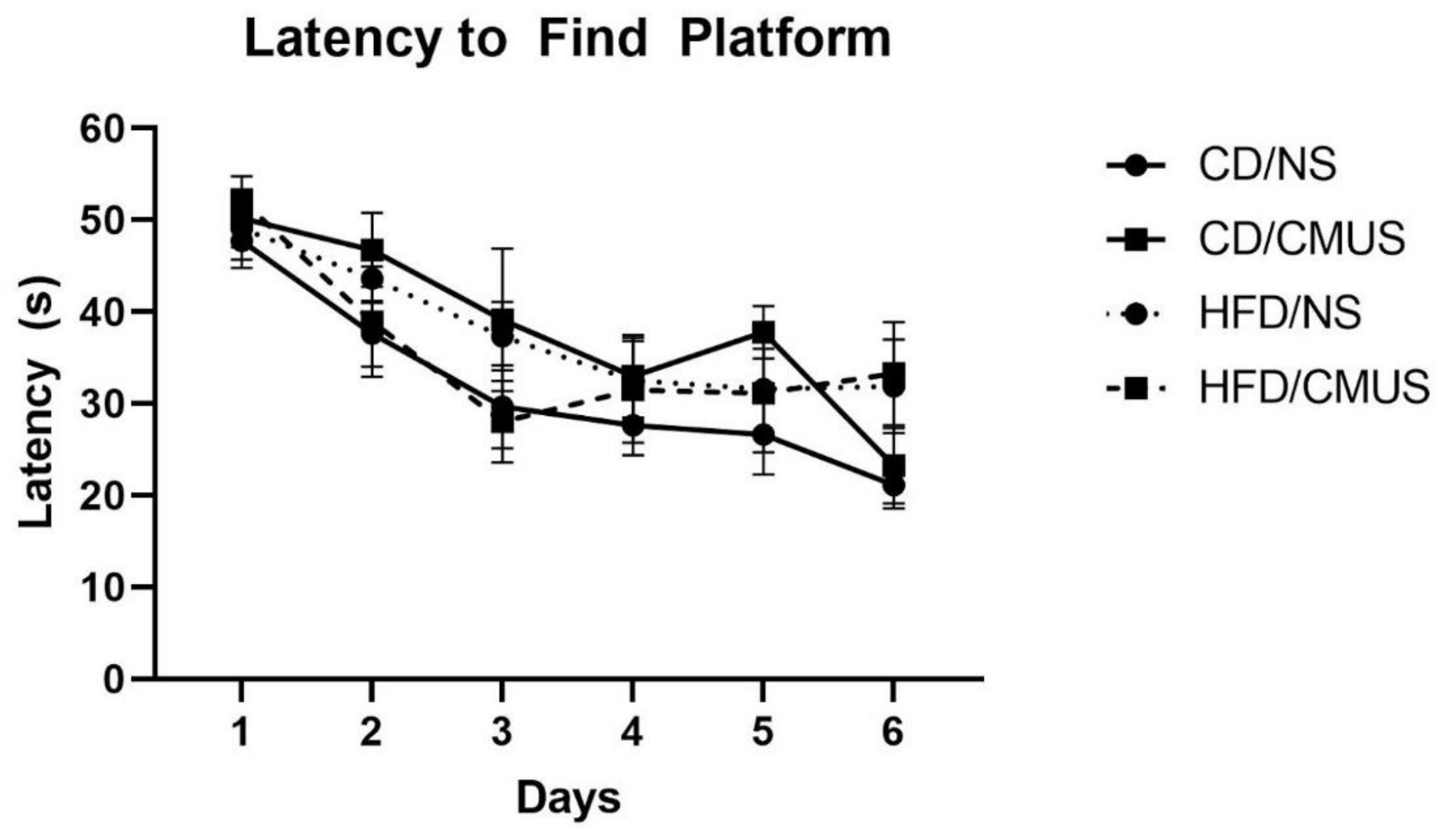
3.4.1. Latency to Find Platform
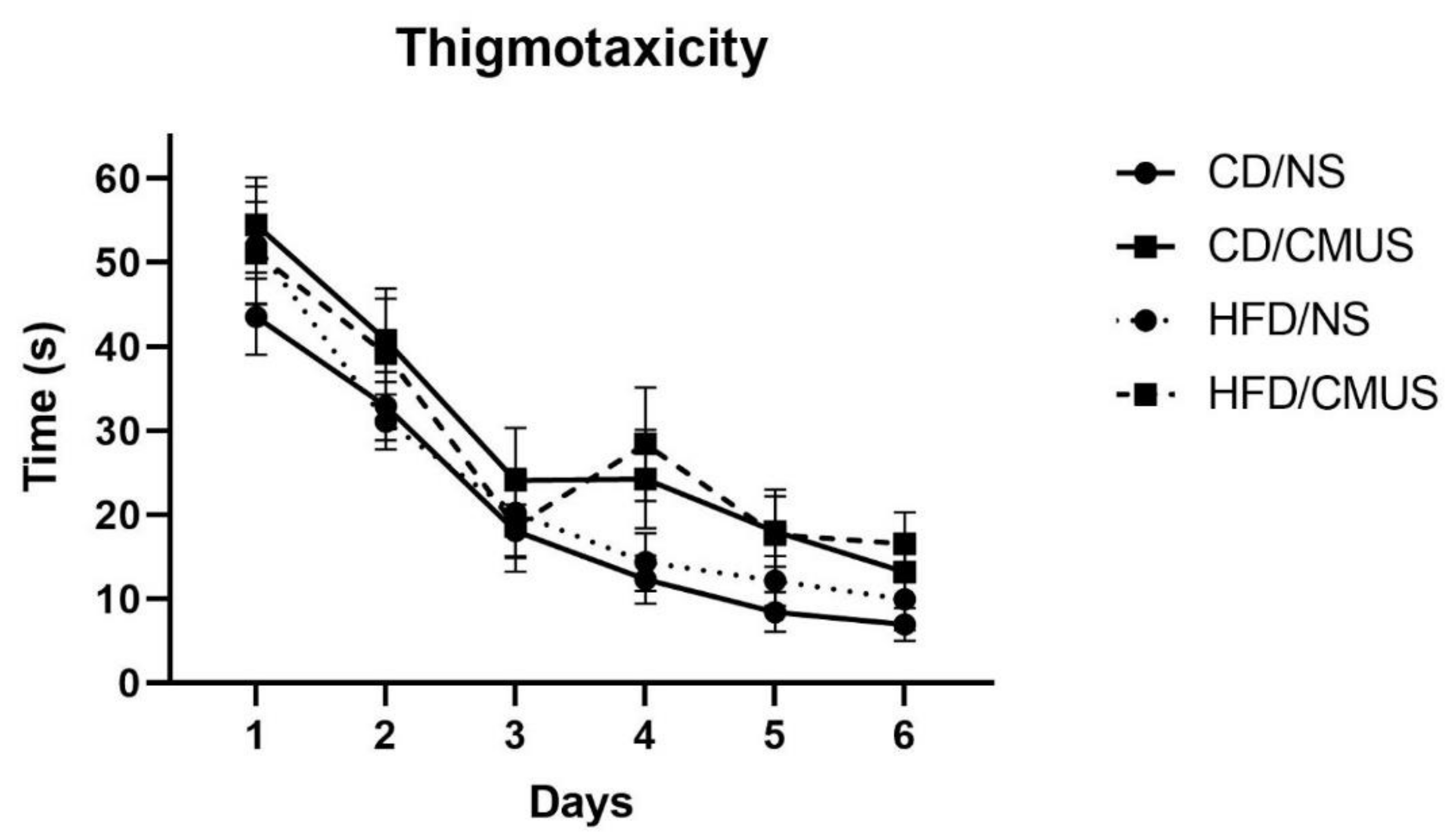
3.4.2. Thigmotaxicity
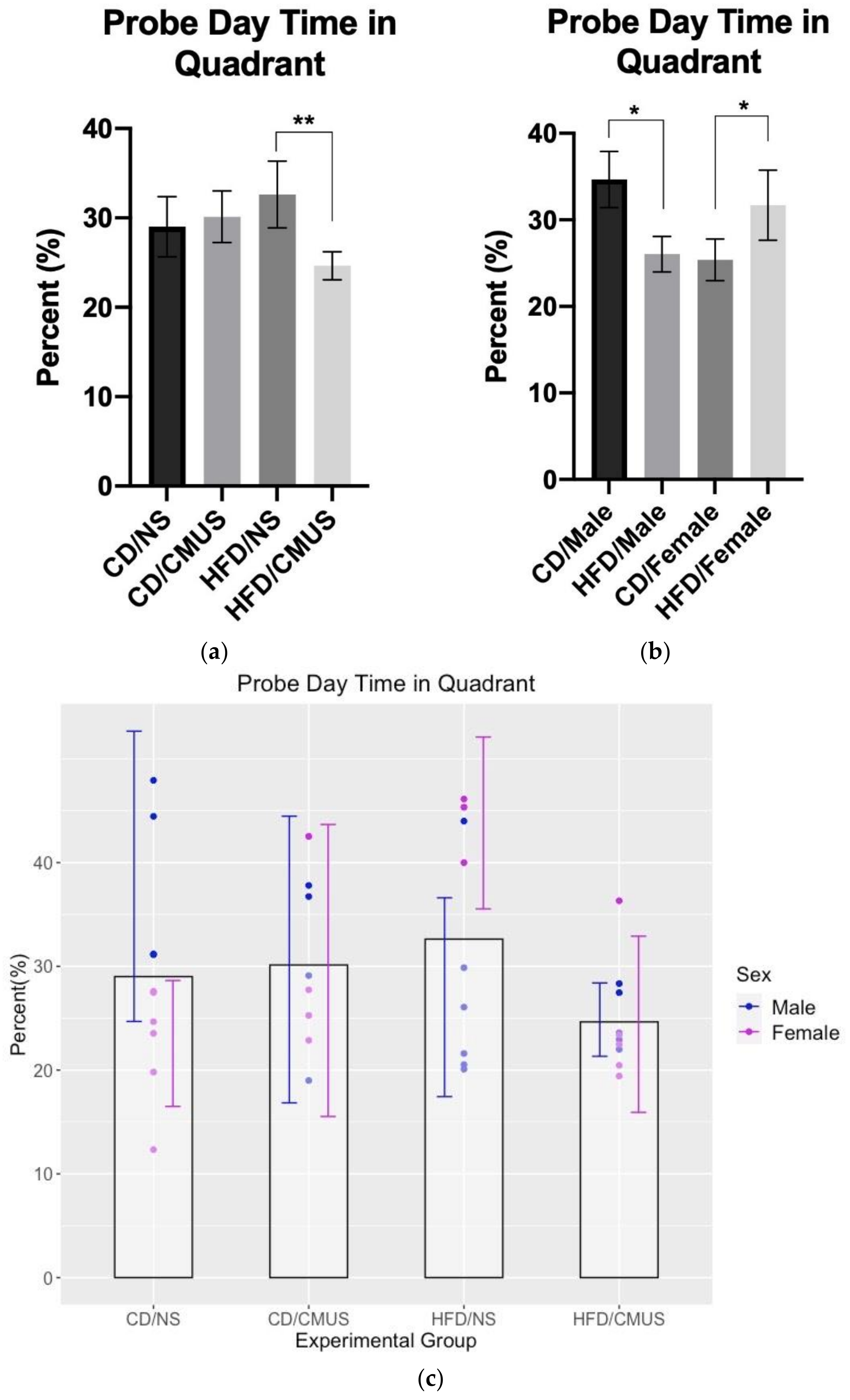
3.4.3. Day 7 Probe Trial (Percent Time Spent in the Target Quadrant, Day 7)
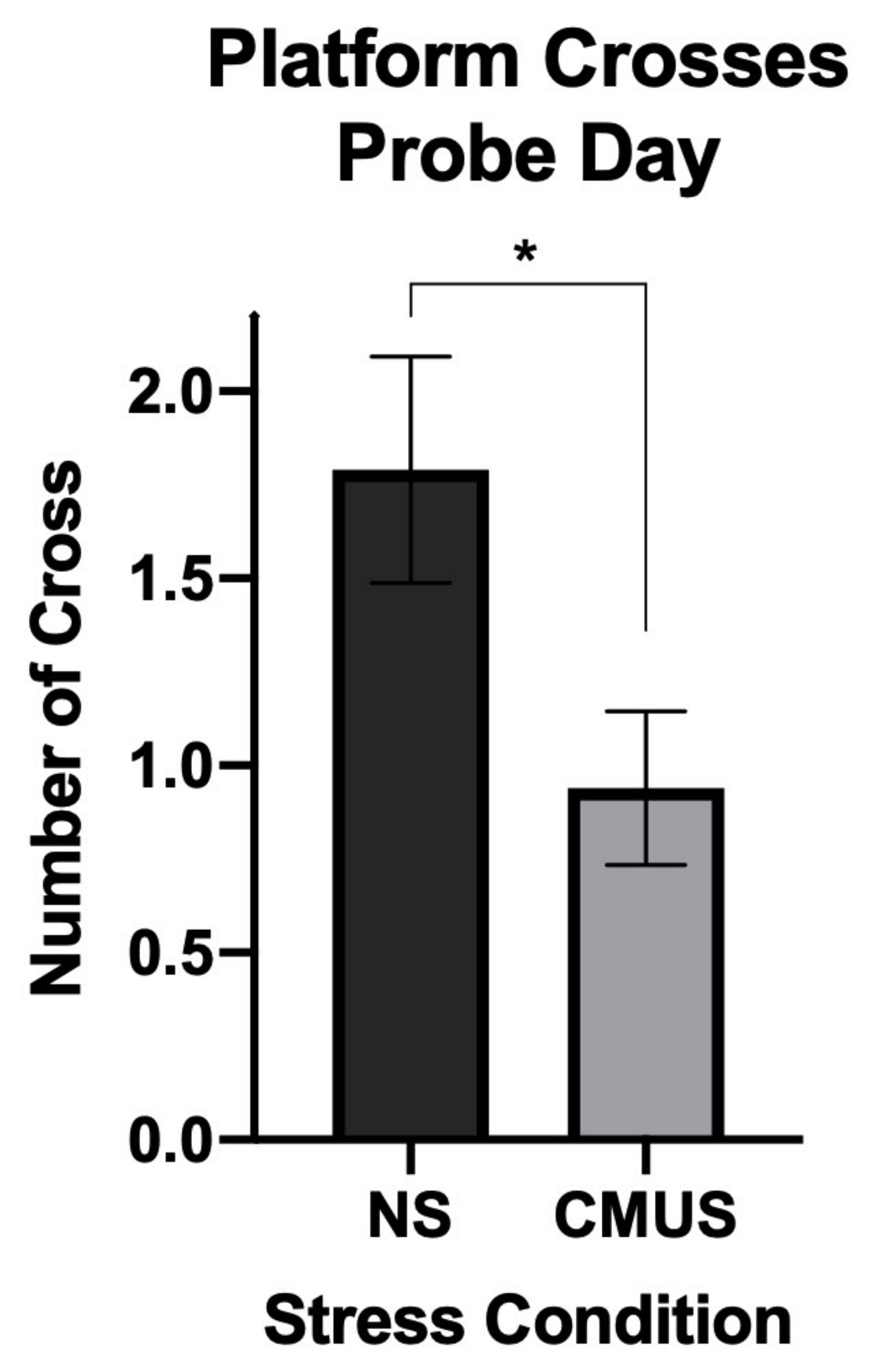
3.4.4. Day 7 Crosses
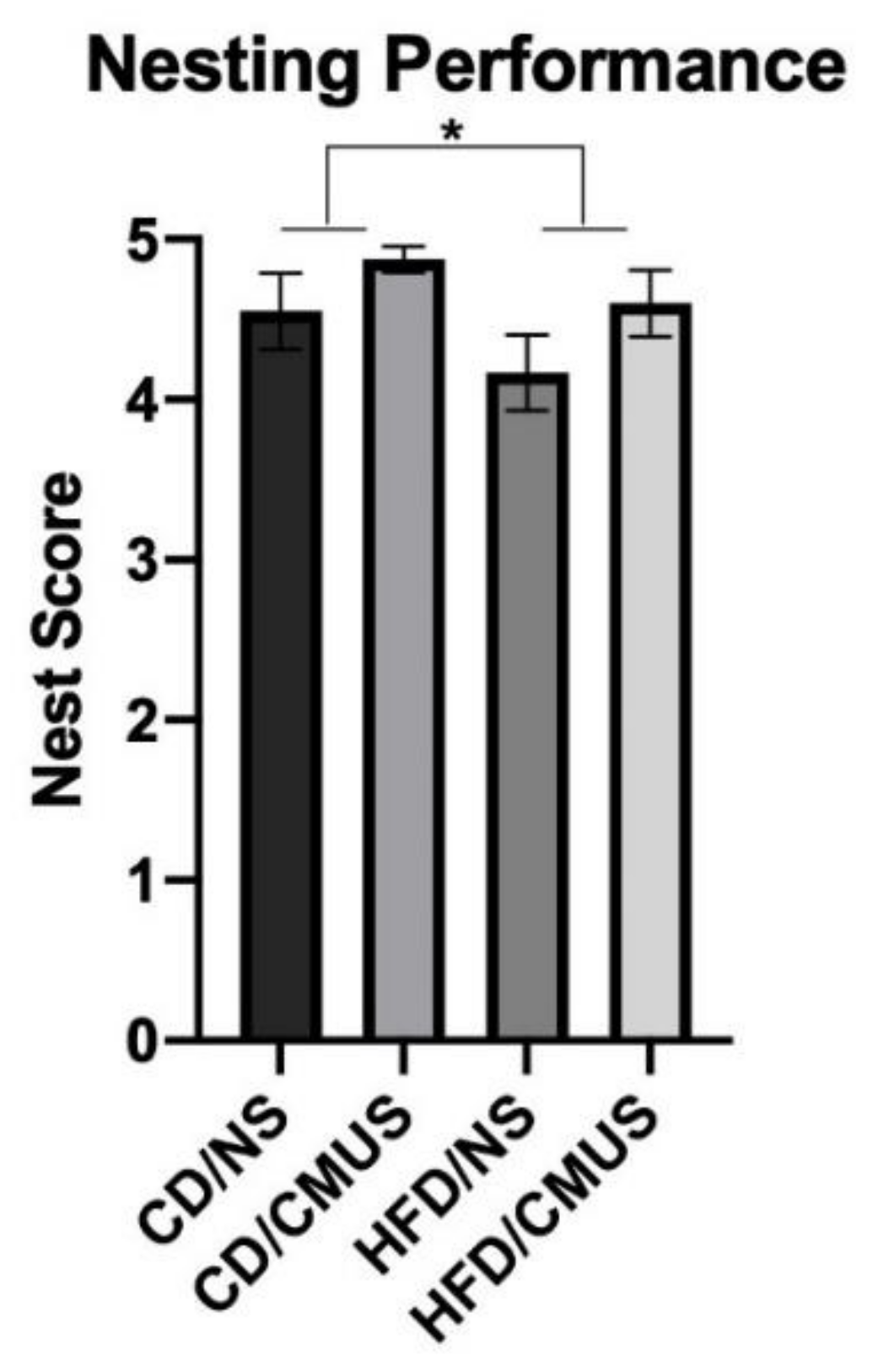
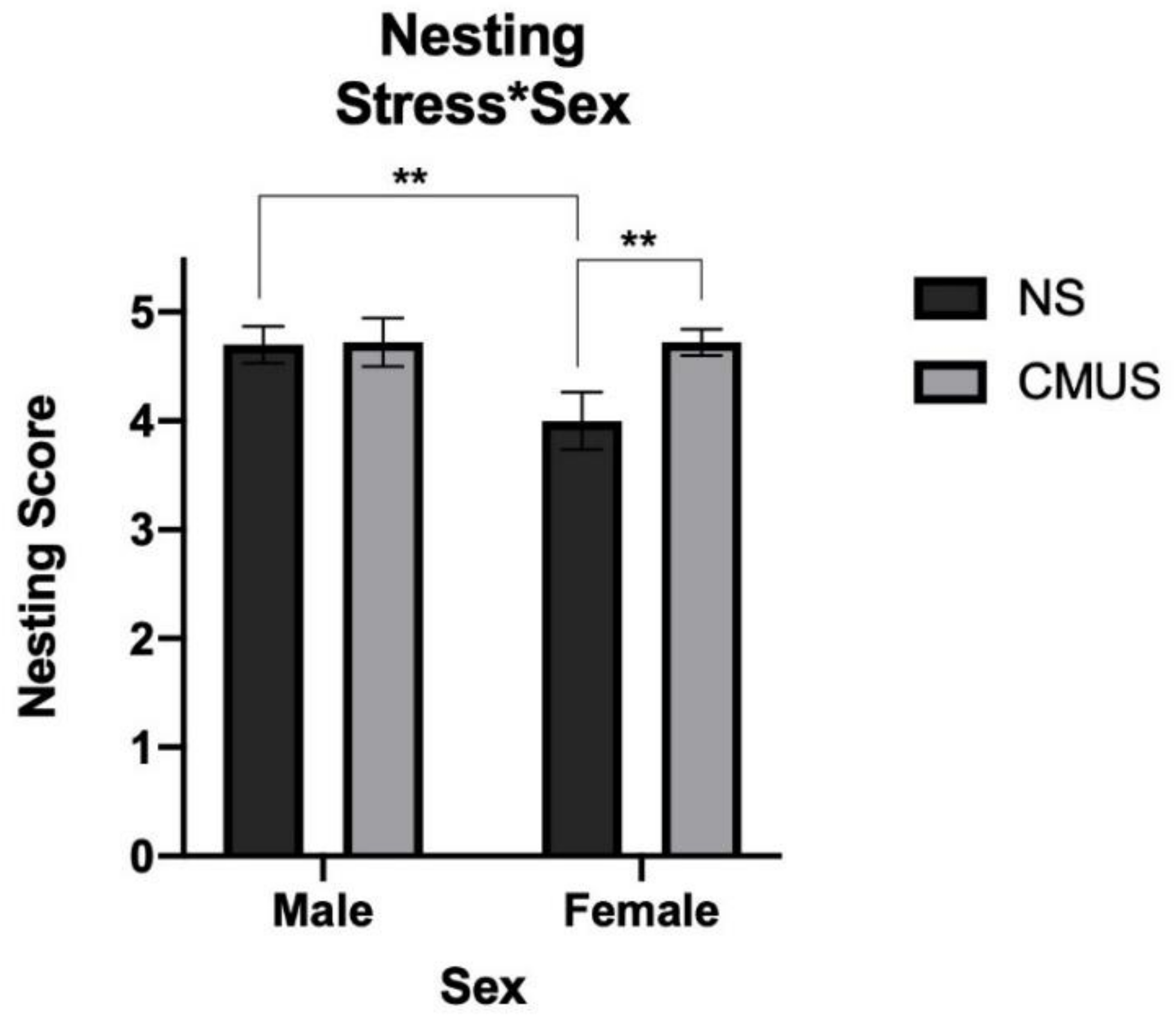
3.5. Nesting
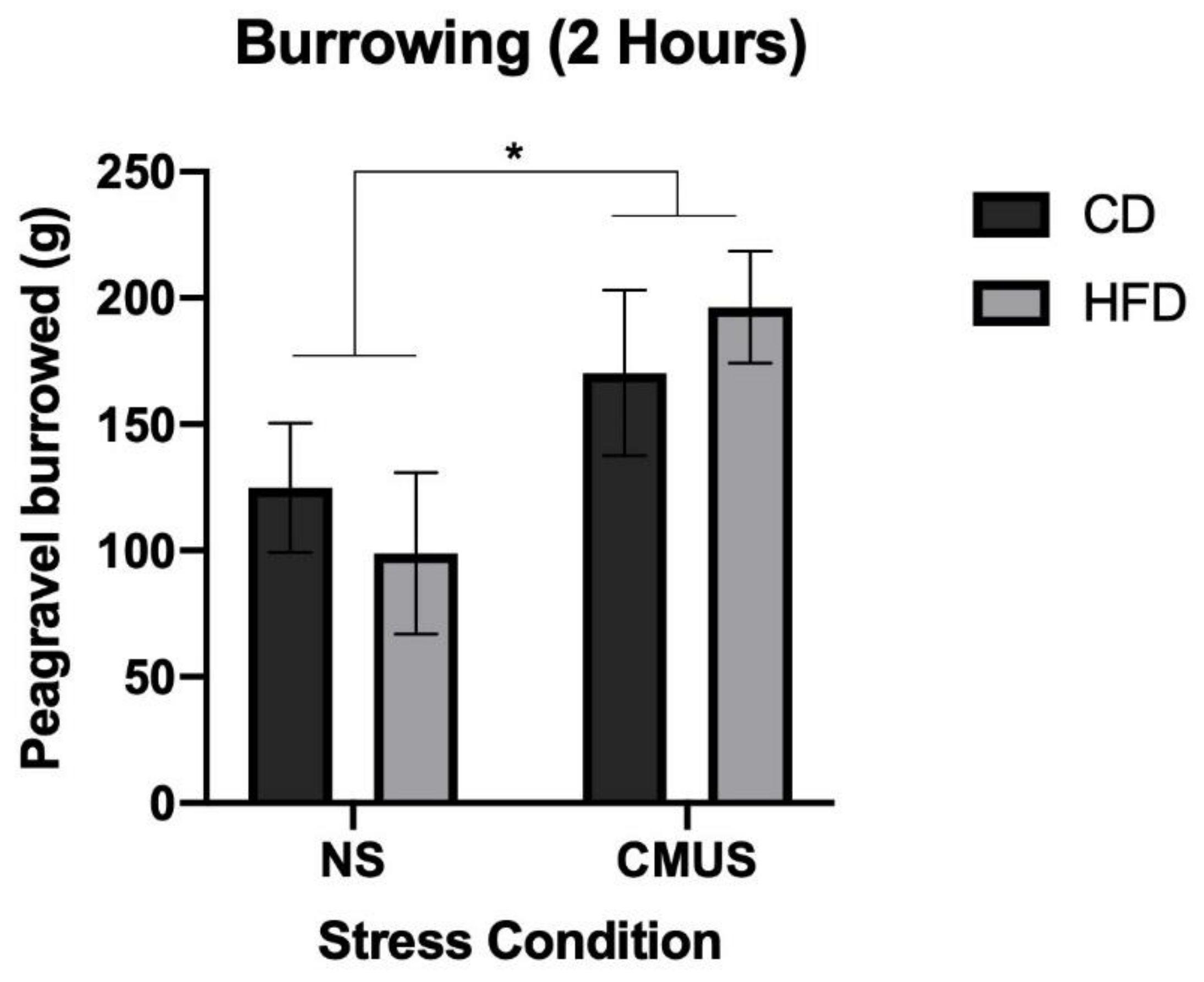
3.6. Burrowing
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yurgelun-Todd, D. Emotional and cognitive changes during adolescence. Curr. Opin. Neurobiol. 2007, 17, 251–257. [Google Scholar] [CrossRef]
- McCormick, C.M.; Smith, C.; Mathews, I.Z. Effects of chronic social stress in adolescence on anxiety and neuroendocrine response to mild stress in male and female rats. Behav. Brain Res. 2008, 187, 228–238. [Google Scholar] [CrossRef]
- Casey, B.; Jones, R.M.; Levita, L.; Libby, V.; Pattwell, S.S.; Ruberry, E.J.; Soliman, F.; Somerville, L.H. The storm and stress of adolescence: Insights from human imaging and mouse genetics. Dev. Psychobiol. 2010, 52, 225–235. [Google Scholar] [CrossRef] [Green Version]
- Bourke, C.H.; Neigh, G.N. Behavioral effects of chronic adolescent stress are sustained and sexually dimorphic. Horm. Behav. 2011, 60, 112–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaby, L.; Cavigelli, S.; Hirrlinger, A.; Caruso, M.; Braithwaite, V. Chronic unpredictable stress during adolescence causes long-term anxiety. Behav. Brain Res. 2015, 278, 492–495. [Google Scholar] [CrossRef] [PubMed]
- Meredith, C.N.; Dwyer, J.T. Nutrition and exercise: Effects on adolescent health. Annu. Rev. Public Health 1991, 12, 309–333. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-M.; Park, J.-M.; Song, M.K.; Kim, Y.J.; Kim, Y.-J. Comparison of the behavioral effects of exercise and high fat diet on cognitive function in adolescent rats. J. Exerc. Rehabil. 2016, 12, 520–525. [Google Scholar] [CrossRef] [Green Version]
- Boitard, C.; Etchamendy, N.; Sauvant, J.; Aubert, A.; Tronel, S.; Marighetto, A.; Layé, S.; Ferreira, G. Juvenile, but not adult exposure to high-fat diet impairs relational memory and hippocampal neurogenesis in mice. Hippocampus 2012, 22, 2095–2100. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Morales, L.; Ruiz-Gayo, M.; del Olmo, N. Effect of high-fat diets on mood and learning performance in adolescent mice. Behav. Brain Res. 2016, 311, 167–172. [Google Scholar] [CrossRef]
- Kelly, A.S.; Barlow, S.E.; Rao, G.; Inge, T.H.; Hayman, L.L.; Steinberger, J.; Urbina, E.M.; Ewing, L.J.; Daniels, S.R. Severe obesity in children and adolescents: Identification, associated health risks, and treatment approaches. Circulation 2013, 128, 1689–1712. [Google Scholar] [CrossRef]
- Gómez-Pinilla, F. Brain foods: The effects of nutrients on brain function. Nat. Rev. Neurosci. 2008, 9, 568–578. [Google Scholar] [CrossRef] [Green Version]
- Turner, J. Your brain on food: A nutrient-rich diet can protect cognitive health. J. Am. Soc. Aging 2011, 35, 99–106. [Google Scholar]
- Kanoski, S.E.; Davidson, T.L. Western diet consumption and cognitive impairment: Links to hippocampal dysfunction and obesity. Physiol. Behav. 2011, 103, 59–68. [Google Scholar] [CrossRef] [Green Version]
- CDC—National Center for Chronic Disease Prevention and Health Promotion (NCCDPHP). Available online: www.cdc.gov/chronicdisease/pdf/factsheets/poor-nutrition-H.pdf (accessed on 12 November 2020).
- O’Dea, J.A. Why do kids eat healthful food? Perceived benefits of and barriers to healthful eating and physical activity among children and adolescents. J. Am. Diet. Assoc. 2003, 103, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Bibiloni, M.D.M.; Pons, A.; Tur, J.A. Prevalence of overweight and obesity in adolescents: A systematic review. Int. Sch. Res. Not. 2013, 2013, 392747. [Google Scholar] [CrossRef] [PubMed]
- Walther, J.; Aldrian, U.; Stüger, H.P.; Kiefer, I.; Ekmekcioglu, C. Nutrition, lifestyle factors, and mental health in adolescents and young adults living in Austria. Int. J. Adolesc. Med. Health 2014, 26, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Heyward, F.D.; Gilliam, D.; Coleman, M.A.; Gavin, C.F.; Wang, J.; Kaas, G.; Trieu, R.; Lewis, J.; Moulden, J.; Sweatt, J.D. Obesity weighs down memory through a mechanism involving the neuroepigenetic dysregulation of sirt1. J. Neurosci. 2016, 36, 1324–1335. [Google Scholar] [CrossRef] [Green Version]
- Langdon, K.; Clarke, J.; Corbett, D. Long-term exposure to high fat diet is bad for your brain: Exacerbation of focal ischemic brain injury. Neuroscience 2011, 182, 82–87. [Google Scholar] [CrossRef]
- Francis, H.; Stevenson, R. The longer-term impacts of Western diet on human cognition and the brain. Appetite 2013, 63, 119–128. [Google Scholar] [CrossRef]
- Stampfer, M.J.; Hu, F.B.; Manson, J.E.; Rimm, E.B.; Willett, W.C. Primary prevention of coronary heart disease in women through diet and lifestyle. N. Engl. J. Med. 2000, 343, 16–22. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Chen, P.-C.; Kuo, H.-K.; Lin, L.-Y.; Lin, J.-W.; Hwang, J.-J. Effects of obesity, physical activity, and cardiorespiratory fitness on blood pressure, inflammation, and insulin resistance in the National Health and Nutrition Survey 1999–2002. Nutr. Metab. Cardiovasc. Dis. 2010, 20, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Kothari, V.; Luo, Y.; Tornabene, T.; O’Neill, A.M.; Greene, M.W.; Geetha, T.; Babu, J.R. High fat diet induces brain insulin resistance and cognitive impairment in mice. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2017, 1863, 499–508. [Google Scholar] [CrossRef]
- Buwalda, B.; Geerdink, M.; Vidal, J.; Koolhaas, J.M. Social behavior and social stress in adolescence: A focus on animal models. Neurosci. Biobehav. Rev. 2011, 35, 1713–1721. [Google Scholar] [CrossRef]
- Lupien, S.J.; McEwen, B.S.; Gunnar, M.R.; Heim, C. Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat. Rev. Neurosci. 2009, 10, 434–445. [Google Scholar] [CrossRef]
- Koch, F.-S.; Sepa, A.; Ludvigsson, J. Psychological stress and obesity. J. Pediatr. 2008, 153, 839–844. [Google Scholar] [CrossRef] [Green Version]
- De Vriendt, T.; Moreno, L.; de Henauw, S. Chronic stress and obesity in adolescents: Scientific evidence and methodological issues for epidemiological research. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Dallman, M.F. Stress-induced obesity and the emotional nervous system. Trends Endocrinol. Metab. 2010, 21, 159–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salleh, M.R. Life Event, Stress and Illness. Malaysian J. Med. Sci. 2008, 15, 9–18. [Google Scholar]
- Yudkin, J.S.; Kumari, M.; E Humphries, S.; Mohamed-Ali, V. Inflammation, obesity, stress and coronary heart disease: Is interleukin-6 the link? Atherosclerosis 2000, 148, 209–214. [Google Scholar] [CrossRef]
- Lupien, S.J.; Lepage, M. Stress, memory, and the hippocampus: Can’t live with it, can’t live without it. Behav. Brain Res. 2001, 127, 137–158. [Google Scholar] [CrossRef]
- Bruder-Nascimento, T.; Campos, D.H.S.; Alves, C.; Thomaz, S.; Cicogna, A.C.; Cordellini, S. Effects of chronic stress and high-fat diet on metabolic and nutritional parameters in Wistar rats. Arq. Bras. Endocrinol. Metabol. 2013, 57, 642–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteiro, S.; Roque, S.; de Sã¡-Calã§ada, D.; Sousa, N.; Correia-Neves, M.; Cerqueira, J.J.; de Sá-Calçada, D. An efficient chronic unpredictable stress protocol to induce stress-related responses in C57BL/6 mice. Front. Psychiatry 2015, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Han, B.; Yu, L.; Geng, Y.; Shen, L.; Wang, H.; Wang, Y.; Wang, J.; Wang, M. Chronic stress aggravates cognitive impairment and suppresses insulin associated signaling pathway in APP/PS1 mice. J. Alzheimer Dis. 2016, 53, 1539–1552. [Google Scholar] [CrossRef] [PubMed]
- DeVallance, E.; Riggs, D.; Jackson, B.; Parkulo, T.; Zaslau, S.; Chantler, P.D.; Olfert, I.M.; Bryner, R.W. Effect of chronic stress on running wheel activity in mice. PLoS ONE 2017, 12, e0184829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridgewater, L.C.; Zhang, C.; Wu, Y.; Hu, W.; Zhang, Q.; Wang, J.; Li, S.; Zhao, L. Gender-based differences in host behavior and gut microbiota composition in response to high fat diet and stress in a mouse model. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Cotella, E.M.; Gomez, A.S.; Lemen, P.; Chen, C.; Fernández, G.; Hansen, C.; Herman, J.P.; Paglini, M.G. Long-term impact of chronic variable stress in adolescence versus adulthood. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2019, 88, 303–310. [Google Scholar] [CrossRef]
- Jianhua, F.; Wei, W.; Xiaomei, L.; Shao-Hui, W. Chronic social defeat stress leads to changes of behaviour and memory-associated proteins of young mice. Behav. Brain Res. 2017, 316, 136–144. [Google Scholar] [CrossRef]
- Naninck, E.F.; Hoeijmakers, L.; Kakava-Georgiadou, N.; Meesters, A.; Lazic, S.E.; Lucassen, P.J.; Korosi, A. Chronic early life stress alters developmental and adult neurogenesis and impairs cognitive function in mice. Hippocampus 2014, 25, 309–328. [Google Scholar] [CrossRef]
- Cartwright, M.; Wardle, J.; Steggles, N.; Simon, A.E.; Croker, H.; Jarvis, M.J. Stress and dietary practices in adolescents. Health Psychol. 2003, 22, 362–369. [Google Scholar] [CrossRef]
- Kuo, L.E.; Czarnecka, M.; Kitlinska, J.B.; Tilan, J.U.; Kvetnansky, R.; Zukowska, Z. Chronic Stress, combined with a high-fat/high-sugar diet, shifts sympathetic signaling toward Neuropeptide Y and leads to obesity and the metabolic syndrome. Ann. N. Y. Acad. Sci. 2008, 1148, 232–237. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.L.; Liu, D.X.; Jiang, H.; Pan, F.; Ho, C.S.; Ho, R.C. The effects of high-fat-diet combined with chronic unpredictable mild stress on depression-like behavior and Leptin/LepRb in male rats. Sci. Rep. 2016, 6, 35239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Xiaoling, G.; Pingting, L.; Shuqiang, L.; Yuaner, Z. Chronic unpredictable mild stress combined with a high-fat diets aggravates atherosclerosis in rats. Lipids Health Dis. 2014, 13, 77. [Google Scholar] [CrossRef] [Green Version]
- Cudnoch-Jedrzejewska, A.; Gomolka, R.; Szczepanska-Sadowska, E.; Czarzasta, K.; Wrzesien, R.; Koperski, L.; Puchalska, L.; Wsol, A. High-fat diet and chronic stress reduce central pressor and tachycardic effects of apelin in Sprague-Dawley rats. Clin. Exp. Pharmacol. Physiol. 2015, 42, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Hwang, L.-L.; Wang, C.-H.; Li, T.-L.; Chang, S.-D.; Lin, L.-C.; Chen, C.-P.; Chen, C.-T.; Liang, K.-C.; Ho, I.-K.; Yang, W.-S.; et al. Sex differences in high-fat diet-induced obesity, metabolic alterations and learning, and synaptic plasticity deficits in mice. Obesity 2010, 18, 463–469. [Google Scholar] [CrossRef]
- Braun, A.A.; Skelton, M.R.; Vorhees, C.V.; Williams, M.T. Comparison of the elevated plus and elevated zero mazes in treated and untreated male Sprague—Dawley rats: Effects of anxiolytic and anxiogenic agents. Pharmacol. Biochem. Behav. 2011, 97, 406–415. [Google Scholar] [CrossRef] [Green Version]
- Morris, R.G.M. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Deacon, R.M.J. Burrowing in rodents: A sensitive method for detecting behavioral dysfunction. Nat. Protoc. 2006, 1, 118–121. [Google Scholar] [CrossRef] [PubMed]
- Deacon, R. Assessing burrowing, nest construction, and hoarding in mice. J. Vis. Exp. 2012, 59, e2607. [Google Scholar] [CrossRef] [Green Version]
- Neely, C.L.; Pedemonte, K.A.; Boggs, K.N.; Flinn, J.M. Nest building behavior as an early indicator of behavioral deficits in mice. J. Vis. Exp. 2019, 152, e60139. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 12 November 2020).
- Pothion, S.; Bizot, J.-C.; Trovero, F.; Belzung, C. Strain differences in sucrose preference and in the consequences of unpredictable chronic mild stress. Behav. Brain Res. 2004, 155, 135–146. [Google Scholar] [CrossRef]
- Schweizer, M.C.; Henniger, M.S.H.; Sillaber, I. Chronic Mild Stress (CMS) in mice: Of Anhedonia, ‘Anomalous Anxiolysis’ and activity. PLoS ONE 2009, 4, e4326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.Y.; Lee, D.H.; Kang, S.S. Effects of chronic restraint stress on body weight, food intake, and Hypothalamic Gene expressions in mice. Endocrinol. Metab. 2013, 28, 288–296. [Google Scholar] [CrossRef]
- Gil-Bea, F.J.; Aisa, B.; Schliebs, R.; Ramirez, M.J. Increase of locomotor activity underlying the behavioral disinhibition in Tg2576 mice. Behav. Neurosci. 2007, 121, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Lippi, S.L.P.; Smith, M.L.; Flinn, J.M. A novel hAPP/htau mouse model of Alzheimer’s disease: Inclusion of APP with Tau Exacerbates behavioral deficits and zinc administration heightens tangle pathology. Front. Aging Neurosci. 2018, 10, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivanathan, S.; Thavartnam, K.; Arif, S.; Elegino, T.; McGowan, P.O. Chronic high fat feeding increases anxiety-like behaviour and reduces transcript abundance of glucocorticoid signalling genes in the hippocampus of female rats. Behav. Brain Res. 2015, 286, 265–270. [Google Scholar] [CrossRef]
- Sah, S.K.; Lee, C.; Jang, J.-H.; Park, G.H. Effect of high-fat diet on cognitive impairment in triple-transgenic mice model of Alzheimer’s disease. Biochem. Biophys. Res. Commun. 2017, 493, 731–736. [Google Scholar] [CrossRef]
- Molteni, R.; Barnard, R.; Ying, Z.; Roberts, C.; Gómez-Pinilla, F. A high-fat, refined sugar diet reduces hippocampal brain-derived neurotrophic factor, neuronal plasticity, and learning. Neuroscience 2002, 112, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Knight, E.M.; Martins, I.V.; Gümüsgöz, S.; Allan, S.M.; Lawrence, C.B. High-fat diet-induced memory impairment in triple-transgenic Alzheimer’s disease (3xTgAD) mice is independent of changes in amyloid and tau pathology. Neurobiol. Aging 2014, 35, 1821–1832. [Google Scholar] [CrossRef]
- Zuloaga, K.L.; Johnson, L.A.; Roese, N.E.; Marzulla, T.; Zhang, W.; Nie, X.; Alkayed, F.N.; Hong, C.; Grafe, M.R.; Pike, M.M.; et al. High fat diet-induced diabetes in mice exacerbates cognitive deficit due to chronic hypoperfusion. J. Cereb. Blood Flow Metab. 2016, 36, 1257–1270. [Google Scholar] [CrossRef]
- Denver, P.; Gault, V.A.; McClean, P.L. Sustained high-fat diet modulates inflammation, insulin signalling and cognition in mice and a modified xenin peptide ameliorates neuropathology in a chronic high-fat model. Diabetes Obes. Metab. 2018, 20, 1166–1175. [Google Scholar] [CrossRef]
- Srivareerat, M.; Tran, T.T.; Alzoubi, K.H.; Alkadhi, K.A. Chronic psychosocial stress exacerbates impairment of cognition and long-term potentiation in β-Amyloid rat model of Alzheimer’s disease. Biol. Psychiatry 2009, 65, 918–926. [Google Scholar] [CrossRef]
- Valladolid-Acebes, I.; Fole, A.; Martín, M.; Morales, L.; Cano, M.V.; Ruiz-Gayo, M.; del Olmo, N. Spatial memory impairment and changes in hippocampal morphology are triggered by high-fat diets in adolescent mice. Is there a role of leptin? Neurobiol. Learn. Mem. 2013, 106, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Hei, M.; Chen, P.; Wang, S.; Li, X.; Xu, M.; Zhu, X.; Wang, Y.; Duan, J.; Huang, Y.; Zhao, S. Effects of chronic mild stress induced depression on synaptic plasticity in mouse hippocampus. Behav. Brain Res. 2019, 365, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.M.; Marriott, D.; Trotter, J.; Hammond, T.; Lyman, D.; Call, T.; Walker, B.; Christensen, N.; Haynie, D.; Badura, Z.; et al. Running exercise mitigates the negative consequences of chronic stress on dorsal hippocampal long-term potentiation in male mice. Neurobiol. Learn. Mem. 2018, 149, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Huang, W.; Haile, C.N.; Kosten, T.A. Hippocampal GluR1 associates with behavior in the elevated plus maze and shows sex differences. Behav. Brain Res. 2011, 222, 326–331. [Google Scholar] [CrossRef] [Green Version]
- Bustamante, C.; Bilbao, P.; Contreras, W.; Martinez, M.; Mendoza, A.; Reyes, Á.; Pascual, R. Effects of prenatal stress and exercise on dentate granule cells maturation and spatial memory in adolescent mice. Int. J. Dev. Neurosci. 2010, 28, 605–609. [Google Scholar] [CrossRef]
- Jung, S.H.; Brownlow, M.L.; Pellegrini, M.; Jankord, R. Divergence in Morris water maze-based cognitive performance under chronic stress is associated with the hippocampal whole transcriptomic modification in mice. Front. Mol. Neurosci. 2017, 10, 275. [Google Scholar] [CrossRef] [Green Version]
- Conrad, C.D. A critical review of chronic stress effects on spatial learning and memory. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2010, 34, 742–755. [Google Scholar] [CrossRef]
- Huang, R.-R.; Hu, W.; Yin, Y.-Y.; Wang, Y.-C.; Li, W.-P. Chronic restraint stress promotes learning and memory impairment due to enhanced neuronal endoplasmic reticulum stress in the frontal cortex and hippocampus in male mice. Int. J. Mol. Med. 2014, 35, 553–559. [Google Scholar] [CrossRef]
- Boitard, C.; Cavaroc, A.; Sauvant, J.; Aubert, A.; Castanon, N.; Layé, S.; Ferreira, G. Impairment of hippocampal-dependent memory induced by juvenile high-fat diet intake is associated with enhanced hippocampal inflammation in rats. Brain Behav. Immun. 2014, 40, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Vinuesa, A.; Pomilio, C.; Menafra, M.; Bonaventura, M.M.; Garay, L.; Mercogliano, M.F.; Schillaci, R.; Lantos, R.S.V.L.; Brites, F.; Beauquis, J.; et al. Juvenile exposure to a high fat diet promotes behavioral and limbic alterations in the absence of obesity. Psychoneuroendocrinology 2016, 72, 22–33. [Google Scholar] [CrossRef]
- Kim, D.-M.; Leem, Y.-H. Chronic stress-induced memory deficits are reversed by regular exercise via AMPK-mediated BDNF induction. Neuroscience 2016, 324, 271–285. [Google Scholar] [CrossRef]
- Jirkof, P. Burrowing and nest building behavior as indicators of well-being in mice. J. Neurosci. Methods 2014, 234, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Torres-Lista, V.; Giménez-Llort, L. Impairment of nesting behaviour in 3xTg-AD mice. Behav. Brain Res. 2013, 247, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Filali, M.; LaLonde, R.; Rivest, S. Subchronic memantine administration on spatial learning, exploratory activity, and nest-building in an APP/PS1 mouse model of Alzheimer’s disease. Neuropharmacology 2011, 60, 930–936. [Google Scholar] [CrossRef]
- Craven, K.M.; Kochen, W.R.; Hernandez, C.M.; Flinn, J.M. Zinc Exacerbates Tau Pathology in a Tau Mouse model. J. Alzheimer Dis. 2018, 64, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Otabi, H.; Goto, T.; Okayama, T.; Kohari, D.; Toyoda, A. Subchronic and mild social defeat stress alter mouse nest building behavior. Behav. Process. 2016, 122, 21–25. [Google Scholar] [CrossRef]
- Otabi, H.; Okayama, T.; Toyoda, A. Assessment of nest building and social interaction behavior in mice exposed to acute social defeat stress using a three-dimensional depth camera. Anim. Sci. J. 2020, 91, 13447. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.-D.; Xiong, W.-D.; Xiong, S.-S.; Chen, G.-H. Age- and gender-based differences in nest-building behavior and learning and memory performance measured using a radial six-armed water maze in C57BL/6 mice. Behav. Neurol. 2018, 2018, 8728415. [Google Scholar] [CrossRef] [Green Version]
- Estep, D.Q.; Lanier, D.L.; Dewsbury, D.A. Copulatory behavior and nest building behavior of wild house mice (Mus musculus). Learn. Behav. 1975, 3, 329–336. [Google Scholar] [CrossRef] [Green Version]
- Tamashiro, K.L.; Terrillion, C.E.; Hyun, J.; Koenig, J.I.; Moran, T.H. Prenatal stress or high-fat diet increases susceptibility to diet-induced Obesity in rat offspring. Diabetes 2009, 58, 1116–1125. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, A.; de Vega, W.; Sivanathan, S.; St-Cyr, S.; McGowan, P. Maternal high-fat diet alters anxiety behavior and glucocorticoid signaling in adolescent offspring. Neuroscience 2014, 272, 92–101. [Google Scholar] [CrossRef]
- Pini, R.T.B.; Vales, L.D.M.F.D.; Costa, T.M.B.; Almeida, S.S. Effects of cafeteria diet and high fat diet intake on anxiety, learning and memory in adult male rats. Nutr. Neurosci. 2016, 20, 396–408. [Google Scholar] [CrossRef]
- Ferreira, A.; Castro, J.P.; Andrade, J.P.; Madeira, M.D.; Cardoso, A. Cafeteria-diet effects on cognitive functions, anxiety, fear response and neurogenesis in the juvenile rat. Neurobiol. Learn. Mem. 2018, 155, 197–207. [Google Scholar] [CrossRef]
- Lewis, A.R.; Singh, S.; Youssef, F.F. Cafeteria-diet induced obesity results in impaired cognitive functioning in a rodent model. Heliyon 2019, 5, e01412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemos, C.; Rial, D.; Gonçalves, F.; Pires, J.; Silva, H.; Matheus, F.; da Silva, A.; Marques, J.; Rodrigues, R.; Jarak, I.; et al. High sucrose consumption induces memory impairment in rats associated with electrophysiological modifications but not with metabolic changes in the hippocampus. Neuroscience 2016, 315, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Beilharz, J.; Maniam, J.; Morris, M. Short-term exposure to a diet high in fat and sugar, or liquid sugar, selectively impairs hippocampal-dependent memory, with differential impacts on inflammation. Behav. Brain Res. 2016, 306, 1–7. [Google Scholar] [CrossRef]
- Vellers, H.L.; Letsinger, A.C.; Walker, N.R.; Granados, J.Z.; Lightfoot, J.T. High fat high sugar diet reduces voluntary wheel running in mice independent of sex hormone involvement. Front. Physiol. 2017, 8, 628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folha, O.A.D.A.C.; Bahia, C.P.; de Aguiar, G.P.S.; Herculano, A.M.; Coelho, N.L.G.; de Sousa, M.B.C.; Shiramizu, V.K.M.; Galvão, A.C.D.M.; de Carvalho, W.A.; Pereira, A. Effect of chronic stress during adolescence in prefrontal cortex structure and function. Behav. Brain Res. 2017, 326, 44–51. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Diet (CD) | High-Fat Diet (HFD) | ||
|---|---|---|---|
| No Stress | 10 | 9 | N = 19 |
| (NS) | (4M, 6F) | (6M, 3F) | |
| Chronic Mild Unpredictable Stress | 8 | 10 | N = 18 |
| (CMUS) | (4M, 4F) | (5M, 5F) | |
| N = 18 | N = 19 | N = 37 |
| Stressor | Procedure |
|---|---|
| Forced Swim—cold water swim (8–10 °C) for 5 min | Mice are placed into a container filled with cold water (8–10 °C) for 5 min. Water is changed between mice and temperature is checked prior to placing mice into the container. Mice are placed under a heat lamp after completion of the stressor. |
| Overnight water deprivation—during the dark phase (12 h) | Water bottles are removed from the cage before lights out and are replaced at lights on the following morning. |
| Bright light/open field exposure (10 min) | Mice are placed in an elevated open field box (not the same as the open field box used in the Open Field Test (see Section Open Field Test (OFT))) with 2 overhead lights shining down on them for 10 min. |
| Altered Light Cycle (during dark phase) | During the dark phase, animals in the stress groups are transported into the testing room and are kept in ‘lights on’ for the evening. At the conclusion of the dark phase, animals are placed back onto the caging racks in the vivarium. |
| Damp bedding (2 h) | Animal bedding is soaked with water and mice are left in the cage for 2 h. |
| No bedding (2 h) | Animal bedding is taken out and mice are placed into the empty cage for 2 h. |
| Overnight social isolation (12 h) | Animals are housed in individual shoebox cages (Ancare) with access to food and water overnight (12 h). |
| Tilted Cage (1 h) | Animal home cages are tilted for 1 h. |
| Predator Urine (1 h) | A small amount (~1 mL) of bobcat urine is placed on a cotton ball in a centrifuge tube in the mouse’s cage (mice have no direct contact with the urine). This is left in the cage for 1 h. After this stressor, mice are placed into a new home cage. |
| Bath (no bedding) (2 h) | Animal bedding is removed and a small amount of water is placed into the cage, covering the floor. Mice remain in this cage for 2 h. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lippi, S.L.P. Chronic Mild Unpredictable Stress and High-Fat Diet Given during Adolescence Impact Both Cognitive and Noncognitive Behaviors in Young Adult Mice. Brain Sci. 2021, 11, 260. https://doi.org/10.3390/brainsci11020260
Lippi SLP. Chronic Mild Unpredictable Stress and High-Fat Diet Given during Adolescence Impact Both Cognitive and Noncognitive Behaviors in Young Adult Mice. Brain Sciences. 2021; 11(2):260. https://doi.org/10.3390/brainsci11020260
Chicago/Turabian StyleLippi, Stephen L. P. 2021. "Chronic Mild Unpredictable Stress and High-Fat Diet Given during Adolescence Impact Both Cognitive and Noncognitive Behaviors in Young Adult Mice" Brain Sciences 11, no. 2: 260. https://doi.org/10.3390/brainsci11020260
APA StyleLippi, S. L. P. (2021). Chronic Mild Unpredictable Stress and High-Fat Diet Given during Adolescence Impact Both Cognitive and Noncognitive Behaviors in Young Adult Mice. Brain Sciences, 11(2), 260. https://doi.org/10.3390/brainsci11020260