
The Effect of Vasopressin Antagonists on Maternal-Separation-Induced Ultrasonic Vocalization and Stress-Hormone Level Increase during the Early Postnatal Period


Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Measurements
2.2.1. Maternal Separation-Induced Ultrasonic Vocalization
2.2.2. Testing Sedative Side Effects
2.2.3. Hormone Measurements
2.3. Experiments
2.3.1. Experiment 1: Genetic AVP Deficiency
2.3.2. Experiment 2: Pharmacological AVP-Effect Deficiency
2.4. Statistical Analysis
3. Results
3.1. Genetic AVP Deficiency
3.2. Pharmacological AVP Deficiency
3.2.1. V1aR Antagonist
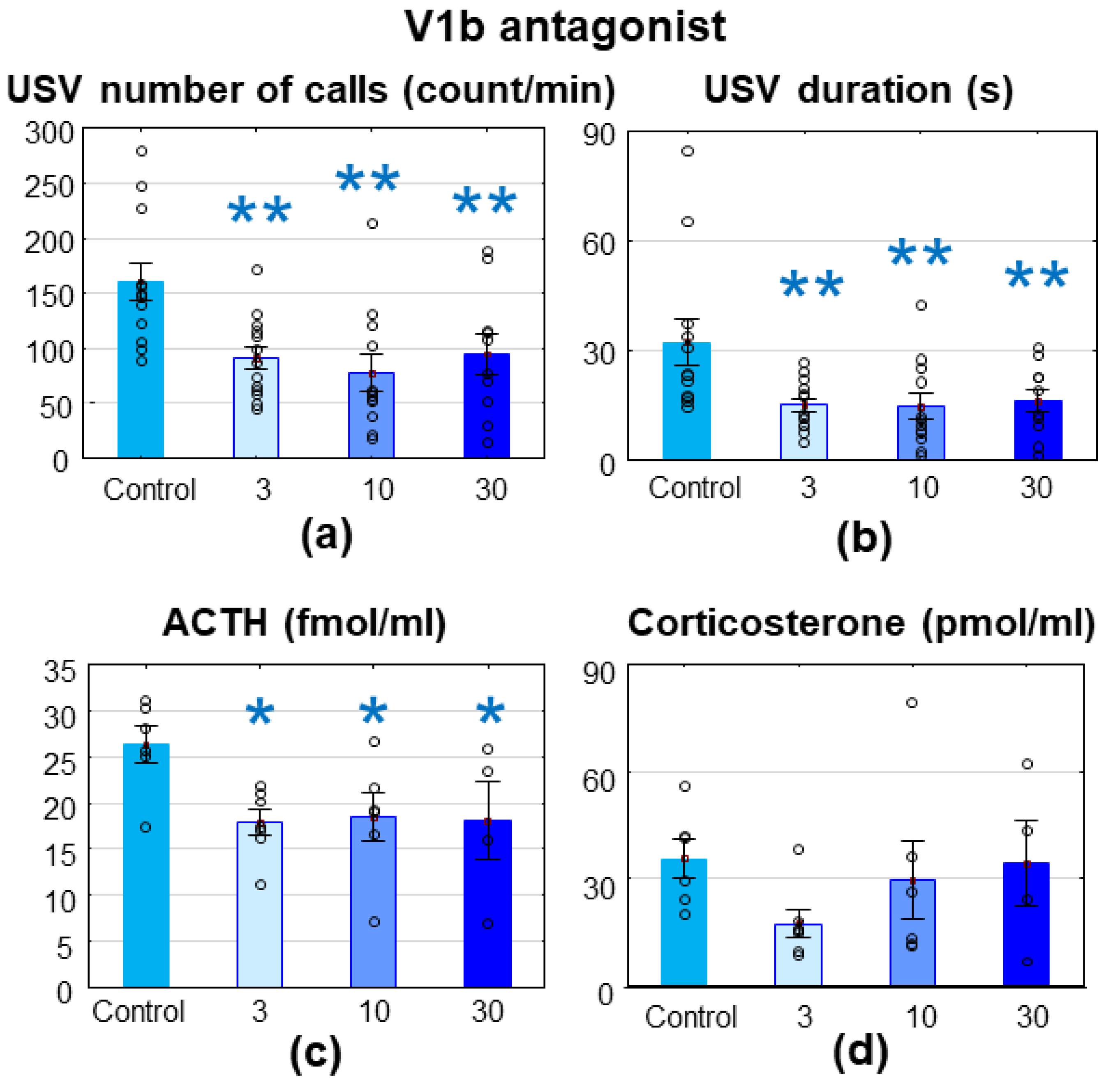
3.2.2. V1bR Antagonist
3.2.3. V2R Antagonist
3.2.4. V1aR + V1bR Antagonists
3.3. Correlations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frank, E.; Landgraf, R. The vasopressin system–from antidiuresis to psychopathology. Eur. J. Pharmacol. 2008, 583, 226–542. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, D. Vasopressin and oxytocin: Hypothalamic modulators of the stress response: A review. Psychoneuroendocrinology 1986, 11, 131–139. [Google Scholar] [CrossRef]
- Keck, M. Corticotropin-releasing factor, vasopressin and receptor systems in depression and anxiety. Amino Acids 2006, 31, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Hofer, M.A. Multiple regulators of ultrasonic vocalization in the infant rat. Psychoneuroendocrinology 1996, 21, 203–217. [Google Scholar] [CrossRef]
- Zelena, D. The role of vasopressin in anxiety: What can we learn from studying ultrasonic vocalization? In Handbook of Behavioral Neuroscience, A Window into the Emotional Brain, 1st ed.; Brudzynski, S.M., Ed.; Elsevier/Academic Press: London, UK, 2018; Chapter 41; pp. 433–441. [Google Scholar]
- Varga, J.; Fodor, A.; Klausz, B.; Zelena, D. Anxiogenic role of vasopressin during the early postnatal period: Maternal separation-induced ultrasound vocalization in vasopressin-deficient Brattleboro rats. Amino Acids 2015, 47, 2409–2418. [Google Scholar] [CrossRef] [PubMed]
- Mlynarik, M.; Zelena, D.; Bagdy, G.; Makara, G.B.; Jezova, D. Signs of attenuated depression-like behavior in vasopressin deficient Brattleboro rats. Horm. Behav. 2007, 51, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Koshimizu, T.; Nakamura, K.; Egashira, N.M.H.; Nonoguchi, H.; Tanoue, A. Vasopressin V1a and V1b receptors: From molecules to physiological systems. Physiol. Rev. 2012, 92, 1813–1864. [Google Scholar] [CrossRef] [Green Version]
- Shimazaki, T.; Iijima, M.; Chaki, S. The pituitary mediates the anxiolytic-like effects of the vasopressin V1B receptor antagonist, SSR149415, in a social interaction test in rats. Eur. J. Pharmacol. 2006, 543, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Bayerl, D.; Hönig, J.; Bosch, O. Vasopressin V1a, but not V1b, receptors within the PVN of lactating rats mediate maternal care and anxiety-related behaviour. Behav. Brain Res. 2016, 305, 318–322. [Google Scholar] [CrossRef]
- Griebel, G.; Beeské, S.; Stahl, S. The vasopressin V(1b) receptor antagonist SSR149415 in the treatment of major depressive and generalized anxiety disorders: Results from 4 randomized, double-blind, placebo-controlled studies. J. Clin. Psychiatry 2012, 73, 1403–1411. [Google Scholar] [CrossRef]
- Zelena, D. Vasopressin in health and disease with a focus on affective disorders. Cent. Nerv. Syst. Agents Med. Chem. 2012, 12, 286–303. [Google Scholar] [CrossRef] [PubMed]
- Le Gal, C.S.; Wagnon, J.; Simiand, J.; Griebel, G.; Lacour, C.; Guillon, G.; Barberis, C.; Brossard, G.; Soubrie, P.; Nisato, D.; et al. Characterization of (2S,4R)-1-[5-chloro-1-[(2,4-dimethoxyphenyl)sulfonyl]-3-(2-methoxy-phenyl)-2-oxo- 2,3-dihydro-1H-indol-3-yl]-4-hydroxy-N,N-dimethyl-2-pyrrolidine carboxamide (SSR149415), a selective and orally active vasopressin V1b receptor antagonist. J. Pharmacol. Exp. Ther. 2002, 300, 1122–1130. [Google Scholar] [CrossRef]
- Williams, D.R.; Carlsson, R.; Burkner, P.C. Between-litter variation in developmental studies of hormones and behavior: Inflated false positives and diminished power. Front. Neuroendocrinol. 2017, 47, 154–166. [Google Scholar] [CrossRef] [Green Version]
- Blumberg, M.S.; Sokoloff, G.; Kent, K.J. A developmental analysis of clonidine’s effects on cardiac rate and ultrasound production in infant rats. Dev. Psychobiol. 2000, 36, 186–193. [Google Scholar] [CrossRef]
- Allin, J.T.; Banks, E.M. Effects of temperature on ultrasound production by infant albino rats. Dev. Psychobiol. 1971, 4, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Zelena, D.; Mergl, Z.; Makara, G.B. Postnatal development in vasopressin deficient Brattleboro rats with special attention to the hypothalamo-pituitary-adrenal axis function: The role of maternal genotype. Int. J. Dev. Neurosci. 2009, 27, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Zelena, D.; Kiem, D.T.; Barna, I.; Makara, G.B. Alpha 2-adrenoreceptor subtypes regulate ACTH and beta-endorphin secretions during stress in the rat. Psychoneuroendocrinology 1999, 24, 333–343. [Google Scholar] [CrossRef]
- Zelena, D.; Mergl, Z.; Foldes, A.; Kovacs, K.J.; Toth, Z.; Makara, G.B. Role of hypothalamic inputs in maintaining pituitary-adrenal responsiveness in repeated restraint. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E1110–E1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelena, D.; Mergl, Z.; Makara, G.B. Maternal genotype influences stress reactivity of vasopressin-deficient brattleboro rats. J Neuroendocrinol. 2003, 15, 1105–1110. [Google Scholar] [CrossRef] [PubMed]
- Serradeil-Le Gal, C.; Wagnon, J.; Garcia, C.; Lacour, C.; Guiraudou, P.; Christophe, B.; Villanova, G.; Nisato, D.; Maffrand, J.P.; Le Fur, G.; et al. Biochemical and pharmacological properties of SR 49059, a new, potent, nonpeptide antagonist of rat and human vasopressin V1a receptors. J. Clin. Investig. 1993, 92, 224–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Gal, C.S.; Raufaste, D.; Double-Cazanave, E.; Guillon, G.; Garcia, C.; Pascal, M.; Maffrand, J.P. Binding properties of a selective tritiated vasopressin V2 receptor antagonist, [H]-SR 121463. Kidney Int. 2000, 58, 1613–1622. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, W.; Gal, C.S.; Ros, J.; Cano, C.; Cejudo, P.; Morales-Ruiz, M.; Arroyo, V.; Pascal, M.; Rivera, F.; Maffrand, J.P.; et al. Long-term aquaretic efficacy of a selective nonpeptide V(2)-vasopressin receptor antagonist, SR121463, in cirrhotic rats. J. Pharmacol. Exp. Ther. 2000, 295, 83–90. [Google Scholar] [PubMed]
- Hodgson, R.A.; Higgins, G.A.; Guthrie, D.H.; Lu, S.X.; Pond, A.J.; Mullins, D.E.; Guzzi, M.F.; Parker, E.M.; Varty, G.B. Comparison of the V1b antagonist, SSR149415, and the CRF1 antagonist, CP-154,526, in rodent models of anxiety and depression. Pharmacol. Biochem. Behav. 2007, 86, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Griebel, G.; Simiand, J.; Serradeil-Le Gal, C.; Wagnon, J.; Pascal, M.; Scatton, B.; Maffrand, J.P.; Soubrie, P. Anxiolytic- and antidepressant-like effects of the non-peptide vasopressin V1b receptor antagonist, SSR149415, suggest an innovative approach for the treatment of stress-related disorders. Proc. Natl. Acad. Sci. USA 2002, 99, 6370–6375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iijima, M.; Chaki, S. Separation-induced ultrasonic vocalization in rat pups: Further pharmacological characterization. Pharmacol. Biochem. Behav. 2005, 82, 652–657. [Google Scholar] [CrossRef]
- Iijima, M.; Yoshimizu, T.; Shimazaki, T.; Tokugawa, K.; Fukumoto, K.; Kurosu, S.; Kuwada, T.; Sekiguchi, Y.; Chaki, S. Antidepressant and anxiolytic profiles of newly synthesized arginine vasopressin V1B receptor antagonists: TASP0233278 and TASP0390325. Br. J. Pharmacol. 2014, 171, 3511–3525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scattoni, M.L.; McFarlane, H.G.; Zhodzishsky, V.; Caldwell, H.K.; Young, W.S.; Ricceri, L.; Crawley, J.N. Reduced ultrasonic vocalizations in vasopressin 1b knockout mice. Behav. Brain Res. 2008, 187, 371–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelena, D.; Makara, G.B. The role of adrenocorticotropin beyond the glucocorticoid horizon. In Advances in Medicine and Biology; Nova Publisher: Hauppauge, NY, USA, 2012; Volume 43, pp. 109–138. [Google Scholar]
- Kitamura, Y.; Gomita, Y. Development of animal models of treatment-resistant depression in rats. Nihon Shinkei Seishin Yakurigaku Zasshi 2008, 28, 93–100. [Google Scholar]
- McCaughey, E.S.; Walker, V.; Rolles, C.J.; Scheurmier, N.I.; Hale, A.C.; Rees, L.H. Ectopic ACTH production by a thymic carcinoid tumour. Eur. J. Pediatr. 1987, 146, 590–591. [Google Scholar] [CrossRef]
- Kokras, N.; Sotiropoulos, I.; Pitychoutis, P.M.; Almeida, O.F.; Papadopoulou-Daifoti, Z. Citalopram-mediated anxiolysis and differing neurobiological responses in both sexes of a genetic model of depression. Neuroscience 2011, 194, 62–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebner, K.; Wotjak, C.; Holsboer, F.; Landgraf, R.; Engelmann, M. Vasopressin released within the septal brain area during swim stress modulates the behavioural stress response in rats. Eur. J. Neurosci. 1999, 11, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Egashira, N.; Tanoue, A.; Matsuda, T.; Koushi, E.; Harada, S.; Takano, Y.; Tsujimoto, G.; Mishima, K.; Iwasaki, K.; Fujiwara, M. Impaired social interaction and reduced anxiety-related behavior in vasopressin V1a receptor knockout mice. Behav. Brain Res. 2007, 178, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Bielsky, I.; Hu, S.; Ren, X.; Terwilliger, E.; Young, L. The V1a vasopressin receptor is necessary and sufficient for normal social recognition: A gene replacement study. Neuron 2005, 47, 503–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bleickardt, C.J.; Mullins, D.E.; Macsweeney, C.P.; Werner, B.J.; Pond, A.J.; Guzzi, M.F.; Martin, F.D.; Varty, G.B.; Hodgson, R.A. Characterization of the V1a antagonist, JNJ-17308616, in rodent models of anxiety-like behavior. Psychopharmacology 2009, 202, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Koshimizu, T.A.; Nasa, Y.; Tanoue, A.; Oikawa, R.; Kawahara, Y.; Kiyono, Y.; Adachi, T.; Tanaka, T.; Kuwaki, T.; Mori, T.; et al. V1a vasopressin receptors maintain normal blood pressure by regulating circulating blood volume and baroreflex sensitivity. Proc. Natl. Acad. Sci. USA 2006, 103, 7807–7812. [Google Scholar] [CrossRef] [Green Version]
- Tribollet, E.; Raufaste, D.; Maffrand, J.; Serradeil-Le Gal, C. Binding of the non-peptide vasopressin V1a receptor antagonist SR-49059 in the rat brain: An in vitro and in vivo autoradiographic study. Neuroendocrinology 1999, 69, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, G.; Parrott, N.; Prinssen, E.; Barrow, P. The great barrier belief: The blood-brain barrier and considerations for juvenile toxicity studies. Reprod. Toxicol. 2017, 72, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Winslow, J.T.; Insel, T.R. Effects of central vasopressin administration to infant rats. Eur. J. Pharmacol. 1993, 233, 101–107. [Google Scholar] [CrossRef]
- Altstein, M.; Gainer, H. Differential biosynthesis and posttranslational processing of vasopressin and oxytocin in rat brain during embryonic and postnatal development. J. Neurosci. 1988, 8, 3967–3977. [Google Scholar] [CrossRef] [Green Version]
- Tribollet, E.; Goumaz, M.; Raggenbass, M.; Dubois-Dauphin, M.; Dreifuss, J.J. Early appearance and transient expression of vasopressin receptors in the brain of rat fetus and infant. An autoradiographical and electrophysiological study. Brain Res. Dev. Brain Res. 1991, 58, 13–24. [Google Scholar] [CrossRef]
- Kato, Y.; Igarashi, N.; Hirasawa, A.; Tsujimoto, G.; Kobayashi, M. Distribution and developmental changes in vasopressin V2 receptor mRNA in rat brain. Differentiation 1995, 59, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Ammar, A.; Roseau, S.; Butlen, D. Postnatal ontogenesis of vasopressin receptors in the rat collecting duct. Mol. Cell Endocrinol. 1992, 86, 193–203. [Google Scholar] [CrossRef]
- Varga, J.; Ferenczi, S.; Kovacs, K.J.; Garafova, A.; Jezova, D.; Zelena, D. Comparison of stress-induced changes in adults and pups: Is aldosterone the main adrenocortical stress hormone during the perinatal period in rats? PLoS ONE 2013, 8, e72313. [Google Scholar] [CrossRef] [Green Version]
- Jurcovicova, J.; Vigas, M.; Klir, P.; Jezova, D. Response of prolactin, growth hormone and corticosterone secretion to morphine administration or stress exposure in Wistar-AVN and Long Evans rats. Endocrinol. Exp. 1984, 18, 209–214. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antagonist Treatment | Time (s) | Doses (mg/kg) | |||
|---|---|---|---|---|---|
| 0 | 3 | 10 | 30 | ||
| V1aR | Righting | 2.333 ± 0.343 | 2.361 ± 0.230 | 2.758 ± 0.397 | 2.212 ± 0.187 |
| Neg.geo. | 19.861 ± 2.687 | 20.750 ± 2.984 | 18.212 ± 2.735 | 16.485 ± 2.929 | |
| V1bR | Righting | 1.303 ± 0.156 | 2.279 ± 0.669 | 1.947 ± 0.303 | 2.103 ± 0.266 |
| Neg.geo. | 9.194 ± 0.963 | 8.487 ± 0.880 | 9.528 ± 1.215 | 10.100 ± 1.673 | |
| V2R | Righting | 2.133 ± 0.218 | 2.267 ± 0.325 | 2.033 ± 0.195 | 2.250 ± 0.300 |
| Neg.geo. | 11.967 ± 1.961 | 13.970 ± 2.707 | 15.267 ± 3.010 | 18.667 ± 2.567 | |
| V1aR + V1bR | Righting | 1.741 ± 0.282 | 2.750 ± 0.552 | ||
| Neg.geo. | 8.296 ± 1.301 | 7.125 ± 0.900 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Török, B.; Fodor, A.; Zsebők, S.; Sipos, E.; Zelena, D. The Effect of Vasopressin Antagonists on Maternal-Separation-Induced Ultrasonic Vocalization and Stress-Hormone Level Increase during the Early Postnatal Period. Brain Sci. 2021, 11, 444. https://doi.org/10.3390/brainsci11040444
Török B, Fodor A, Zsebők S, Sipos E, Zelena D. The Effect of Vasopressin Antagonists on Maternal-Separation-Induced Ultrasonic Vocalization and Stress-Hormone Level Increase during the Early Postnatal Period. Brain Sciences. 2021; 11(4):444. https://doi.org/10.3390/brainsci11040444
Chicago/Turabian StyleTörök, Bibiána, Anna Fodor, Sándor Zsebők, Eszter Sipos, and Dóra Zelena. 2021. "The Effect of Vasopressin Antagonists on Maternal-Separation-Induced Ultrasonic Vocalization and Stress-Hormone Level Increase during the Early Postnatal Period" Brain Sciences 11, no. 4: 444. https://doi.org/10.3390/brainsci11040444
APA StyleTörök, B., Fodor, A., Zsebők, S., Sipos, E., & Zelena, D. (2021). The Effect of Vasopressin Antagonists on Maternal-Separation-Induced Ultrasonic Vocalization and Stress-Hormone Level Increase during the Early Postnatal Period. Brain Sciences, 11(4), 444. https://doi.org/10.3390/brainsci11040444