
A Scientometric Approach to Review the Role of the Medial Preoptic Area (MPOA) in Parental Behavior

 , , and
, , and
Abstract
:1. Introduction
2. The Present Study
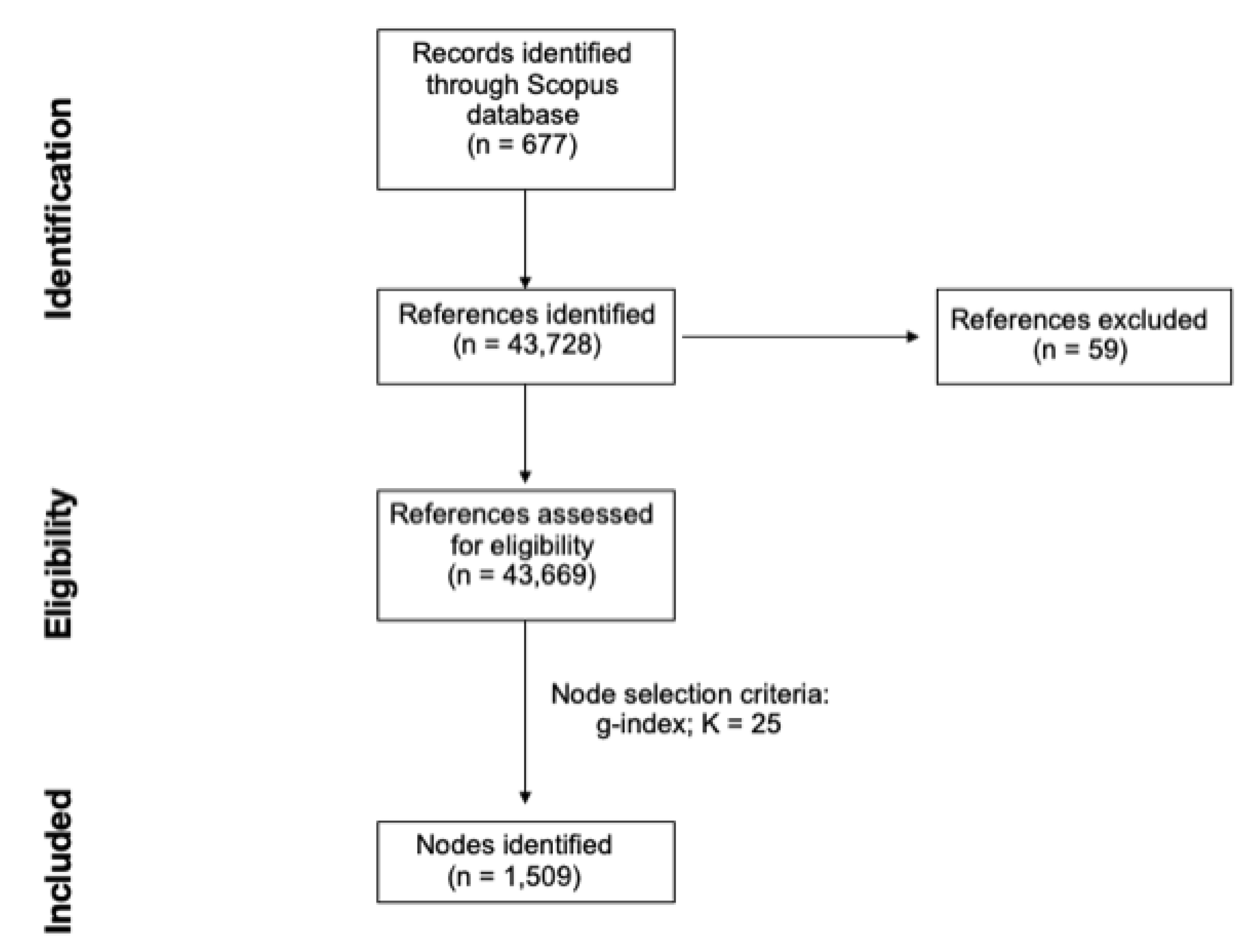
3. Materials and Methods
4. Results
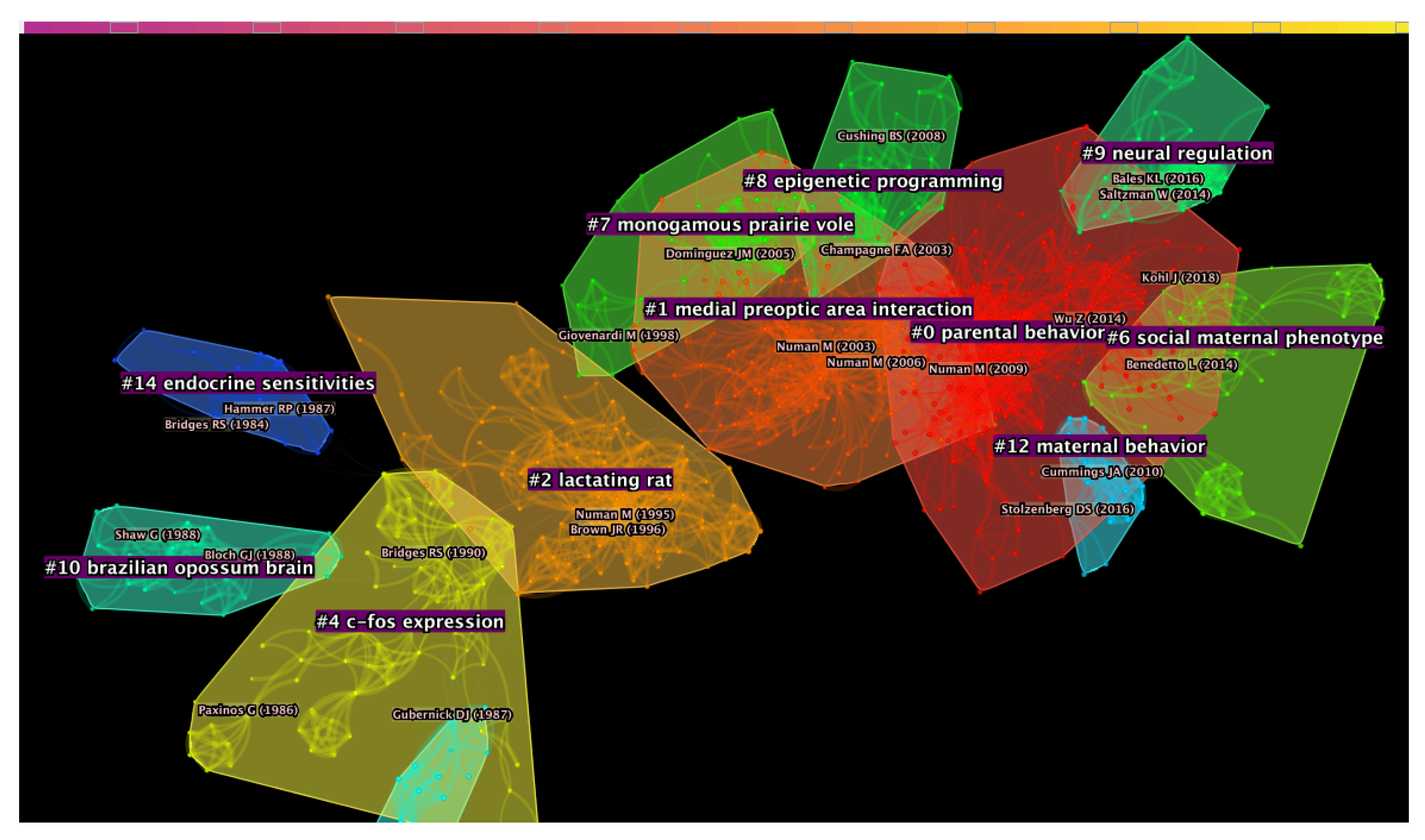
4.1. Document Co-Citation Analysis
4.2. Keywords Analysis
5. Discussion
5.1. Document Co-Citation Analysis
5.1.1. Cluster #4: “C-Fos Expression”
5.1.2. Cluster #2: “Lactating Rat”
5.1.3. Cluster #1: “Medial Preoptic Area Interaction”
5.1.4. Cluster #0: “Parental Behavior”
5.2. Keywords Analysis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| DCA | Document co-citation analysis |
| Fos-ir | Fos-immunoreactive |
| Fos-lir | Fos-like immunoreactivity |
| HPA | hypothalamic–pituitary–adrenal |
| MPOA | Medial preoptic area |
References
- Rosenblatt, J.; Lehrman, D. Maternal Behavior in Mammals; John Wiley & Sons, Inc.: New York, NY, USA, 1963. [Google Scholar]
- Lonstein, J.S.; Fleming, A.S. Parental behaviors in rats and mice. Curr. Protoc. Neurosci. 2001, 17, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Numan, M. Neurobiology of Social Behavior: Toward an Understanding of the Prosocial and Antisocial Brain; Academic Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Trivers, R.L. Parental investment and sexual selection. In Sexual Selection & the Descent of Man, Aldine de Gruyter, New York; Aldine-Atherton: Chicago, IL, USA, 1972; pp. 136–179. [Google Scholar]
- Elwood, R. Changes in the responses of male and female gerbils (Meriones unguiculatus) towards test pups during the pregnancy of the female. Anim. Behav. 1977, 25, 46–51. [Google Scholar] [CrossRef]
- Vom Saal, F.S. Time-contingent change in infanticide and parental behavior induced by ejaculation in male mice. Physiol. Behav. 1985, 34, 7–15. [Google Scholar] [CrossRef]
- Mennella, J.A.; Moltz, H. Infanticide in the male rat: The role of the vomeronasal organ. Physiol. Behav. 1988, 42, 303–306. [Google Scholar] [CrossRef]
- Jakubowski, M.; Terkel, J. Incidence of pup killing and parental behavior in virgin female and male rats (Rattus norvegicus): Differences between Wistar and Sprague-Dawley stocks. J. Comp. Psychol. 1985, 99, 93. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.E. Social and hormonal factors influencing infanticide and its suppression in adult male Long-Evans rats (Rattus norvegicus). J. Comp. Psychol. 1986, 100, 155. [Google Scholar] [CrossRef] [PubMed]
- Quadagno, D.M.; Rockwell, J. The effect of gonadal hormones in infancy on maternal behavior in the adult rat. Horm. Behav. 1972, 3, 55–62. [Google Scholar] [CrossRef]
- Fleischer, S.; Kordower, J.H.; Kaplan, B.; Dicker, R.; Smerling, R.; Ilgner, J. Olfactory bulbectomy and gender differences in maternal behaviors of rats. Physiol. Behav. 1981, 26, 957–959. [Google Scholar] [CrossRef]
- Samuels, M.H.; Bridges, R.S. Plasma prolactin concentrations in parental male and female rats: Effects of exposure to rat young. Endocrinology 1983, 113, 1647–1654. [Google Scholar] [CrossRef]
- Mayer, A.D.; Freeman, N.; Rosenblatt, J.S. Ontogeny of maternal behavior in the laboratory rat: Factors underlying changes in responsiveness from 30 to 90 days. Dev. Psychobiol. J. Int. Soc. Dev. Psychobiol. 1979, 12, 425–439. [Google Scholar] [CrossRef]
- Gray, P.; Chesley, S. Development of maternal behavior in nulliparous rats (Rattus norvegicus): Effects of sex and early maternal experience. J. Comp. Psychol. 1984, 98, 91. [Google Scholar] [CrossRef]
- Lee, A.; Clancy, S.; Fleming, A.S. Mother rats bar-press for pups: Effects of lesions of the mpoa and limbic sites on maternal behavior and operant responding for pup-reinforcement. Behav. Brain Res. 1999, 100, 15–31. [Google Scholar] [CrossRef]
- Numan, M.; Insel, T.R. The Neurobiology of Parental Behavior; Springer Science & Business Media: Berlin, Germany, 2003; Volume 1. [Google Scholar]
- Rosenblatt, J.S.; Ceus, K. Estrogen implants in the medial preoptic area stimulate maternal behavior in male rats. Horm. Behav. 1998, 33, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Rosenblatt, J.S.; Olufowobi, A.; Siegel, H.I. Effects of pregnancy hormones on maternal responsiveness, responsiveness to estrogen stimulation of maternal behavior, and the lordosis response to estrogen stimulation. Horm. Behav. 1998, 33, 104–114. [Google Scholar] [CrossRef]
- Wu, Z.; Autry, A.E.; Bergan, J.F.; Watabe-Uchida, M.; Dulac, C.G. Galanin neurons in the medial preoptic area govern parental behaviour. Nature 2014, 509, 325–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohl, J.; Autry, A.E.; Dulac, C. The neurobiology of parenting: A neural circuit perspective. Bioessays 2017, 39, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kohl, J.; Dulac, C. Neural control of parental behaviors. Curr. Opin. Neurobiol. 2018, 49, 116–122. [Google Scholar] [CrossRef]
- Chen, C.; Song, I.Y.; Yuan, X.; Zhang, J. The thematic and citation landscape of data and knowledge engineering (1985–2007). Data Knowl. Eng. 2008, 67, 234–259. [Google Scholar] [CrossRef] [Green Version]
- Maia, S.C.; de Benedicto, G.C.; do Prado, J.W.; Robb, D.A.; de Almeida Bispo, O.N.; de Brito, M.J. Mapping the literature on credit unions: A bibliometric investigation grounded in Scopus and Web of Science. Scientometrics 2019, 120, 929–960. [Google Scholar] [CrossRef]
- Aryadoust, V.; Zakaria, A.; Lim, M.H.; Chen, C. An extensive knowledge mapping review of measurement and validity in language assessment and SLA research. Front. Psychol. 2020, 11, 1941. [Google Scholar] [CrossRef]
- Chen, C. CiteSpace: A Practical Guide for Mapping Scientific Literature; Nova Science Publishers Hauppauge: Harpak, NY, USA, 2016. [Google Scholar]
- Gaggero, G.; Bonassi, A.; Dellantonio, S.; Pastore, L.; Aryadoust, V.; Esposito, G. A scientometric review of alexithymia: Mapping thematic and disciplinary shifts in half a century of research. Front. Psychiatry 2020, 11, 1405. [Google Scholar] [CrossRef]
- Egghe, L. Theory and practise of the g-index. Scientometrics 2006, 69, 131–152. [Google Scholar] [CrossRef]
- Aryadoust, V.; Tan, H.A.H.; Ng, L.Y. A Scientometric review of Rasch measurement: The rise and progress of a specialty. Front. Psychol. 2019, 10, 2197. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Ibekwe-SanJuan, F.; Hou, J. The structure and dynamics of cocitation clusters: A multiple-perspective cocitation analysis. J. Am. Soc. Inf. Sci. Technol. 2010, 61, 1386–1409. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Song, M. Visualizing a field of research: A methodology of systematic scientometric reviews. PLoS ONE 2019, 14, e0223994. [Google Scholar] [CrossRef] [Green Version]
- Chen, C. Science mapping: A systematic review of the literature. J. Data Inf. Sci. 2017, 2, 1–40. [Google Scholar] [CrossRef] [Green Version]
- Kleinberg, J. Bursty and hierarchical structure in streams. Data Min. Knowl. Discov. 2003, 7, 373–397. [Google Scholar] [CrossRef]
- Numan, M.; Stolzenberg, D.S. Medial preoptic area interactions with dopamine neural systems in the control of the onset and maintenance of maternal behavior in rats. Front. Neuroendocrinol. 2009, 30, 46–64. [Google Scholar] [CrossRef]
- Numan, M.; Numan, M.J.; Schwarz, J.M.; Neuner, C.M.; Flood, T.F.; Smith, C.D. Medial preoptic area interactions with the nucleus accumbens–ventral pallidum circuit and maternal behavior in rats. Behav. Brain Res. 2005, 158, 53–68. [Google Scholar] [CrossRef]
- Champagne, F.A.; Chretien, P.; Stevenson, C.W.; Zhang, T.Y.; Gratton, A.; Meaney, M.J. Variations in nucleus accumbens dopamine associated with individual differences in maternal behavior in the rat. J. Neurosci. 2004, 24, 4113–4123. [Google Scholar] [CrossRef] [PubMed]
- Tsuneoka, Y.; Maruyama, T.; Yoshida, S.; Nishimori, K.; Kato, T.; Numan, M.; Kuroda, K.O. Functional, anatomical, and neurochemical differentiation of medial preoptic area subregions in relation to maternal behavior in the mouse. J. Comp. Neurol. 2013, 521, 1633–1663. [Google Scholar] [CrossRef]
- Numan, M. Hypothalamic neural circuits regulating maternal responsiveness toward infants. Behav. Cogn. Neurosci. Rev. 2006, 5, 163–190. [Google Scholar] [CrossRef] [Green Version]
- Numan, M.; Numan, M.J. Importance of pup-related sensory inputs and maternal performance for the expression of Fos-like immunoreactivity in the preoptic area and ventral bed nucleus of the stria terminalis of postpartum rats. Behav. Neurosci. 1995, 109, 135. [Google Scholar] [CrossRef] [PubMed]
- Dulac, C.; O’Connell, L.A.; Wu, Z. Neural control of maternal and paternal behaviors. Science 2014, 345, 765–770. [Google Scholar] [CrossRef] [Green Version]
- Shahrokh, D.K.; Zhang, T.Y.; Diorio, J.; Gratton, A.; Meaney, M.J. Oxytocin-dopamine interactions mediate variations in maternal behavior in the rat. Endocrinology 2010, 151, 2276–2286. [Google Scholar] [CrossRef]
- Pereira, M.; Morrell, J.I. Functional mapping of the neural circuitry of rat maternal motivation: Effects of site-specific transient neural inactivation. J. Neuroendocrinol. 2011, 23, 1020–1035. [Google Scholar] [CrossRef] [Green Version]
- Bridges, R.S. Neuroendocrine regulation of maternal behavior. Front. Neuroendocrinol. 2015, 36, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Stack, E.C.; Balakrishnan, R.; Numan, M.J.; Numan, M. A functional neuroanatomical investigation of the role of the medial preoptic area in neural circuits regulating maternal behavior. Behav. Brain Res. 2002, 131, 17–36. [Google Scholar] [CrossRef]
- Bosch, O.J.; Neumann, I.D. Both oxytocin and vasopressin are mediators of maternal care and aggression in rodents: From central release to sites of action. Horm. Behav. 2012, 61, 293–303. [Google Scholar] [CrossRef]
- Brown, J.R.; Ye, H.; Bronson, R.T.; Dikkes, P.; Greenberg, M.E. A defect in nurturing in mice lacking the immediate early gene fosB. Cell 1996, 86, 297–309. [Google Scholar] [CrossRef] [Green Version]
- Keer, S.; Stern, J. Dopamine receptor blockade in the nucleus accumbens inhibits maternal retrieval and licking, but enhances nursing behavior in lactating rats. Physiol. Behav. 1999, 67, 659–669. [Google Scholar] [CrossRef]
- Numan, M.; Numan, M.J. Expression of Fos-like immunoreactivity in the preoptic area of maternally behaving virgin and postpartum rats. Behav. Neurosci. 1994, 108, 379. [Google Scholar] [CrossRef]
- Numan, M. Motivational systems and the neural circuitry of maternal behavior in the rat. Dev. Psychobiol. J. Int. Soc. Dev. Psychobiol. 2007, 49, 12–21. [Google Scholar] [CrossRef]
- Meddle, S.L.; Bishop, V.R.; Gkoumassi, E.; van Leeuwen, F.W.; Douglas, A.J. Dynamic changes in oxytocin receptor expression and activation at parturition in the rat brain. Endocrinology 2007, 148, 5095–5104. [Google Scholar] [CrossRef]
- Tsuneoka, Y.; Tokita, K.; Yoshihara, C.; Amano, T.; Esposito, G.; Huang, A.J.; Yu, L.M.; Odaka, Y.; Shinozuka, K.; McHugh, T.J.; et al. Distinct preoptic-BST nuclei dissociate paternal and infanticidal behavior in mice. EMBO J. 2015, 34, 2652–2670. [Google Scholar] [CrossRef] [Green Version]
- Baum, M.; Everitt, B. Increased expression of c-fos in the medial preoptic area after mating in male rats: Role of afferent inputs from the medial amygdala and midbrain central tegmental field. Neuroscience 1992, 50, 627–646. [Google Scholar] [CrossRef]
- Numan, M.; Numan, M. Fos production in preoptic neurons correlated with different aspects of maternal behaviour in rats. Soc. Neurosci. Abs 1993, 19, 1020. [Google Scholar]
- Fleming, A.S.; Walsh, C. Neuropsychology of maternal behavior in the rat: C-fos expression during mother-litter interactions. Psychoneuroendocrinology 1994, 19, 429–443. [Google Scholar] [CrossRef]
- Fleming, A.S.; Korsmit, M. Plasticity in the maternal circuit: Effects of maternal experience on Fos-Lir in hypothalamic, limbic, and cortical structures in the postpartum rat. Behav. Neurosci. 1996, 110, 567. [Google Scholar] [CrossRef]
- Bridges, R.S.; Freemark, M.S. Human placental lactogen infusions into the medial preoptic area stimulate maternal behavior in steroid-primed, nulliparous female rats. Horm. Behav. 1995, 29, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Bridges, R.S.; Robertson, M.C.; Shiu, R.P.; Friesen, H.G.; Stuer, A.M.; Mann, P.E. Endocrine communication between conceptus and mother: Placental lactogen stimulation of maternal behavior. Neuroendocrinology 1996, 64, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Kinsley, C.H.; Bridges, R.S. Prolactin modulation of the maternal-like behavior displayed by juvenile rats. Horm. Behav. 1988, 22, 49–65. [Google Scholar] [CrossRef]
- Stern, J.M. Maternal behavior: Sensory, hormonal, and neural determinants. In Psychoendocrinology; Elsevier: Amsterdam, The Netherlands, 1989; pp. 105–226. [Google Scholar]
- Bridges, R.S.; Numan, M.; Ronsheim, P.M.; Mann, P.E.; Lupini, C.E. Central prolactin infusions stimulate maternal behavior in steroid-treated, nulliparous female rats. Proc. Natl. Acad. Sci. USA 1990, 87, 8003–8007. [Google Scholar] [CrossRef] [Green Version]
- Bridges, R.S.; Ronsheim, P.M. Prolactin (PRL) regulation of maternal behavior in rats: Bromocriptine treatment delays and PRL promotes the rapid onset of behavior. Endocrinology 1990, 126, 837–848. [Google Scholar] [CrossRef]
- Walsh, R.J.; Mangurian, L.P.; Posner, B.I. The distribution of lactogen receptors in the mammalian hypothalamus: An autoradiographic analysis of the rabbit and rat. Brain Res. 1990, 530, 1–11. [Google Scholar] [CrossRef]
- Emanuele, N.; Jurgens, J.; Halloran, M.; Tentler, J.; Lawrence, A.; Kelley, M. The rat prolactin gene is expressed in brain tissue: Detection of normal and alternatively spliced prolactin messenger RNA. Mol. Endocrinol. 1992, 6, 35–42. [Google Scholar] [PubMed]
- Hnasko, R.M.; Buntin, J.D. Functional mapping of neural sites mediating prolactin-induced hyperphagia in doves. Brain Res. 1993, 623, 257–266. [Google Scholar] [CrossRef]
- Buntin, J.D.; Ruzycki, E.; Witebsky, J. Prolactin receptors in dove brain: Autoradiographic analysis of binding characteristics in discrete brain regions and accessibility to blood-borne prolactin. Neuroendocrinology 1993, 57, 738–750. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Minoura, H.; Kawabe, N.; Tanaka, M.; Nakashima, K. Preferential expression of long form prolactin receptor mRNA in the rat brain during the oestrous cycle, pregnancy and lactation: Hormones involved in its gene expression. J. Endocrinol. 1994, 141, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Dutt, A.; Kaplitt, M.G.; Kow, L.M.; Pfaff, D.W. Prolactin, central nervous system and behavior: A critical review. Neuroendocrinology 1994, 59, 413–419. [Google Scholar] [CrossRef]
- Giordano, A.L.; Siegel, H.I.; Rosenblatt, J.S. Nuclear estrogen receptor binding in the preoptic area and hypothalamus of pregnancy-terminated rats: Correlation with the onset of maternal behavior. Neuroendocrinology 1989, 50, 248–258. [Google Scholar] [CrossRef]
- Giordano, A.L.; Siegel, H.I.; Rosenblatt, J.S. Nuclear estrogen receptor binding in microdissected brain regions of female rats during pregnancy: Implications for maternal and sexual behavior. Physiol. Behav. 1991, 50, 1263–1267. [Google Scholar] [CrossRef]
- Ehret, G.; Buckenmaier, J. Estrogen-receptor occurrence in the female mouse brain: Effects of maternal experience, ovariectomy, estrogen and anosmia. J. Physiol.-Paris 1994, 88, 315–329. [Google Scholar] [CrossRef]
- Rosenblatt, J.S.; Hazelwood, S.; Poole, J. Maternal behavior in male rats: Effects of medial preoptic area lesions and presence of maternal aggression. Horm. Behav. 1996, 30, 201–215. [Google Scholar] [CrossRef]
- Cintra, A.; Fuxe, K.; Harfstrand, A.; Agnati, L.; Miller, L. On the cellular localization and distribution of estrogen receptors in the rat tel-and diencephalon using monoclonal antibodies to human estrogen receptor. Neurochem. Int. 1986, 8, 587–595. [Google Scholar] [CrossRef]
- Balthazart, J.; Foidart, A.; Surlemont, C.; Harada, N.; Naftolin, F. Neuroanatomical specificity in the autoregulation of aromatase-immunoreactive neurons by androgens and estrogens: An immunocytochemical study. Brain Res. 1992, 574, 280–290. [Google Scholar] [CrossRef]
- Bloch, G.; Gorski, R. Cytoarchitectonic analysis of the SDN-POA of the intact and gonadectomized rat. J. Comp. Neurol. 1988, 275, 604–612. [Google Scholar] [CrossRef]
- Simerly, R.; Swanson, L.; Chang, C.; Muramatsu, M. Distribution of androgen and estrogen receptor mRNA-containing cells in the rat brain: An in situ hybridization study. J. Comp. Neurol. 1990, 294, 76–95. [Google Scholar] [CrossRef]
- Rhees, R.W.; Shryne, J.E.; Gorski, R.A. Termination of the hormone-sensitive period for differentiation of the sexually dimorphic nucleus of the preoptic area in male and female rats. Dev. Brain Res. 1990, 52, 17–23. [Google Scholar] [CrossRef]
- Sanghera, M.K.; Simpson, E.R.; McPHAUL, M.J.; Kozlowski, G.; Conley, A.J.; Lephart, E.D. Immunocytochemical distribution of aromatase cytochrome P450 in the rat brain using peptide-generated polyclonal antibodies. Endocrinology 1991, 129, 2834–2844. [Google Scholar] [CrossRef]
- Balthazart, J.; Foidart, A.; Surlemont, C.; Harada, N. Distribution of aromatase-immunoreactive cells in the mouse forebrain. Cell Tissue Res. 1991, 263, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Balthazart, J.; Foidart, A.; Surlemont, C.; Harada, N. Neuroanatomical specificity in the co-localization of aromatase and estrogen receptors. J. Neurobiol. 1991, 22, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Dellovade, T.; Rissman, E.; Thompson, N.; Harada, N.; Ottinger, M. Co-localization of aromatase enzyme and estrogen receptor immunoreactivity in the preoptic area during reproductive aging. Brain Res. 1995, 674, 181–187. [Google Scholar] [CrossRef]
- Nagano, M.; Shinoda, K. Coexistence of the stigmoid body and estrogen receptor in some neuronal groups involved in rat reproductive functions. Brain Res. 1994, 634, 296–304. [Google Scholar] [CrossRef]
- Rizvi, T.A.; Ennis, M.; Shipley, M.T. Reciprocal connections between the medial preoptic area and the midbrain periaqueductal gray in rat: A WGA-HRP and PHA-L study. J. Comp. Neurol. 1992, 315, 1–15. [Google Scholar] [CrossRef]
- Paut-Pagano, L.; Roky, R.; Valatx, J.L.; Kitahama, K.; Jouvet, M. Anatomical distribution of prolactin-like immunoreactivity in the rat brain. Neuroendocrinology 1993, 58, 682–695. [Google Scholar] [CrossRef]
- Bridges, R.S.; Mann, P.E. Prolactin-brain interactions in the induction of maternal behavior in rats. Psychoneuroendocrinology 1994, 19, 611–622. [Google Scholar] [CrossRef]
- Numan, M. A neural circuitry analysis of maternal behavior in the rat. Acta Paediatr. 1994, 83, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Roky, R.; Paut-Pagano, L.; Goffin, V.; Kitahama, K.; Valatx, J.L.; Kelly, P.A.; Jouvet, M. Distribution of prolactin receptors in the rat forebrain. Neuroendocrinology 1996, 63, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Bakowska, J.C.; Morrell, J.I. Atlas of the neurons that express mRNA for the long form of the prolactin receptor in the forebrain of the female rat. J. Comp. Neurol. 1997, 386, 161–177. [Google Scholar] [CrossRef]
- Bole-Feysot, C.; Goffin, V.; Edery, M.; Binart, N.; Kelly, P.A. Prolactin (PRL) and its receptor: Actions, signal transduction pathways and phenotypes observed in PRL receptor knockout mice. Endocr. Rev. 1998, 19, 225–268. [Google Scholar] [CrossRef]
- Pi, X.J.; Grattan, D.R. Distribution of prolactin receptor immunoreactivity in the brain of estrogen-treated, ovariectomized rats. J. Comp. Neurol. 1998, 394, 462–474. [Google Scholar] [CrossRef]
- Pi, X.; Grattan, D. Increased expression of both short and long forms of prolactin receptor mRNA in hypothalamic nuclei of lactating rats. J. Mol. Endocrinol. 1999, 23, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argiolas, A.; Gessa, G.L. Central functions of oxytocin. Neurosci. Biobehav. Rev. 1991, 15, 217–231. [Google Scholar] [CrossRef]
- Crowley, W.R.; Armstrong, W.E. Neurochemical regulation of oxytocin secretion in lactation. Endocr. Rev. 1992, 13, 33–65. [Google Scholar]
- Yoshimura, R.; Kiyama, H.; Kimura, T.; Araki, T.; Maeno, H.; Tanizawa, O.; Tohyama, M. Localization of oxytocin receptor messenger ribonucleic acid in the rat brain. Endocrinology 1993, 133, 1239–1246. [Google Scholar] [CrossRef]
- Pedersen, C.A.; Caldwell, J.D.; Walker, C.; Ayers, G.; Mason, G.A. Oxytocin activates the postpartum onset of rat maternal behavior in the ventral tegmental and medial preoptic areas. Behav. Neurosci. 1994, 108, 1163–1171. [Google Scholar] [CrossRef]
- Corodimas, K.P.; Rosenblatt, J.S.; Morrell, J.I. The habenular complex mediates hormonal stimulation of maternal behavior in rats. Behav. Neurosci. 1992, 106, 853. [Google Scholar] [CrossRef]
- Corodimas, K.P.; Rosenblatt, J.S.; Canfield, M.E.; Morrell, J.I. Neurons in the lateral subdivision of the habenular complex mediate the hormonal onset of maternal behavior in rats. Behav. Neurosci. 1993, 107, 827. [Google Scholar] [CrossRef]
- Matthews-Felton, T.; Corodimas, K.P.; Rosenblatt, J.S.; Morrell, J.I. Lateral habenula neurons are necessary for the hormonal onset of maternal behavior and for the display of postpartum estrus in naturally parturient female rats. Behav. Neurosci. 1995, 109, 1172. [Google Scholar] [CrossRef]
- Fleming, A.S.; Suh, E.J.; Korsmit, M.; Rusak, B. Activation of Fos-like immunoreactivity in the medial preoptic area and limbic structures of maternal and social interactions in rats. Behav. Neurosci. 1994, 108, 724. [Google Scholar] [CrossRef]
- Kirkpatrick, B.; Kim, J.W.; Insel, T.R. Limbic system fos expression associated with paternal behavior. Brain Res. 1994, 658, 112–118. [Google Scholar] [CrossRef]
- Lonstein, J.; Simmons, D.; Swann, J.; Stern, J. Forebrain expression of c-fos due to active maternal behaviour in lactating rats. Neuroscience 1997, 82, 267–281. [Google Scholar] [CrossRef]
- Numan, M.; Numan, M.J.; Marzella, S.R.; Palumbo, A. Expression of c-fos, fos B, and egr-1 in the medial preoptic area and bed nucleus of the stria terminalis during maternal behavior in rats. Brain Res. 1998, 792, 348–352. [Google Scholar] [CrossRef]
- Li, C.; Chen, P.; Smith, M. Neural populations in the rat forebrain and brainstem activated by the suckling stimulus as demonstrated by cFos expression. Neuroscience 1999, 94, 117–129. [Google Scholar] [CrossRef]
- Kalinichev, M.; Rosenblatt, J.S.; Nakabeppu, Y.; Morrell, J.I. Induction of c-fos-like and fosB-like immunoreactivity reveals forebrain neuronal populations involved differentially in pup-mediated maternal behavior in juvenile and adult rats. J. Comp. Neurol. 2000, 416, 45–78. [Google Scholar] [CrossRef]
- Morgan, J.I.; Curran, T. Stimulus-transcription coupling in the nervous system: Involvement of the inducible proto-oncogenes fos and jun. Annu. Rev. Neurosci. 1991, 14, 421–451. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, G.E.; Lee, W.S.; Smith, M.S.; Abbud, R. C-Fos and Fos-Related Antigens as Markers for Neuronal Activity: Perspectives From Neuroendocrine. Act. Immed. Early Genes Drugs Abus. 1993, 125, 117. [Google Scholar]
- Hoffman, G.E.; Smith, M.S.; Verbalis, J.G. c-Fos and related immediate early gene products as markers of activity in neuroendocrine systems. Front. Neuroendocrinol. 1993, 14, 173–213. [Google Scholar] [CrossRef]
- Bridges, R.S. Biochemical basis of parental behavior in the rat. In Advances in the Study of Behavior; Elsevier: Amsterdam, The Netherlands, 1996; Volume 25, pp. 215–242. [Google Scholar]
- Stack, E.C.; Numan, M. The temporal course of expression of c-Fos and Fos B within the medial preoptic area and other brain regions of postpartum female rats during prolonged mother–young interactions. Behav. Neurosci. 2000, 114, 609. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.H.; Miyata, S.; Weng, W.; Matsunaga, W.; Ichikawa, J.; Furuya, K.; Nakashima, T.; Kiyohara, T. Comparison of the expression of two immediate early gene proteins, FosB and Fos in the rat preoptic area, hypothalamus and brainstem during pregnancy, parturition and lactation. Neurosci. Res. 1998, 32, 333–341. [Google Scholar] [CrossRef]
- Walsh, C.J.; Fleming, A.S.; Lee, A.; Magnusson, J.E. The effects of olfactory and somatosensory desensitization on Fos-like immunoreactivity in the brains of pup-exposed postpartum rats. Behav. Neurosci. 1996, 110, 134. [Google Scholar] [CrossRef] [PubMed]
- Lonstein, J.; De Vries, G. Maternal behaviour in lactating rats stimulates c-fos in glutamate decarboxylase-synthesizing neurons of the medial preoptic area, ventral bed nucleus of the stria terminalis, and ventrocaudal periaqueductal gray. Neuroscience 2000, 100, 557–568. [Google Scholar] [CrossRef]
- Bridges, R.; Mann, P.; Coppeta, J. Hypothalamic involvement in the regulation of maternal behaviour in the rat: Inhibitory roles for the ventromedial hypothalamus and the dorsal/anterior hypothalamic areas. J. Neuroendocrinol. 1999, 11, 259–266. [Google Scholar] [CrossRef]
- Sheehan, T.P.; Cirrito, J.; Numan, M.J.; Numan, M. Using c-Fos immunocytochemistry to identify forebrain regions that may inhibit maternal behavior in rats. Behav. Neurosci. 2000, 114, 337. [Google Scholar] [CrossRef]
- Komisaruk, B.R.; Rosenblatt, J.S.; Barona, M.L.; Chinapen, S.; Nissanov, J.; T O’Bannon III, R.; Johnson, B.M.; Del Cerro, M.C.R. Combined c-fos and 14C-2-deoxyglucose method to differentiate site-specific excitation from disinhibition: Analysis of maternal behavior in the rat. Brain Res. 2000, 859, 262–272. [Google Scholar] [CrossRef]
- Lonstein, J.S.; Gréco, B.; De Vries, G.J.; Stern, J.M.; Blaustein, J.D. Maternal behavior stimulates c-fos activity within estrogen receptor alpha-containing neurons in lactating rats. Neuroendocrinology 2000, 72, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, T.; Numan, M. Estrogen, progesterone, and pregnancy termination alter neural activity in brain regions that control maternal behavior in rats. Neuroendocrinology 2002, 75, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Insel, T.R. Regional induction of c-fos-like protein in rat brain after estradiol administration. Endocrinology 1990, 126, 1849–1853. [Google Scholar] [CrossRef]
- Cattaneo, E.; Maggi, A. C-fos induction by estrogen in specific rat brain areas. Eur. J. Pharmacol. Mol. Pharmacol. 1990, 188, 153–159. [Google Scholar] [CrossRef]
- Mayer, A.D.; Monroy, M.A.; Rosenblatt, J.S. Prolonged estrogen-progesterone treatment of nonpregnant ovariectomized rats: Factors stimulating home-cage and maternal aggression and short-latency maternal behavior. Horm. Behav. 1990, 24, 342–364. [Google Scholar] [CrossRef]
- Grattan, D.R. The actions of prolactin in the brain during pregnancy and lactation. Prog. Brain Res. 2001, 133, 153–171. [Google Scholar] [PubMed]
- Gammie, S.C. Current models and future directions for understanding the neural circuitries of maternal behaviors in rodents. Behav. Cogn. Neurosci. Rev. 2005, 4, 119–135. [Google Scholar] [CrossRef]
- Arrati, P.G.; Carmona, C.; Dominguez, G.; Beyer, C.; Rosenblatt, J.S. GABA receptor agonists in the medial preoptic area and maternal behavior in lactating rats. Physiol. Behav. 2006, 87, 51–65. [Google Scholar] [CrossRef]
- Olazabal, D.E.; Young, L.J. Oxytocin receptors in the nucleus accumbens facilitate “spontaneous” maternal behavior in adult female prairie voles. Neuroscience 2006, 141, 559–568. [Google Scholar] [CrossRef]
- Broad, K.; Lévy, F.; Evans, G.; Kimura, T.; Keverne, E.; Kendrick, K. Previous maternal experience potentiates the effect of parturition on oxytocin receptor mRNA expression in the paraventricular nucleus. Eur. J. Neurosci. 1999, 11, 3725–3737. [Google Scholar] [CrossRef]
- Francis, D.D.; Champagne, F.C.; Meaney, M.J. Variations in maternal behaviour are associated with differences in oxytocin receptor levels in the rat. J. Neuroendocrinol. 2000, 12, 1145–1148. [Google Scholar] [CrossRef] [PubMed]
- Champagne, F.; Diorio, J.; Sharma, S.; Meaney, M.J. Naturally occurring variations in maternal behavior in the rat are associated with differences in estrogen-inducible central oxytocin receptors. Proc. Natl. Acad. Sci. USA 2001, 98, 12736–12741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lightman, S.L.; Windle, R.J.; Wood, S.A.; Kershaw, Y.M.; Shanks, N.; Ingram, C.D. Peripartum plasticity within the hypothalamo-pituitary-adrenal axis. Prog. Brain Res. 2001, 133, 111–129. [Google Scholar]
- Silva, M.R.P.; Bernardi, M.M.; Cruz-Casallas, P.E.; Felicio, L.F. Pimozide injections into the nucleus accumbens disrupt maternal behaviour in lactating rats. Pharmacol. Toxicol. 2003, 93, 42–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Numan, M.; Numan, M.J.; Pliakou, N.; Stolzenberg, D.S.; Mullins, O.J.; Murphy, J.M.; Smith, C.D. The effects of D1 or D2 dopamine receptor antagonism in the medial preoptic area, ventral pallidum, or nucleus accumbens on the maternal retrieval response and other aspects of maternal behavior in rats. Behav. Neurosci. 2005, 119, 1588. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.M.; Lonstein, J.S. Dopamine d1 and d2 receptor antagonism in the preoptic area produces different effects on maternal behavior in lactating rats. Behav. Neurosci. 2005, 119, 1072. [Google Scholar] [CrossRef]
- Li, M.; Fleming, A.S. The nucleus accumbens shell is critical for normal expression of pup-retrieval in postpartum female rats. Behav. Brain Res. 2003, 145, 99–111. [Google Scholar] [CrossRef]
- Lee, A.; Li, M.; Watchus, J.; Fleming, A.S. Neuroanatomical basis of maternal memory in postpartum rats: Selective role for the nucleus accumbens. Behav. Neurosci. 1999, 113, 523. [Google Scholar] [CrossRef]
- Li, M.; Fleming, A.S. Differential involvement of nucleus accumbens shell and core subregions in maternal memory in postpartum female rats. Behav. Neurosci. 2003, 117, 426. [Google Scholar] [CrossRef] [PubMed]
- Lonstein, J.S. Effects of dopamine receptor antagonism with haloperidol on nurturing behavior in the biparental prairie vole. Pharmacol. Biochem. Behav. 2002, 74, 11–19. [Google Scholar] [CrossRef]
- Afonso, V.M.; King, S.; Chatterjee, D.; Fleming, A.S. Hormones that increase maternal responsiveness affect accumbal dopaminergic responses to pup-and food-stimuli in the female rat. Horm. Behav. 2009, 56, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Vernotica, E.M.; Rosenblatt, J.S.; Morrell, J.I. Microinfusion of cocaine into the medial preoptic area or nucleus accumbens transiently impairs maternal behavior in the rat. Behav. Neurosci. 1999, 113, 377. [Google Scholar] [CrossRef]
- Insel, T.R. Is social attachment an addictive disorder? Physiol. Behav. 2003, 79, 351–357. [Google Scholar] [CrossRef]
- Ferris, C.F.; Kulkarni, P.; Sullivan, J.M.; Harder, J.A.; Messenger, T.L.; Febo, M. Pup suckling is more rewarding than cocaine: Evidence from functional magnetic resonance imaging and three-dimensional computational analysis. J. Neurosci. 2005, 25, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Seip, K.M.; Morrell, J.I. Increasing the incentive salience of cocaine challenges preference for pup-over cocaine-associated stimuli during early postpartum: Place preference and locomotor analyses in the lactating female rat. Psychopharmacology 2007, 194, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Stolzenberg, D.S.; McKenna, J.B.; Keough, S.; Hancock, R.; Numan, M.J.; Numan, M. Dopamine D1 receptor stimulation of the nucleus accumbens or the medial preoptic area promotes the onset of maternal behavior in pregnancy-terminated rats. Behav. Neurosci. 2007, 121, 907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Numan, M.; Stolzenberg, D.S. Hypothalamic interaction with the mesolimbic dopamine system and the regulation of maternal responsiveness. In Neurobiology of the Parental Brain; Academic Press: Cambridge, MA, USA, 2008; pp. 3–22. [Google Scholar]
- Pereira, M.; Ferreira, A. Demanding pups improve maternal behavioral impairments in sensitized and haloperidol-treated lactating female rats. Behav. Brain Res. 2006, 175, 139–148. [Google Scholar] [CrossRef]
- Bardo, M.T.; Donohew, R.; Harrington, N.G. Psychobiology of novelty seeking and drug seeking behavior. Behav. Brain Res. 1996, 77, 23–43. [Google Scholar] [CrossRef]
- Berridge, K.C.; Robinson, T.E. What is the role of dopamine in reward: Hedonic impact, reward learning, or incentive salience? Brain Res. Rev. 1998, 28, 309–369. [Google Scholar] [CrossRef]
- Ikemoto, S.; Panksepp, J. The role of nucleus accumbens dopamine in motivated behavior: A unifying interpretation with special reference to reward-seeking. Brain Res. Rev. 1999, 31, 6–41. [Google Scholar] [CrossRef]
- Becker, J.B. Gender differences in dopaminergic function in striatum and nucleus accumbens. Pharmacol. Biochem. Behav. 1999, 64, 803–812. [Google Scholar] [CrossRef]
- Hunt, G.E.; McGregor, I.S. Contrasting effects of dopamine antagonists and frequency reduction on Fos expression induced by lateral hypothalamic stimulation. Behav. Brain Res. 2002, 132, 187–201. [Google Scholar] [CrossRef]
- Reynolds, S.M.; Berridge, K.C. Positive and negative motivation in nucleus accumbens shell: Bivalent rostrocaudal gradients for GABA-elicited eating, taste “liking”/“disliking” reactions, place preference/avoidance, and fear. J. Neurosci. 2002, 22, 7308–7320. [Google Scholar] [CrossRef]
- Horvitz, J.C. Dopamine gating of glutamatergic sensorimotor and incentive motivational input signals to the striatum. Behav. Brain Res. 2002, 137, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Olazabal, D.; Abercrombie, E.; Rosenblatt, J.; Morrell, J. The content of dopamine, serotonin, and their metabolites in the neural circuit that mediates maternal behavior in juvenile and adult rats. Brain Res. Bull. 2004, 63, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Bosch, O.; Pförtsch, J.; Beiderbeck, D.; Landgraf, R.; Neumann, I. Maternal behaviour is associated with vasopressin release in the medial preoptic area and bed nucleus of the stria terminalis in the rat. J. Neuroendocrinol. 2010, 22, 420–429. [Google Scholar] [CrossRef]
- Anderson, G.M.; Grattan, D.R.; van den Ancker, W.; Bridges, R.S. Reproductive experience increases prolactin responsiveness in the medial preoptic area and arcuate nucleus of female rats. Endocrinology 2006, 147, 4688–4694. [Google Scholar] [CrossRef]
- Bridges, R.S.; Robertson, M.C.; Shiu, R.P.; Sturgis, J.D.; Henriquez, B.M.; Mann, P.E. Central lactogenic regulation of maternal behavior in rats: Steroid dependence, hormone specificity, and behavioral potencies of rat prolactin and rat placental lactogen I. Endocrinology 1997, 138, 756–763. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.A.; Douglas, A.J.; Ingram, C.D. Brain preparations for maternity—adaptive changes in behavioral and neuroendocrine systems during pregnancy and lactation. An overview. Prog. Brain Res. 2001, 133, 1–38. [Google Scholar]
- Bridges, R.S.; Rigero, B.A.; Byrnes, E.M.; Yang, L.; Walker, A.M. Central infusions of the recombinant human prolactin receptor antagonist, S179D-PRL, delay the onset of maternal behavior in steroid-primed, nulliparous female rats. Endocrinology 2001, 142, 730–739. [Google Scholar] [CrossRef] [PubMed]
- Leckman, J.F.; Herman, A.E. Maternal behavior and developmental psychopathology. Biol. Psychiatry 2002, 51, 27–43. [Google Scholar] [CrossRef]
- Torner, L.; Toschi, N.; Nava, G.; Clapp, C.; Neumann, I.D. Increased hypothalamic expression of prolactin in lactation: Involvement in behavioural and neuroendocrine stress responses. Eur. J. Neurosci. 2002, 15, 1381–1389. [Google Scholar] [CrossRef]
- Wettschureck, N.; Moers, A.; Hamalainen, T.; Lemberger, T.; Schütz, G.; Offermanns, S. Heterotrimeric G proteins of the Gq/11 family are crucial for the induction of maternal behavior in mice. Mol. Cell. Biol. 2004, 24, 8048–8054. [Google Scholar] [CrossRef] [Green Version]
- Bridges, R.S.; Hays, L.E. Steroid-induced alterations in mRNA expression of the long form of the prolactin receptor in the medial preoptic area of female rats: Effects of exposure to a pregnancy-like regimen of progesterone and estradiol. Mol. Brain Res. 2005, 140, 10–16. [Google Scholar] [CrossRef]
- Grattan, D.R.; Steyn, F.J.; Kokay, I.C.; Anderson, G.M.; Bunn, S.J. Pregnancy-induced adaptation in the neuroendocrine control of prolactin secretion. J. Neuroendocrinol. 2008, 20, 497–507. [Google Scholar] [CrossRef]
- Gammie, S.C.; Nelson, R.J. Maternal aggression is reduced in neuronal nitric oxide synthase-deficient mice. J. Neurosci. 1999, 19, 8027–8035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lonstein, J.S.; Gammie, S.C. Sensory, hormonal, and neural control of maternal aggression in laboratory rodents. Neurosci. Biobehav. Rev. 2002, 26, 869–888. [Google Scholar] [CrossRef]
- Gammie, S.C.; Negron, A.; Newman, S.M.; Rhodes, J.S. Corticotropin-releasing factor inhibits maternal aggression in mice. Behav. Neurosci. 2004, 118, 805. [Google Scholar] [CrossRef] [Green Version]
- Popeski, N.; Woodside, B. Central nitric oxide synthase inhibition disrupts maternal behavior in the rat. Behav. Neurosci. 2004, 118, 1305. [Google Scholar] [CrossRef] [PubMed]
- Numan, M. Maternal behaviors: Central integration or independent parallel circuits? Theoretical comment on Popeski and Woodside (2004). Behav. Neurosci. 2004, 118, 1469–1472. [Google Scholar] [CrossRef]
- Bosch, O.J.; Meddle, S.L.; Beiderbeck, D.I.; Douglas, A.J.; Neumann, I.D. Brain oxytocin correlates with maternal aggression: Link to anxiety. J. Neurosci. 2005, 25, 6807–6815. [Google Scholar] [CrossRef] [Green Version]
- Lonstein, J.; Dominguez, J.; Putnam, S.; De Vries, G.; Hull, E. Intracellular preoptic and striatal monoamines in pregnant and lactating rats: Possible role in maternal behavior. Brain Res. 2003, 970, 149–158. [Google Scholar] [CrossRef]
- Miller, S.M.; Lonstein, J.S. Dopaminergic projections to the medial preoptic area of postpartum rats. Neuroscience 2009, 159, 1384–1396. [Google Scholar] [CrossRef] [Green Version]
- Numan, M.; Numan, M.J. Projection sites of medial preoptic area and ventral bed nucleus of the stria terminalis neurons that express Fos during maternal behavior in female rats. J. Neuroendocrinol. 1997, 9, 369–384. [Google Scholar] [CrossRef] [PubMed]
- Kudo, T.; Uchigashima, M.; Miyazaki, T.; Konno, K.; Yamasaki, M.; Yanagawa, Y.; Minami, M.; Watanabe, M. Three types of neurochemical projection from the bed nucleus of the stria terminalis to the ventral tegmental area in adult mice. J. Neurosci. 2012, 32, 18035–18046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jennings, J.H.; Sparta, D.R.; Stamatakis, A.M.; Ung, R.L.; Pleil, K.E.; Kash, T.L.; Stuber, G.D. Distinct extended amygdala circuits for divergent motivational states. Nature 2013, 496, 224–228. [Google Scholar] [CrossRef]
- Numan, M.; Stolzenberg, D.S.; Dellevigne, A.A.; Correnti, C.M.; Numan, M.J. Temporary inactivation of ventral tegmental area neurons with either muscimol or baclofen reversibly disrupts maternal behavior in rats through different underlying mechanisms. Behav. Neurosci. 2009, 123, 740. [Google Scholar] [CrossRef] [Green Version]
- Seip, K.M.; Morrell, J.I. Transient inactivation of the ventral tegmental area selectively disrupts the expression of conditioned place preference for pup-but not cocaine-paired contexts. Behav. Neurosci. 2009, 123, 1325. [Google Scholar] [CrossRef] [Green Version]
- Febo, M.; Felix-Ortiz, A.C.; Johnson, T.R. Inactivation or inhibition of neuronal activity in the medial prefrontal cortex largely reduces pup retrieval and grouping in maternal rats. Brain Res. 2010, 1325, 77–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Numan, M.; Woodside, B. Maternity: Neural mechanisms, motivational processes, and physiological adaptations. Behav. Neurosci. 2010, 124, 715. [Google Scholar] [CrossRef] [PubMed]
- Mattson, B.J.; Williams, S.E.; Rosenblatt, J.S.; Morrell, J.I. Preferences for cocaine-or pup-associated chambers differentiates otherwise behaviorally identical postpartum maternal rats. Psychopharmacology 2003, 167, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Curtis, J.T.; Liu, Y.; Aragona, B.J.; Wang, Z. Dopamine and monogamy. Brain Res. 2006, 1126, 76–90. [Google Scholar] [CrossRef]
- Perrin, G.; Meurisse, M.; Lévy, F. Inactivation of the medial preoptic area or the bed nucleus of the stria terminalis differentially disrupts maternal behavior in sheep. Horm. Behav. 2007, 52, 461–473. [Google Scholar] [CrossRef]
- Rutherford, H.; Williams, S.; Moy, S.; Mayes, L.; Johns, J. Disruption of maternal parenting circuitry by addictive process: Rewiring of reward and stress systems. Front. Psychiatry 2011, 2, 37. [Google Scholar] [CrossRef] [Green Version]
- Lavi-Avnon, Y.; Weller, A.; Finberg, J.P.; Gispan-Herman, I.; Kinor, N.; Stern, Y.; Schroeder, M.; Gelber, V.; Bergman, S.Y.; Overstreet, D.H.; et al. The reward system and maternal behavior in an animal model of depression: A microdialysis study. Psychopharmacology 2008, 196, 281–291. [Google Scholar] [CrossRef]
- Stolzenberg, D.S.; Numan, M. Hypothalamic interaction with the mesolimbic DA system in the control of the maternal and sexual behaviors in rats. Neurosci. Biobehav. Rev. 2011, 35, 826–847. [Google Scholar] [CrossRef] [PubMed]
- Mattson, B.; Morrell, J. Preference for cocaine-versus pup-associated cues differentially activates neurons expressing either Fos or cocaine-and amphetamine-regulated transcript in lactating, maternal rodents. Neuroscience 2005, 135, 315–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wansaw, M.P.; Pereira, M.; Morrell, J.I. Characterization of maternal motivation in the lactating rat: Contrasts between early and late postpartum responses. Horm. Behav. 2008, 54, 294–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, M.; Morrell, J.I. The medial preoptic area is necessary for motivated choice of pup-over cocaine-associated environments by early postpartum rats. Neuroscience 2010, 167, 216–231. [Google Scholar] [CrossRef] [Green Version]
- Dölen, G.; Darvishzadeh, A.; Huang, K.W.; Malenka, R.C. Social reward requires coordinated activity of nucleus accumbens oxytocin and serotonin. Nature 2013, 501, 179–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rilling, J.K. The neural and hormonal bases of human parentalcare. Neuropsychologia 2013, 51, 731–747. [Google Scholar] [CrossRef]
- Stolzenberg, D.S.; Rissman, E.F. Oestrogen-independent, experience-induced maternal behaviour in female mice. J. Neuroendocrinol. 2011, 23, 345–354. [Google Scholar] [CrossRef] [Green Version]
- Afonso, V.M.; Grella, S.L.; Chatterjee, D.; Fleming, A.S. Previous maternal experience affects accumbal dopaminergic responses to pup-stimuli. Brain Res. 2008, 1198, 115–123. [Google Scholar] [CrossRef]
- Parada, M.; King, S.; Li, M.; Fleming, A.S. The roles of accumbal dopamine D1 and D2 receptors in maternal memory in rats. Behav. Neurosci. 2008, 122, 368. [Google Scholar] [CrossRef] [Green Version]
- Olazábal, D.E.; Pereira, M.; Agrati, D.; Ferreira, A.; Fleming, A.S.; González-Mariscal, G.; Lévy, F.; Lucion, A.B.; Morrell, J.I.; Numan, M.; et al. Flexibility and adaptation of the neural substrate that supports maternal behavior in mammals. Neurosci. Biobehav. Rev. 2013, 37, 1875–1892. [Google Scholar] [CrossRef]
- Pereira, M.; Morrell, J.I. The changing role of the medial preoptic area in the regulation of maternal behavior across the postpartum period: Facilitation followed by inhibition. Behav. Brain Res. 2009, 205, 238–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rilling, J.K.; Young, L.J. The biology of mammalian parenting and its effect on offspring social development. Science 2014, 345, 771–776. [Google Scholar] [CrossRef] [Green Version]
- Lonstein, J.S.; Lévy, F.; Fleming, A.S. Common and divergent psychobiological mechanisms underlying maternal behaviors in non-human and human mammals. Horm. Behav. 2015, 73, 156–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McHenry, J.A.; Rubinow, D.R.; Stuber, G.D. Maternally responsive neurons in the bed nucleus of the stria terminalis and medial preoptic area: Putative circuits for regulating anxiety and reward. Front. Neuroendocrinol. 2015, 38, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunton, P.J.; Russell, J.A. The expectant brain: Adapting for motherhood. Nat. Rev. Neurosci. 2008, 9, 11–25. [Google Scholar] [CrossRef]
- Schiller, C.E.; O’Hara, M.W.; Rubinow, D.R.; Johnson, A.K. Estradiol modulates anhedonia and behavioral despair in rats and negative affect in a subgroup of women at high risk for postpartum depression. Physiol. Behav. 2013, 119, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Walker, D.L.; Toufexis, D.J.; Davis, M. Role of the bed nucleus of the stria terminalis versus the amygdala in fear, stress, and anxiety. Eur. J. Pharmacol. 2003, 463, 199–216. [Google Scholar] [CrossRef]
- Nephew, B.C.; Bridges, R.S. Arginine vasopressin V1a receptor antagonist impairs maternal memory in rats. Physiol. Behav. 2008, 95, 182–186. [Google Scholar] [CrossRef] [Green Version]
- d’Cunha, T.; King, S.; Fleming, A.; Lévy, F. Oxytocin receptors in the nucleus accumbens shell are involved in the consolidation of maternal memory in postpartum rats. Horm. Behav. 2011, 59, 14–21. [Google Scholar] [CrossRef]
- Scanlan, V.F.; Byrnes, E.M.; Bridges, R.S. Reproductive experience and activation of maternal memory. Behav. Neurosci. 2006, 120, 676. [Google Scholar] [CrossRef] [PubMed]
- Vecsey, C.G.; Hawk, J.D.; Lattal, K.M.; Stein, J.M.; Fabian, S.A.; Attner, M.A.; Cabrera, S.M.; McDonough, C.B.; Brindle, P.K.; Abel, T.; et al. Histone deacetylase inhibitors enhance memory and synaptic plasticity via CREB: CBP-dependent transcriptional activation. J. Neurosci. 2007, 27, 6128–6140. [Google Scholar] [CrossRef]
- Brusco, J.; Wittmann, R.; de Azevedo, M.S.; Lucion, A.B.; Franci, C.R.; Giovenardi, M.; Rasia-Filho, A.A. Plasma hormonal profiles and dendritic spine density and morphology in the hippocampal CA1 stratum radiatum, evidenced by light microscopy, of virgin and postpartum female rats. Neurosci. Lett. 2008, 438, 346–350. [Google Scholar] [CrossRef]
- Kim, P.; Leckman, J.F.; Mayes, L.C.; Feldman, R.; Wang, X.; Swain, J.E. The plasticity of human maternal brain: Longitudinal changes in brain anatomy during the early postpartum period. Behav. Neurosci. 2010, 124, 695. [Google Scholar] [CrossRef] [Green Version]
- Leuner, B.; Gould, E. Dendritic growth in medial prefrontal cortex and cognitive flexibility are enhanced during the postpartum period. J. Neurosci. 2010, 30, 13499–13503. [Google Scholar] [CrossRef] [Green Version]
- Shams, S.; Pawluski, J.L.; Chatterjee-Chakraborty, M.; Oatley, H.; Mastroianni, A.; Fleming, A.S. Dendritic morphology in the striatum and hypothalamus differentially exhibits experience-dependent changes in response to maternal care and early social isolation. Behav. Brain Res. 2012, 233, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Pawluski, J.L.; Galea, L.A. Hippocampal morphology is differentially affected by reproductive experience in the mother. J. Neurobiol. 2006, 66, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Pawluski, J.L.; Brummelte, S.; Barha, C.K.; Crozier, T.M.; Galea, L.A. Effects of steroid hormones on neurogenesis in the hippocampus of the adult female rodent during the estrous cycle, pregnancy, lactation and aging. Front. Neuroendocrinol. 2009, 30, 343–357. [Google Scholar] [CrossRef]
- Barrett, J.; Fleming, A.S. Annual research review: All mothers are not created equal: Neural and psychobiological perspectives on mothering and the importance of individual differences. J. Child Psychol. Psychiatry 2011, 52, 368–397. [Google Scholar] [CrossRef]
- Moses-Kolko, E.L.; Fraser, D.; Wisner, K.L.; James, J.A.; Saul, A.T.; Fiez, J.A.; Phillips, M.L. Rapid habituation of ventral striatal response to reward receipt in postpartum depression. Biol. Psychiatry 2011, 70, 395–399. [Google Scholar] [CrossRef] [Green Version]
- Agrati, D.; Zuluaga, M.; Fernández-Guasti, A.; Meikle, A.; Ferreira, A. Maternal condition reduces fear behaviors but not the endocrine response to an emotional threat in virgin female rats. Horm. Behav. 2008, 53, 232–240. [Google Scholar] [CrossRef]
- Slattery, D.A.; Neumann, I.D. No stress please! Mechanisms of stress hyporesponsiveness of the maternal brain. J. Physiol. 2008, 586, 377–385. [Google Scholar] [CrossRef]
- Bosch, O.J. Maternal nurturing is dependent on her innate anxiety: The behavioral roles of brain oxytocin and vasopressin. Horm. Behav. 2011, 59, 202–212. [Google Scholar] [CrossRef]
- Bosch, O.; Krömer, S.; Brunton, P.; Neumann, I. Release of oxytocin in the hypothalamic paraventricular nucleus, but not central amygdala or lateral septum in lactating residents and virgin intruders during maternal defence. Neuroscience 2004, 124, 439–448. [Google Scholar] [CrossRef]
- Nephew, B.C.; Bridges, R.S. Central actions of arginine vasopressin and a V1a receptor antagonist on maternal aggression, maternal behavior, and grooming in lactating rats. Pharmacol. Biochem. Behav. 2008, 91, 77–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Numan, M. Maternal behavior: Neural circuits, stimulus valence, and motivational processes. Parenting 2012, 12, 105–114. [Google Scholar] [CrossRef]
- Takayanagi, Y.; Yoshida, M.; Bielsky, I.F.; Ross, H.E.; Kawamata, M.; Onaka, T.; Yanagisawa, T.; Kimura, T.; Matzuk, M.M.; Young, L.J.; et al. Pervasive social deficits, but normal parturition, in oxytocin receptor-deficient mice. Proc. Natl. Acad. Sci. USA 2005, 102, 16096–16101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consiglio, A.R.; Borsoi, A.; Pereira, G.A.; Lucion, A.B. Effects of oxytocin microinjected into the central amygdaloid nucleus and bed nucleus of stria terminalis on maternal aggressive behavior in rats. Physiol. Behav. 2005, 85, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, C.; Vadlamudi, S.; Boccia, M.; Amico, J. Maternal behavior deficits in nulliparous oxytocin knockout mice. Genes Brain Behav. 2006, 5, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Champagne, F.A.; Meaney, M.J. Stress during gestation alters postpartum maternal care and the development of the offspring in a rodent model. Biol. Psychiatry 2006, 59, 1227–1235. [Google Scholar] [CrossRef] [PubMed]
- Lonstein, J.S. Regulation of anxiety during the postpartum period. Front. Neuroendocrinol. 2007, 28, 115–141. [Google Scholar] [CrossRef]
- Bosch, O.J.; Neumann, I.D. Brain vasopressin is an important regulator of maternal behavior independent of dams’ trait anxiety. Proc. Natl. Acad. Sci. USA 2008, 105, 17139–17144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leng, G.; Meddle, S.L.; Douglas, A.J. Oxytocin and the maternal brain. Curr. Opin. Pharmacol. 2008, 8, 731–734. [Google Scholar] [CrossRef] [PubMed]
- Veenema, A.H.; Neumann, I.D. Central vasopressin and oxytocin release: Regulation of complex social behaviours. Prog. Brain Res. 2008, 170, 261–276. [Google Scholar] [PubMed]
- Neumann, I.D. Brain oxytocin: A key regulator of emotional and social behaviours in both females and males. J. Neuroendocrinol. 2008, 20, 858–865. [Google Scholar] [CrossRef]
- Bosch, O.J.; Neumann, I.D. Vasopressin released within the central amygdala promotes maternal aggression. Eur. J. Neurosci. 2010, 31, 883–891. [Google Scholar] [CrossRef]
- Neumann, I.D.; Landgraf, R. Balance of brain oxytocin and vasopressin: Implications for anxiety, depression, and social behaviors. Trends Neurosci. 2012, 35, 649–659. [Google Scholar] [CrossRef]
- Yoshihara, C.; Numan, M.; Kuroda, K.O. Oxytocin and parental behaviors. In Behavioral Pharmacology of Neuropeptides: Oxytocin; Springer: Berlin/Heidelberg, Germany, 2017; pp. 119–153. [Google Scholar]
- Landgraf, R.; Neumann, I.D. Vasopressin and oxytocin release within the brain: A dynamic concept of multiple and variable modes of neuropeptide communication. Front. Neuroendocrinol. 2004, 25, 150–176. [Google Scholar] [CrossRef]
- Beiderbeck, D.I.; Neumann, I.D.; Veenema, A.H. Differences in intermale aggression are accompanied by opposite vasopressin release patterns within the septum in rats bred for low and high anxiety. Eur. J. Neurosci. 2007, 26, 3597–3605. [Google Scholar] [CrossRef]
- Caldwell, H.K.; Lee, H.J.; Macbeth, A.H.; Young III, W.S. Vasopressin: Behavioral roles of an “original” neuropeptide. Prog. Neurobiol. 2008, 84, 1–24. [Google Scholar] [CrossRef] [Green Version]
- McGregor, I.; Callaghan, P.; Hunt, G. From ultrasocial to antisocial: A role for oxytocin in the acute reinforcing effects and long-term adverse consequences of drug use? Br. J. Pharmacol. 2008, 154, 358–368. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.S. Oxytocin pathways and the evolution of human behavior. Annu. Rev. Psychol. 2014, 65, 17–39. [Google Scholar] [CrossRef] [Green Version]
- Knobloch, H.S.; Grinevich, V. Evolution of oxytocin pathways in the brain of vertebrates. Front. Behav. Neurosci. 2014, 8, 31. [Google Scholar] [CrossRef]
- Febo, M.; Numan, M.; Ferris, C.F. Functional magnetic resonance imaging shows oxytocin activates brain regions associated with mother–pup bonding during suckling. J. Neurosci. 2005, 25, 11637–11644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Febo, M.; Stolberg, T.L.; Numan, M.; Bridges, R.S.; Kulkarni, P.; Ferris, C.F. Nursing stimulation is more than tactile sensation: It is a multisensory experience. Horm. Behav. 2008, 54, 330–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Febo, M. A bold view of the lactating brain: Functional magnetic resonance imaging studies of suckling in awake dams. J. Neuroendocrinol. 2011, 23, 1009–1019. [Google Scholar] [CrossRef] [PubMed]
- Dobolyi, A.; Grattan, D.; Stolzenberg, D. Preoptic inputs and mechanisms that regulate maternal responsiveness. J. Neuroendocrinol. 2014, 26, 627–640. [Google Scholar] [CrossRef] [Green Version]
- Gammie, S.C.; Hasen, N.S.; Awad, T.A.; Auger, A.P.; Jessen, H.M.; Panksepp, J.B.; Bronikowski, A.M. Gene array profiling of large hypothalamic CNS regions in lactating and randomly cycling virgin mice. Mol. Brain Res. 2005, 139, 201–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolzenberg, D.S.; Stevens, J.S.; Rissman, E.F. Experience-facilitated improvements in pup retrieval; evidence for an epigenetic effect. Horm. Behav. 2012, 62, 128–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbari, E.M.; Shams, S.; Belay, H.T.; Kaiguo, M.; Razak, Z.; Kent, C.F.; Westwood, T.; Sokolowski, M.B.; Fleming, A.S. The effects of parity and maternal behavior on gene expression in the medial preoptic area and the medial amygdala in postpartum and virgin female rats: A microarray study. Behav. Neurosci. 2013, 127, 913. [Google Scholar] [CrossRef]
- Roozendaal, B.; Hernandez, A.; Cabrera, S.M.; Hagewoud, R.; Malvaez, M.; Stefanko, D.P.; Haettig, J.; Wood, M.A. Membrane-associated glucocorticoid activity is necessary for modulation of long-term memory via chromatin modification. J. Neurosci. 2010, 30, 5037–5046. [Google Scholar] [CrossRef] [Green Version]
- Malvaez, M.; Sanchis-Segura, C.; Vo, D.; Lattal, K.M.; Wood, M.A. Modulation of chromatin modification facilitates extinction of cocaine-induced conditioned place preference. Biol. Psychiatry 2010, 67, 36–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wynne-Edwards, K.E.; Timonin, M.E. Paternal care in rodents: Weakening support for hormonal regulation of the transition to behavioral fatherhood in rodent animal models of biparental care. Horm. Behav. 2007, 52, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Kenkel, W.; Paredes, J.; Yee, J.; Pournajafi-Nazarloo, H.; Bales, K.; Carter, C. Neuroendocrine and behavioural responses to exposure to an infant in male prairie voles. J. Neuroendocrinol. 2012, 24, 874–886. [Google Scholar] [CrossRef]
- De Jong, T.R.; Chauke, M.; Harris, B.N.; Saltzman, W. From here to paternity: Neural correlates of the onset of paternal behavior in California mice (Peromyscus californicus). Horm. Behav. 2009, 56, 220–231. [Google Scholar] [CrossRef]
- De Jong, T.; Measor, K.; Chauke, M.; Harris, B.; Saltzman, W. Brief pup exposure induces Fos expression in the lateral habenula and serotonergic caudal dorsal raphe nucleus of paternally experienced male California mice (Peromyscus californicus). Neuroscience 2010, 169, 1094–1104. [Google Scholar] [CrossRef]
- Lambert, K.G.; Franssen, C.L.; Bardi, M.; Hampton, J.E.; Hainley, L.; Karsner, S.; Tu, E.B.; Hyer, M.M.; Crockett, A.; Baranova, A.; et al. Characteristic neurobiological patterns differentiate paternal responsiveness in two Peromyscus species. Brain Behav. Evol. 2011, 77, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Tachikawa, K.S.; Yoshihara, Y.; Kuroda, K.O. Behavioral transition from attack to parenting in male mice: A crucial role of the vomeronasal system. J. Neurosci. 2013, 33, 5120–5126. [Google Scholar] [CrossRef]
- Isogai, Y.; Wu, Z.; Love, M.I.; Ahn, M.H.Y.; Bambah-Mukku, D.; Hua, V.; Farrell, K.; Dulac, C. Multisensory logic of infant-directed aggression by males. Cell 2018, 175, 1827–1841. [Google Scholar] [CrossRef] [Green Version]
- Tsuneoka, Y.; Yoshida, S.; Takase, K.; Oda, S.; Kuroda, M.; Funato, H. Neurotransmitters and neuropeptides in gonadal steroid receptor-expressing cells in medial preoptic area subregions of the male mouse. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Numan, M.; Young, L.J. Neural mechanisms of mother–infant bonding and pair bonding: Similarities, differences, and broader implications. Horm. Behav. 2016, 77, 98–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Numan, M. Parental Behavior. In Reference Module in Neuroscience and Biobehavioral Psychology; Elsevier: London, UK, 2017. [Google Scholar]
- Chen, C. CiteSpace II: Detecting and visualizing emerging trends and transient patterns in scientific literature. J. Am. Soc. Inf. Sci. Technol. 2006, 57, 359–377. [Google Scholar] [CrossRef] [Green Version]
- Haddaway, N.R.; Collins, A.M.; Coughlin, D.; Kirk, S. The role of Google Scholar in evidence reviews and its applicability to grey literature searching. PLoS ONE 2015, 10, e0138237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Soeken, T.A.; Cromer, S.J.; Martinez, S.R.; Hardy, L.R.; Strathearn, L. Oxytocin and postpartum depression: Delivering on what’s known and what’s not. Brain Res. 2014, 1580, 219–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Cluster ID | Size | Silhouette | Mean Year |
|---|---|---|---|
| 0 | 190 | 0.879 | 2010 |
| 1 | 129 | 0.885 | 2002 |
| 2 | 115 | 0.902 | 1994 |
| 4 | 69 | 0.915 | 1990 |
| 6 | 50 | 0.968 | 2014 |
| 7 | 41 | 0.966 | 2001 |
| 8 | 39 | 0.983 | 2004 |
| 9 | 36 | 0.99 | 2014 |
| 10 | 29 | 0.989 | 1987 |
| 11 | 27 | 0.983 | 1987 |
| Reference | Strenght of Burstness | Year | Beginning of Burstness | End of Burstness | Burst Duration | Sigma | Centrality |
|---|---|---|---|---|---|---|---|
| Numan and Insel [16] | 18.61 | 2003 | 2005 | 2011 | 6 | 3.02 | 0.06 |
| Wu et al. [19] | 16.05 | 2014 | 2015 | 2020 | 5 | 1.84 | 0.04 |
| Numan and Stolzenberg [33] | 14.55 | 2009 | 2010 | 2017 | 7 | 2.29 | 0.06 |
| Tsuneoka et al. [36] | 13.67 | 2013 | 2014 | 2020 | 6 | 1.49 | 0.03 |
| Numan [37] | 10.95 | 2006 | 2007 | 2013 | 6 | 1.71 | 0.05 |
| Numan and Numan [38] | 10.24 | 1995 | 1997 | 2002 | 5 | 1.16 | 0.01 |
| Dulac et al. [39] | 9.58 | 2014 | 2015 | 2020 | 5 | 1.19 | 0.02 |
| Shahrokh et al. [40] | 8.94 | 2010 | 2011 | 2017 | 6 | 1.48 | 0.04 |
| Pereira and Morrell [41] | 8.90 | 2011 | 2013 | 2020 | 7 | 1.29 | 0.03 |
| Numan et al. [34] | 8.26 | 2005 | 2005 | 2013 | 8 | 1.58 | 0.06 |
| Bridges [42] | 7.39 | 2015 | 2016 | 2020 | 4 | 1.00 | 0.00 |
| Stack et al. [43] | 7.20 | 2002 | 2005 | 2010 | 5 | 1.28 | 0.04 |
| Bosch and Neumann [44] | 7.19 | 2012 | 2013 | 2020 | 7 | 1.11 | 0.01 |
| Champagne et al. [35] | 7.09 | 2004 | 2005 | 2012 | 7 | 1.91 | 0.10 |
| Brown et al. [45] | 6.99 | 1996 | 1997 | 2002 | 5 | 1.41 | 0.05 |
| Keer and Stern [46] | 6.59 | 1999 | 2004 | 2007 | 3 | 1.18 | 0.03 |
| Numan and Numan [47] | 6.30 | 1994 | 1998 | 2002 | 4 | 1.21 | 0.03 |
| Numan [48] | 5.92 | 2007 | 2008 | 2015 | 7 | 1.05 | 0.01 |
| Meddle et al. [49] | 5.83 | 2007 | 2010 | 2014 | 4 | 1.01 | 0.00 |
| Tsuneoks et al. [50] | 5.82 | 2015 | 2017 | 2020 | 3 | 1.01 | 0.00 |
| Reference | Strength of Burstness | Beginning of Burstness | End of Burstness | Burst Duration |
|---|---|---|---|---|
| support | 49.07 | 1979 | 1995 | 16 |
| metabolism | 42.12 | 2015 | 2020 | 5 |
| physiology | 33.13 | 2015 | 2020 | 5 |
| central nervous system | 28.94 | 1978 | 1987 | 9 |
| hypothalamus | 24.34 | 1972 | 1993 | 21 |
| medial preoptic area | 14.02 | 2010 | 2020 | 10 |
| maternal behavior | 11.07 | 2016 | 2018 | 2 |
| pregnancy | 10.05 | 1978 | 1985 | 7 |
| male | 8.59 | 2018 | 2020 | 2 |
| protein expression | 7.90 | 2002 | 2010 | 8 |
| preoptic area | 7.82 | 2004 | 2008 | 4 |
| rat | 7.15 | 1977 | 1999 | 22 |
| estradiol | 6.55 | 1987 | 1989 | 2 |
| radioisotope | 6.54 | 1984 | 1990 | 6 |
| animal tissue | 5.57 | 2009 | 2011 | 2 |
| endocrine system | 5.37 | 1982 | 1984 | 2 |
| ovariectomy | 5.24 | 1987 | 1989 | 2 |
| theoretical study | 4.82 | 1974 | 1977 | 3 |
| histology | 4.67 | 1980 | 1982 | 2 |
| aging | 4.55 | 1978 | 1989 | 12 |
| brain | 4.24 | 1987 | 1988 | 1 |
| animal cell | 3.92 | 1987 | 1989 | 2 |
| Cluster | Citing Document | GCS | Coverage |
|---|---|---|---|
| 4 | Nagano and Shinoda [80] | 20 | 9 |
| 4 | Bridges et al. [56] | 78 | 9 |
| 4 | Fleming and Walsh [53] | 79 | 9 |
| 4 | Fleming and Korsmit [54] | 108 | 8 |
| 4 | Dellovade et al. [79] | 32 | 7 |
| 4 | Rizvi et al. [81] | 135 | 7 |
| 4 | Rosenblatt et al. [70] | 75 | 6 |
| 4 | Bridges and Freemark [55] | 34 | 6 |
| 4 | Ehret and Buckenmaier [69] | 36 | 5 |
| Cluster | Citing Document | GCS | Coverage |
|---|---|---|---|
| 2 | Kalinichev et al. [102] | 54 | 21 |
| 2 | Stack et al. [43] | 103 | 15 |
| 2 | Lonstein et al. [114] | 72 | 14 |
| 2 | Komisaruk et al. [113] | 31 | 13 |
| 2 | Sheehan and Numan [115] | 48 | 13 |
| 2 | Stack and Numan [107] | 56 | 13 |
| 2 | Grattan [119] | 114 | 12 |
| 2 | Lin et al. [108] | 29 | 12 |
| 2 | Li et al. [101] | 88 | 12 |
| 2 | Lonstein and De Vries [110] | 64 | 11 |
| Cluster | Citing Document | GCS | Coverage |
|---|---|---|---|
| 1 | Gammie [120] | 69 | 25 |
| 1 | Curtis et al. [176] | 57 | 19 |
| 1 | Numan [37] | 159 | 17 |
| 1 | Numan and Stolzenberg [33] | 224 | 17 |
| 1 | Numan et al. [128] | 119 | 15 |
| 1 | Numan and Woodside [174] | 89 | 15 |
| 1 | Pereira and Morrell [41] | 84 | 14 |
| 1 | Perrin et al. [177] | 37 | 14 |
| 1 | Numan et al. [34] | 91 | 14 |
| 1 | Olazabal and Young [122] | 176 | 12 |
| Cluster | Citing Document | GCS | Coverage |
|---|---|---|---|
| 0 | Lonstein et al. [192] | 62 | 31 |
| 0 | Numan and Young [250] | 131 | 23 |
| 0 | Mchenry et al. [193] | 20 | 20 |
| 0 | Olazabal et al. [189] | 55 | 18 |
| 0 | Dobolyi et al. [236] | 40 | 18 |
| 0 | Yoshihara et al. [226] | 15 | 18 |
| 0 | Rutherford et al. [178] | 81 | 17 |
| 0 | Bosch [211] | 100 | 16 |
| 0 | Bridges [42] | 146 | 16 |
| 0 | Numan [251] | 0 | 16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carollo, A.; Balagtas, J.P.M.; Neoh, M.J.-Y.; Esposito, G. A Scientometric Approach to Review the Role of the Medial Preoptic Area (MPOA) in Parental Behavior. Brain Sci. 2021, 11, 393. https://doi.org/10.3390/brainsci11030393
Carollo A, Balagtas JPM, Neoh MJ-Y, Esposito G. A Scientometric Approach to Review the Role of the Medial Preoptic Area (MPOA) in Parental Behavior. Brain Sciences. 2021; 11(3):393. https://doi.org/10.3390/brainsci11030393
Chicago/Turabian StyleCarollo, Alessandro, Jan Paolo Macapinlac Balagtas, Michelle Jin-Yee Neoh, and Gianluca Esposito. 2021. "A Scientometric Approach to Review the Role of the Medial Preoptic Area (MPOA) in Parental Behavior" Brain Sciences 11, no. 3: 393. https://doi.org/10.3390/brainsci11030393
APA StyleCarollo, A., Balagtas, J. P. M., Neoh, M. J.-Y., & Esposito, G. (2021). A Scientometric Approach to Review the Role of the Medial Preoptic Area (MPOA) in Parental Behavior. Brain Sciences, 11(3), 393. https://doi.org/10.3390/brainsci11030393