
Extracellular Dopamine Levels in Nucleus Accumbens after Chronic Stress in Rats with Persistently High vs. Low 50-kHz Ultrasonic Vocalization Response


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Procedure
2.2. Tickling
2.3. Chronic Variable Stress Regimen (CVS)
2.4. Microdialysis Procedure
2.5. Quantification of Dopamine in Microdialysates
2.6. Data Analysis
3. Results
3.1. Stress Effect on Body Weight Gain
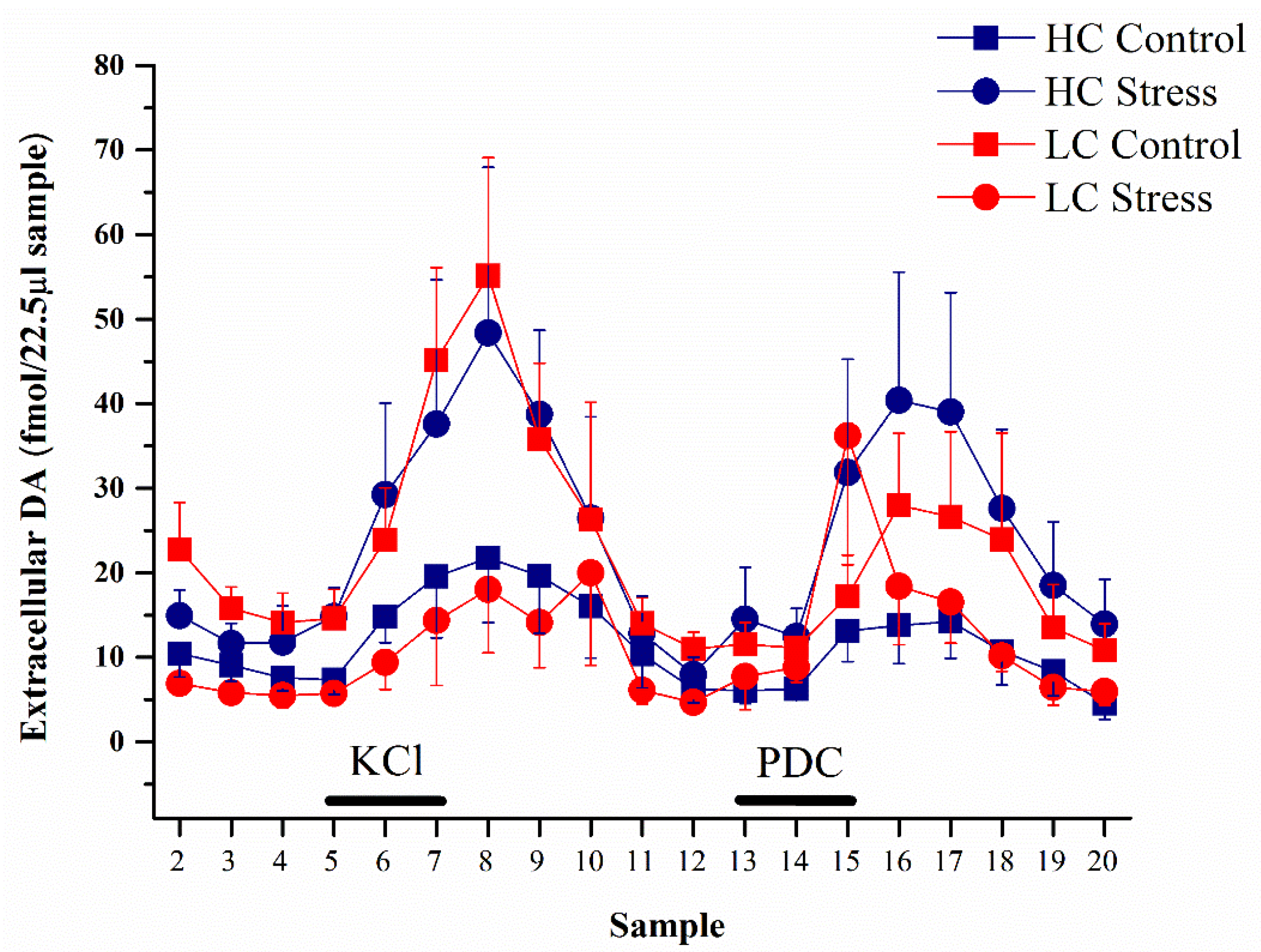
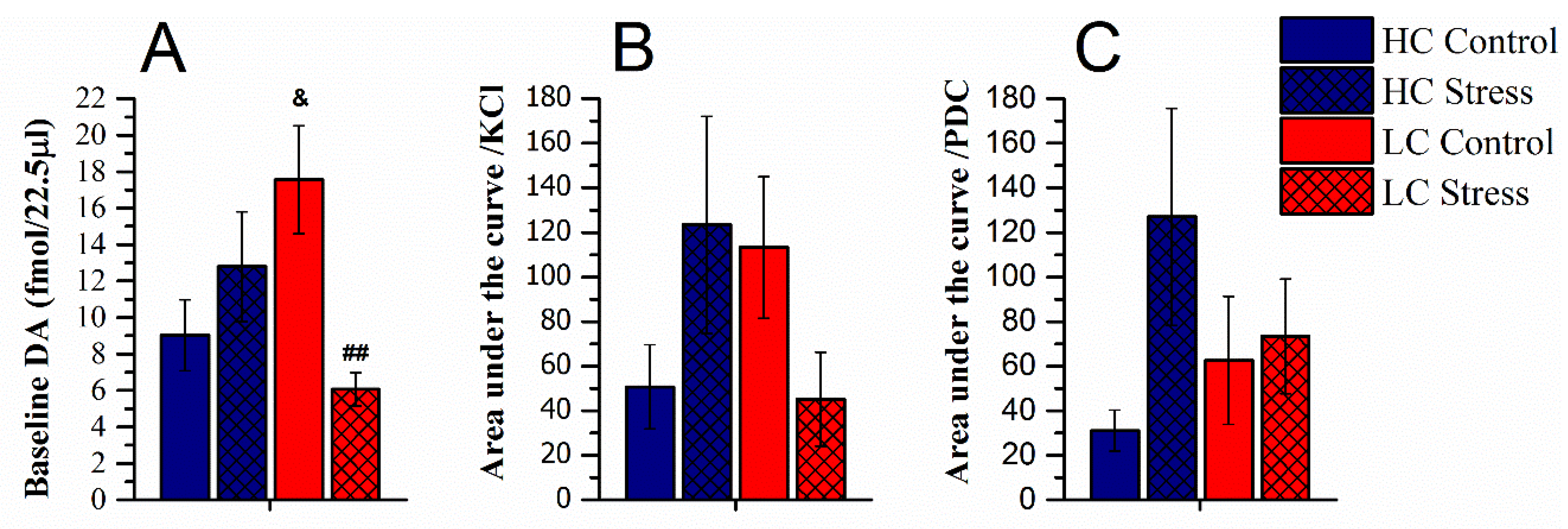
3.2. Dopamine Content in Microdialysates
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Brudzynski, S.M. Ethotransmission: Communication of emotional states through ultrasonic vocalization in rats. Curr. Opin. Neurobiol. 2013, 23, 310–317. [Google Scholar] [CrossRef]
- Wöhr, M.; Schwarting, R.K.W. Affective communication in rodents: Ultrasonic vocalizations as a tool for research on emotion and motivation. Cell Tissue Res. 2013, 354, 81–97. [Google Scholar] [CrossRef]
- Wöhr, M. Ultrasonic communication in rats: Appetitive 50-kHz ultrasonic vocalizations as social contact calls. Behav. Ecol. Sociobiol. 2018, 72, 1–12. [Google Scholar] [CrossRef]
- Panksepp, J.; Burgdorf, J. 50-kHz chirping (laughter?) in response to conditioned and unconditioned tickle-induced reward in rats: Effects of social housing and genetic variables. Behav. Brain Res. 2000, 115, 25–38. [Google Scholar] [CrossRef]
- Burgdorf, J.; Panksepp, J. Tickling induces reward in adolescent rats. Physiol. Behav. 2001, 72, 167–173. [Google Scholar] [CrossRef]
- Burgdorf, J.; Wood, P.L.; Kroes, R.A.; Moskal, J.R.; Panksepp, J. Neurobiology of 50-kHz ultrasonic vocalizations in rats: Electrode mapping, lesion, and pharmacology studies. Behav. Brain Res. 2007, 182, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Grant, L.M.; Barth, K.J.; Muslu, C.; Kelm-Nelson, C.A.; Bakshi, V.P.; Ciucci, M.R. Noradrenergic receptor modulation influences the acoustic parameters of pro-social rat ultrasonic vocalizations. Behav. Neurosci. 2018, 132, 269–283. [Google Scholar] [CrossRef]
- Wöhr, M.; van Gaalen, M.M.; Schwarting, R.K.W. Affective communication in rodents. Behav. Pharmacol. 2015, 26, 506–521. [Google Scholar] [CrossRef] [PubMed]
- Wöhr, M.; Schwarting, R.K.W. Ultrasonic communication in rats: Effects of morphine and naloxone on vocal and behavioral responses to playback of 50-kHz vocalizations. Pharmacol. Biochem. Behav. 2009, 94, 285–295. [Google Scholar] [CrossRef]
- Simola, N.; Costa, G.; Morelli, M. Activation of adenosine A2A receptors suppresses the emission of pro-social and drug-stimulated 50-kHz ultrasonic vocalizations in rats: Possible relevance to reward and motivation. Psychopharmacology 2016, 233, 507–519. [Google Scholar] [CrossRef]
- Fu, X.W.; Brudzynski, S.M. High-frequency ultrasonic vocalization induced by intracerebral glutamate in rats. Pharmacol. Biochem. Behav. 1994, 49, 835–841. [Google Scholar] [CrossRef]
- Harro, J. Animal models of depression vulnerability. Curr. Top. Behav. Neurosci. 2013, 14, 29–54. [Google Scholar] [CrossRef]
- Mällo, T.; Matrov, D.; Herm, L.; Kõiv, K.; Eller, M.; Rinken, A.; Harro, J. Tickling-induced 50-kHz ultrasonic vocalization is individually stable and predicts behaviour in tests of anxiety and depression in rats. Behav. Brain Res. 2007, 184, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Mällo, T.; Matrov, D.; Kõiv, K.; Harro, J. Effect of chronic stress on behavior and cerebral oxidative metabolism in rats with high or low positive affect. Neuroscience 2009, 164, 963–974. [Google Scholar] [CrossRef]
- Raudkivi, K.; Mällo, T.; Harro, J. Effect of chronic variable stress on corticosterone levels and hippocampal extracellular 5-HT in rats with persistent differences in positive affectivity. Acta Neuropsychiatr. 2012, 24, 208–214. [Google Scholar] [CrossRef]
- Kõiv, K.; Metelitsa, M.; Vares, M.; Tiitsaar, K.; Raudkivi, K.; Jaako, K.; Vulla, K.; Shimmo, R.; Harro, J. Chronic variable stress prevents amphetamine-elicited 50-kHz calls in rats with low positive affectivity. Eur. Neuropsychopharmacol. 2016, 26. [Google Scholar] [CrossRef]
- Puglisi-Allegra, S.; Imperato, A.; Angelucci, L.; Cabib, S. Acute stress induces time-dependent responses in dopamine mesolimbic system. Brain Res. 1991, 554, 217–222. [Google Scholar] [CrossRef]
- Yadid, G.; Overstreet, D.H.; Zangen, A. Limbic dopaminergic adaptation to a stressful stimulus in a rat model of depression. Brain Res. 2001, 896, 43–47. [Google Scholar] [CrossRef]
- Imperato, A.; Puglisi-Allegra, S.; Casolini, P.; Angelucci, L. Changes in brain dopamine and acetylcholine release during and following stress are independent of the pituitary-adrenocortical axis. Brain Res. 1991, 538, 111–117. [Google Scholar] [CrossRef]
- Abercrombie, E.D.; Keefe, K.A.; DiFrischia, D.S.; Zigmond, M.J. Differential effect of stress on in vivo dopamine release in striatum, nucleus accumbens, and medial frontal cortex. J. Neurochem. 1989, 52, 1655–1658. [Google Scholar] [CrossRef] [PubMed]
- Gambarana, C.; Masi, F.; Tagliamonte, A.; Scheggi, S.; Ghiglieri, O.; De Montis, M.G. A chronic stress that impairs reactivity in rats also decreases dopaminergic transmission in the nucleus accumbens: A microdialysis study. J. Neurochem. 1999, 72, 2039–2046. [Google Scholar] [CrossRef]
- Di Chiara, G.; Tanda, G. Blunting of reactivity of dopamine transmission to palatable food: A biochemical marker of anhedonia in the cms model? Psychopharmacology 1997, 134, 351–353. [Google Scholar] [CrossRef]
- Di Chiara, G.; Loddo, P.; Tanda, G. Reciprocal changes in prefrontal and limbic dopamine responsiveness to aversive and rewarding stimuli after chronic mild stress: Implications for the psychobiology of depression. Biol. Psychiatry 1999, 46, 1624–1633. [Google Scholar] [CrossRef]
- Żurawek, D.; Faron-Górecka, A.; Kuśmider, M.; Kolasa, M.; Gruca, P.; Papp, M.; Dziedzicka-Wasylewska, M. Mesolimbic dopamine D2 receptor plasticity contributes to stress resilience in rats subjected to chronic mild stress. Psychopharmacology 2013, 227, 583–593. [Google Scholar] [CrossRef] [Green Version]
- Simola, N.; Paci, E.; Serra, M.; Costa, G.; Morelli, M. Modulation of Rat 50-kHz Ultrasonic Vocalizations by Glucocorticoid Signaling: Possible Relevance to Reward and Motivation. Int. J. Neuropsychopharmacol. 2018, 21, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Popik, P.; Kos, T.; Pluta, H.; Nikiforuk, A.; Rojek, K.; Ryguła, R. Inhibition of the glucocorticoid synthesis reverses stress-induced decrease in rat’s 50-kHz ultrasonic vocalizations. Behav. Brain Res. 2014, 260, 53–57. [Google Scholar] [CrossRef]
- Russo, S.J.; Nestler, E.J. The brain reward circuitry in mood disorders. Nat. Rev. Neurosci. 2013, 14, 609–625. [Google Scholar] [CrossRef] [Green Version]
- Richard, J.M.; Berridge, K.C. Nucleus accumbens dopamine/glutamate interaction switches modes to generate desire versus dread: D1 alone for appetitive eating but D1 and D2 together for fear. J. Neurosci. 2011, 31, 12866–12879. [Google Scholar] [CrossRef] [Green Version]
- Wintink, A.J.; Brudzynski, S.M. The related roles of dopamine and glutamate in the initiation of 50-kHz ultrasonic calls in adult rats. Pharmacol. Biochem. Behav. 2001, 70, 317–323. [Google Scholar] [CrossRef]
- Hamed, A.; Kursa, M.B. Social deprivation substantially changes multi-structural neurotransmitter signature of social interaction: Glutamate concentration in amygdala and VTA as a key factor in social encounter-induced 50-kHz ultrasonic vocalization. Eur. Neuropsychopharmacol. 2020, 37, 82–99. [Google Scholar] [CrossRef]
- O’Leary, A.; Kõiv, K.; Raudkivi, K.; Harro, J. Antidepressants differentially affect striatal amphetamine-stimulated dopamine and serotonin release in rats with high and low novelty-oriented behaviour. Pharmacol. Res. 2016, 113, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Academic Press: London, UK, 2007. [Google Scholar]
- Ripley, T.L.; Jaworski, J.; Randall, P.K.; Gonzales, R.A. Repeated perfusion with elevated potassium in in vivo microdialysis—A method for detecting small changes in extracellular dopamine. J. Neurosci. Methods 1997, 78, 7–14. [Google Scholar] [CrossRef]
- Häidkind, R.; Kivastik, T.; Eller, M.; Kolts, I.; Oreland, L.; Harro, J. Denervation of the locus coeruleus projections by treatment with the selective neurotoxin DSP-4 [N (2-chloroethyl)-N-ethyl-2-bromobenzylamine] reduces dopamine release potential in the nucleus accumbens shell in conscious rats. Neurosci. Lett. 2002, 332, 79–82. [Google Scholar] [CrossRef]
- Del Arco, A.; Segovia, G.; Prieto, L.; Mora, F. Endogenous glutamate-taurine interaction in striatum and nucleus accumbens of the freely moving rat: Studies during the normal process of aging. Mech. Ageing Dev. 2001, 122, 401–414. [Google Scholar] [CrossRef]
- Behrens, P.F.; Franz, P.; Woodman, B.; Lindenberg, K.S.; Landwehrmeyer, G.B. Impaired glutamate transport and glutamate–glutamine cycling: Downstream effects of the Huntington mutation. Brain 2002, 125, 1908–1922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridges, R.J.; Stanley, M.S.; Chamberlin, A.R.; Anderson, M.W.; Cotman, C.W. Conformationally defined neurotransmitter analogues. Selective inhibition of glutamate uptake by one pyrrolidine-2,4-dicarboxylate diastereomer. J. Med. Chem. 1991, 34, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, O.; Piras, G.; Corda, M.G. The psychogenetically selected Roman high- and low-avoidance rat lines: A model to study the individual vulnerability to drug addiction. Neurosci. Biobehav. Rev. 2007, 31, 148–163. [Google Scholar] [CrossRef]
- Shimamoto, A.; Holly, E.N.; Boyson, C.O.; Debold, J.F.; Miczek, K.A. Individual differences in anhedonic and accumbal dopamine responses to chronic social stress and their link to cocaine self-administration in female rats. Psychopharmacology 2015, 232, 825–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verheij, M.M.M.; Cools, A.R. Twenty years of dopamine research: Individual differences in the response of accumbal dopamine to environmental and pharmacological challenges. Eur. J. Pharmacol. 2008, 585, 228–244. [Google Scholar] [CrossRef]
- Mulvihill, K.G.; Brudzynski, S.M. Effect of microinjections of dopamine into the nucleus accumbens shell on emission of 50 kHz USV: Comparison with effects of D-amphetamine. Pharmacol. Biochem. Behav. 2019, 176, 23–32. [Google Scholar] [CrossRef]
- Simola, N.; Brudzynski, S.M. Rat 50-kHz ultrasonic vocalizations as a tool in studying neurochemical mechanisms that regulate positive emotional states. J. Neurosci. Methods 2018, 310, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Brudzynski, S.M.; Silkstone, M.; Komadoski, M.; Scullion, K.; Duffus, S.; Burgdorf, J.; Kroes, R.A.; Moskal, J.R.; Panksepp, J. Effects of intraaccumbens amphetamine on production of 50kHz vocalizations in three lines of selectively bred Long-Evans rats. Behav. Brain Res. 2011, 217, 32–40. [Google Scholar] [CrossRef]
- Wisłowska-Stanek, A.; Płaźnik, A.; Kołosowska, K.; Skórzewska, A.; Turzyńska, D.; Liguz-Lęcznar, M.; Krząścik, P.; Gryz, M.; Szyndler, J.; Sobolewska, A.; et al. Differences in the dopaminergic reward system in rats that passively and actively behave in the Porsolt test. Behav. Brain Res. 2019, 359, 181–189. [Google Scholar] [CrossRef]
- Rakovska, A.; Baranyi, M.; Windisch, K.; Petkova-Kirova, P.; Gagov, H.; Kalfin, R. Neurochemical evidence that cocaine- and amphetamine-regulated transcript (CART) 55–102 peptide modulates the dopaminergic reward system by decreasing the dopamine release in the mouse nucleus accumbens. Brain Res. Bull. 2017, 134, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Bubar, M.J.; McMahon, L.R.; De Deurwaerdère, P.; Spampinato, U.; Cunningham, K.A. Selective serotonin reuptake inhibitors enhance cocaine-induced locomotor activity and dopamine release in the nucleus accumbens. Neuropharmacology 2003, 44, 342–353. [Google Scholar] [CrossRef]
- Amato, J.L.; Bankson, M.G.; Yamamoto, B.K. Prior exposure to chronic stress and MDMA potentiates mesoaccumbens dopamine release mediated by the 5-HT1B receptor. Neuropsychopharmacology 2007, 32, 946–954. [Google Scholar] [CrossRef] [PubMed]
- Cathala, A.; Devroye, C.; Maitre, M.; Piazza, P.V.; Abrous, D.N.; Revest, J.M.; Spampinato, U. Serotonin2C receptors modulate dopamine transmission in the nucleus accumbens independently of dopamine release: Behavioral, neurochemical and molecular studies with cocaine. Addict. Biol. 2015, 20, 445–457. [Google Scholar] [CrossRef]
- Yoshimoto, K.; Watanabe, Y.; Tanaka, M.; Kimura, M. Serotonin 2C receptors in the nucleus accumbens are involved in enhanced alcohol-drinking behavior. Eur. J. Neurosci. 2012, 35, 1368–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowry, C.A.; Hale, M.W.; Plant, A.; Windle, R.J.; Shanks, N.; Wood, S.A.; Ingram, C.D.; Renner, K.J.; Lightman, S.L.; Summers, C.H. Fluoxetine inhibits corticotropin-releasing factor (CRF)-induced behavioural responses in rats. Stress 2009, 12, 225–239. [Google Scholar] [CrossRef]
- Segovia, G.; Del Arco, A.; Mora, F. Effects of aging on the interaction between glutamate, dopamine, and GABA in striatum and nucleus accumbens of the awake rat. J. Neurochem. 1999, 73, 2063–2072. [Google Scholar] [CrossRef]
- Segovia, G.; Mora, F. Involvement of NMDA and AMPA/kainate receptors in the effects of endogenous glutamate on extracellular concentrations of dopamine and GABA in the nucleus accumbens of the awake rat. Brain Res. Bull. 2001, 54, 153–157. [Google Scholar] [CrossRef]
- Yu, J.; Ishikawa, M.; Wang, J.; Schlüter, O.M.; Sesack, S.R.; Dong, Y. Ventral tegmental area projection regulates glutamatergic transmission in nucleus accumbens. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Krupina, N.A.; Shirenova, S.D.; Khlebnikova, N.N. Prolonged Social Isolation, Started Early in Life, Impairs Cognitive Abilities in Rats Depending on Sex. Brain Sci. 2020, 10, 799. [Google Scholar] [CrossRef] [PubMed]
- Noschang, C.; Lampert, C.; Krolow, R.; de Almeida, R.M.M. Social isolation at adolescence: A systematic review on behaviour related to cocaine, amphetamine and nicotine use in rats and mice. Psychopharmacology 2021, 238, 927–947. [Google Scholar] [CrossRef] [PubMed]
- Kõiv, K.; Vares, M.; Kroon, C.; Metelitsa, M.; Tiitsaar, K.; Laugus, K.; Jaako, K.; Harro, J. Effect of chronic variable stress on sensitization to amphetamine in high and low sucrose-consuming rats. J. Psychopharmacol. 2019, 33. [Google Scholar] [CrossRef]
- Kanarik, M.; Alttoa, A.; Matrov, D.; Kõiv, K.; Sharp, T.; Panksepp, J.; Harro, J. Brain responses to chronic social defeat stress: Effects on regional oxidative metabolism as a function of a hedonic trait, and gene expression in susceptible and resilient rats. Eur. Neuropsychopharmacol. 2011, 21, 92–107. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kõiv, K.; Tiitsaar, K.; Laugus, K.; Harro, J. Extracellular Dopamine Levels in Nucleus Accumbens after Chronic Stress in Rats with Persistently High vs. Low 50-kHz Ultrasonic Vocalization Response. Brain Sci. 2021, 11, 470. https://doi.org/10.3390/brainsci11040470
Kõiv K, Tiitsaar K, Laugus K, Harro J. Extracellular Dopamine Levels in Nucleus Accumbens after Chronic Stress in Rats with Persistently High vs. Low 50-kHz Ultrasonic Vocalization Response. Brain Sciences. 2021; 11(4):470. https://doi.org/10.3390/brainsci11040470
Chicago/Turabian StyleKõiv, Kadri, Kai Tiitsaar, Karita Laugus, and Jaanus Harro. 2021. "Extracellular Dopamine Levels in Nucleus Accumbens after Chronic Stress in Rats with Persistently High vs. Low 50-kHz Ultrasonic Vocalization Response" Brain Sciences 11, no. 4: 470. https://doi.org/10.3390/brainsci11040470
APA StyleKõiv, K., Tiitsaar, K., Laugus, K., & Harro, J. (2021). Extracellular Dopamine Levels in Nucleus Accumbens after Chronic Stress in Rats with Persistently High vs. Low 50-kHz Ultrasonic Vocalization Response. Brain Sciences, 11(4), 470. https://doi.org/10.3390/brainsci11040470