
The Effect of Light Sedation with Midazolam on Functional Connectivity of the Dorsal Attention Network

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
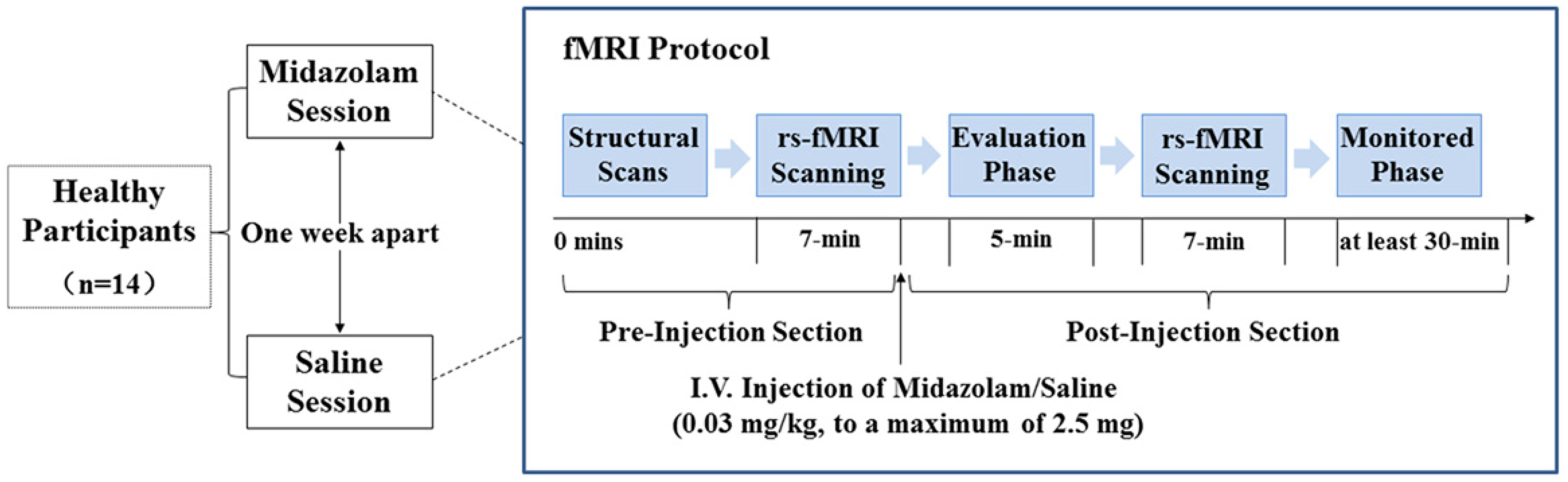
2. Materials and Methods
2.1. Participants
2.2. Image Preprocessing
2.3. Functional Connectivity Analysis
2.4. Group-Level Connectivity Analysis
3. Results
3.1. Head-Motion Estimation
3.2. Functional Connectivity Analysis
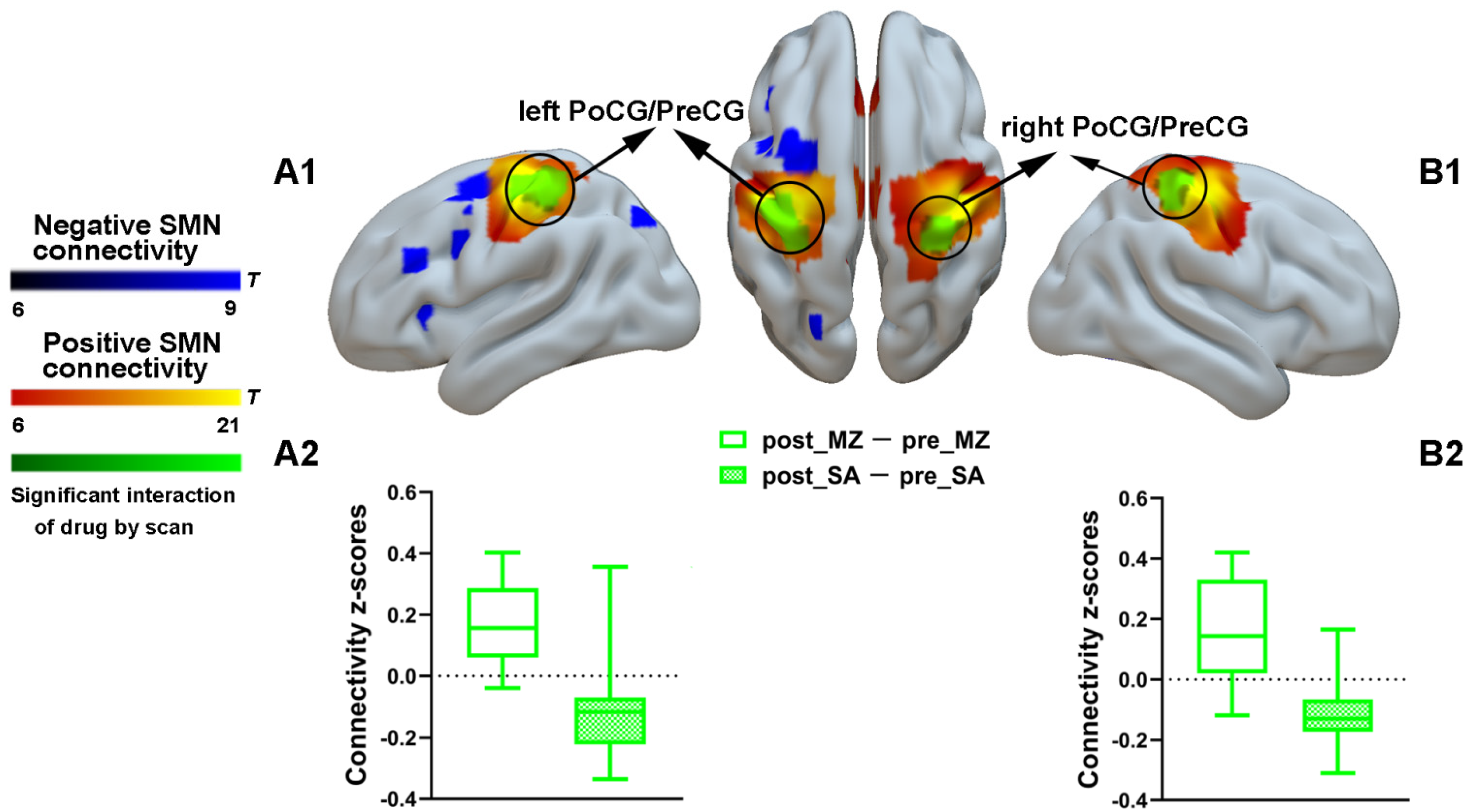
3.2.1. SMN Connectivity Results
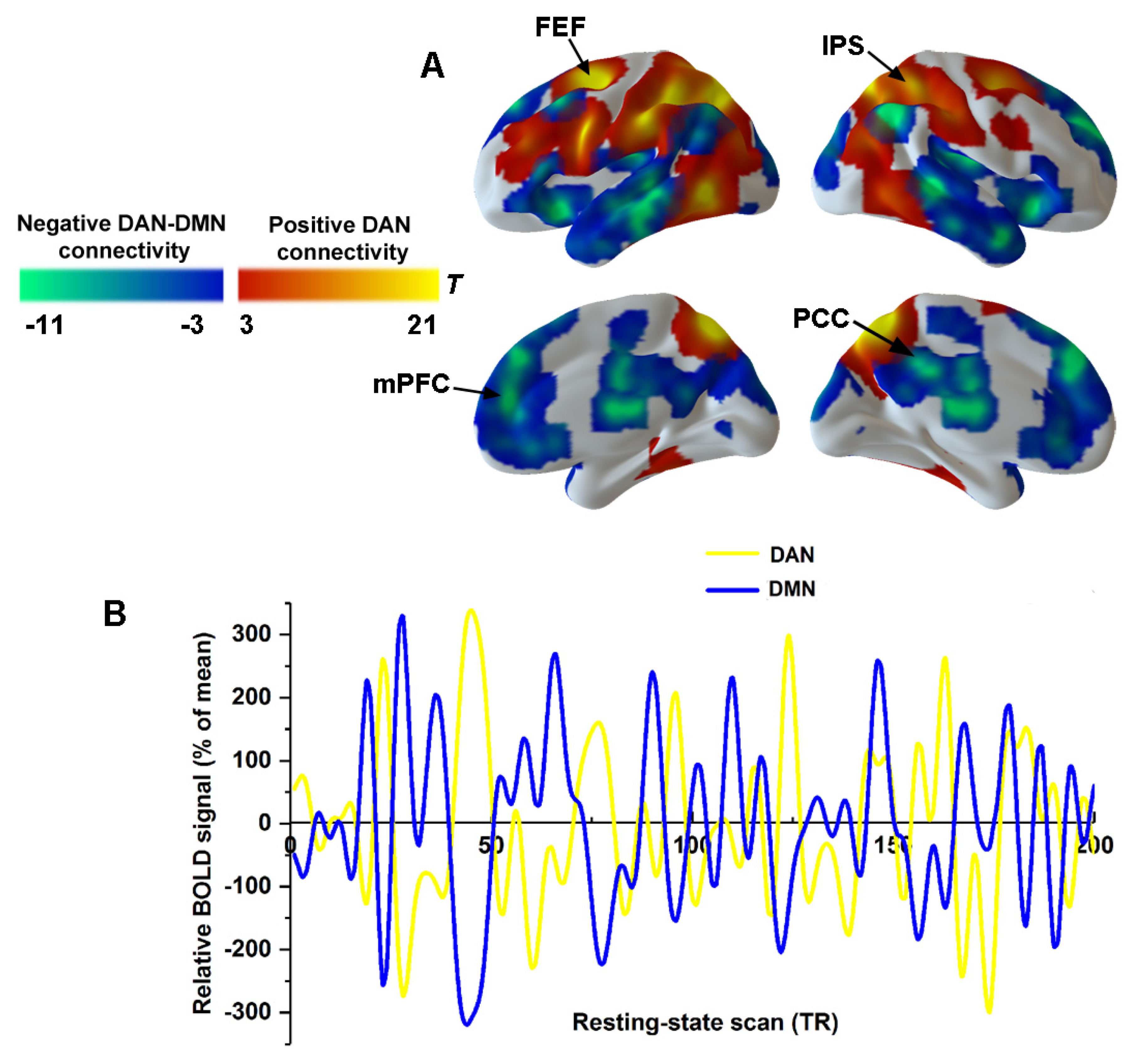
3.2.2. The Spatial Distribution of the DAN and DMN
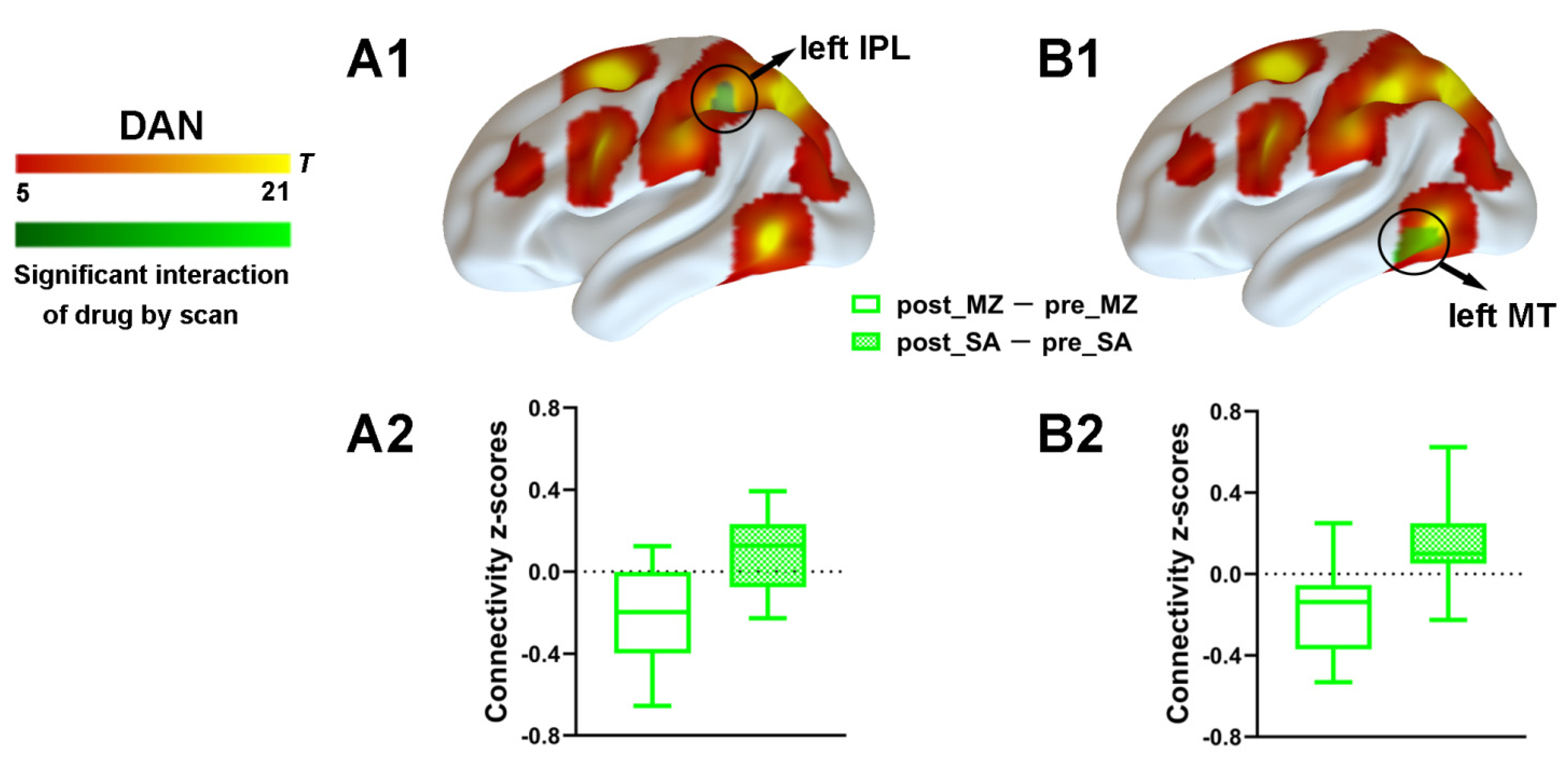
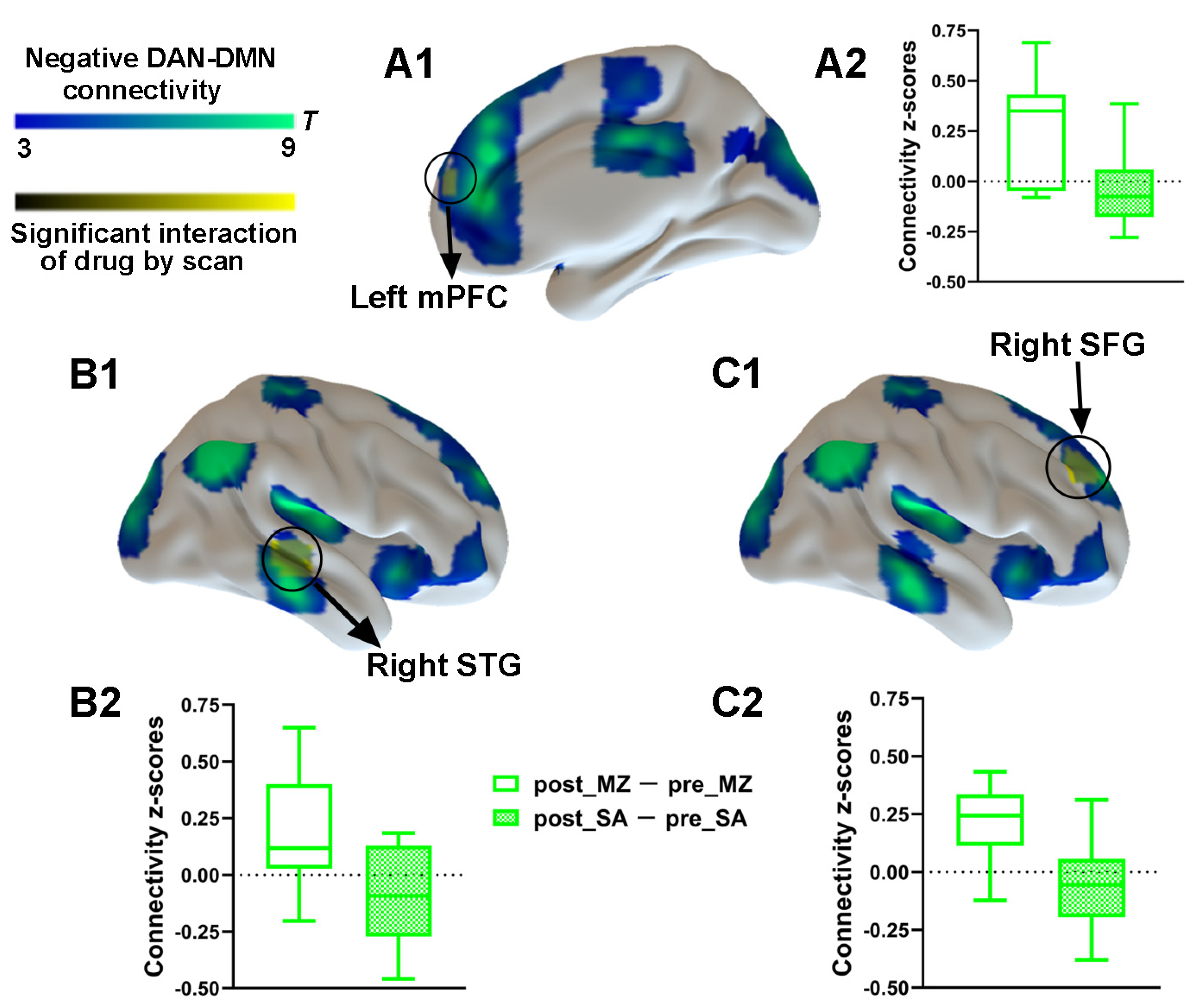
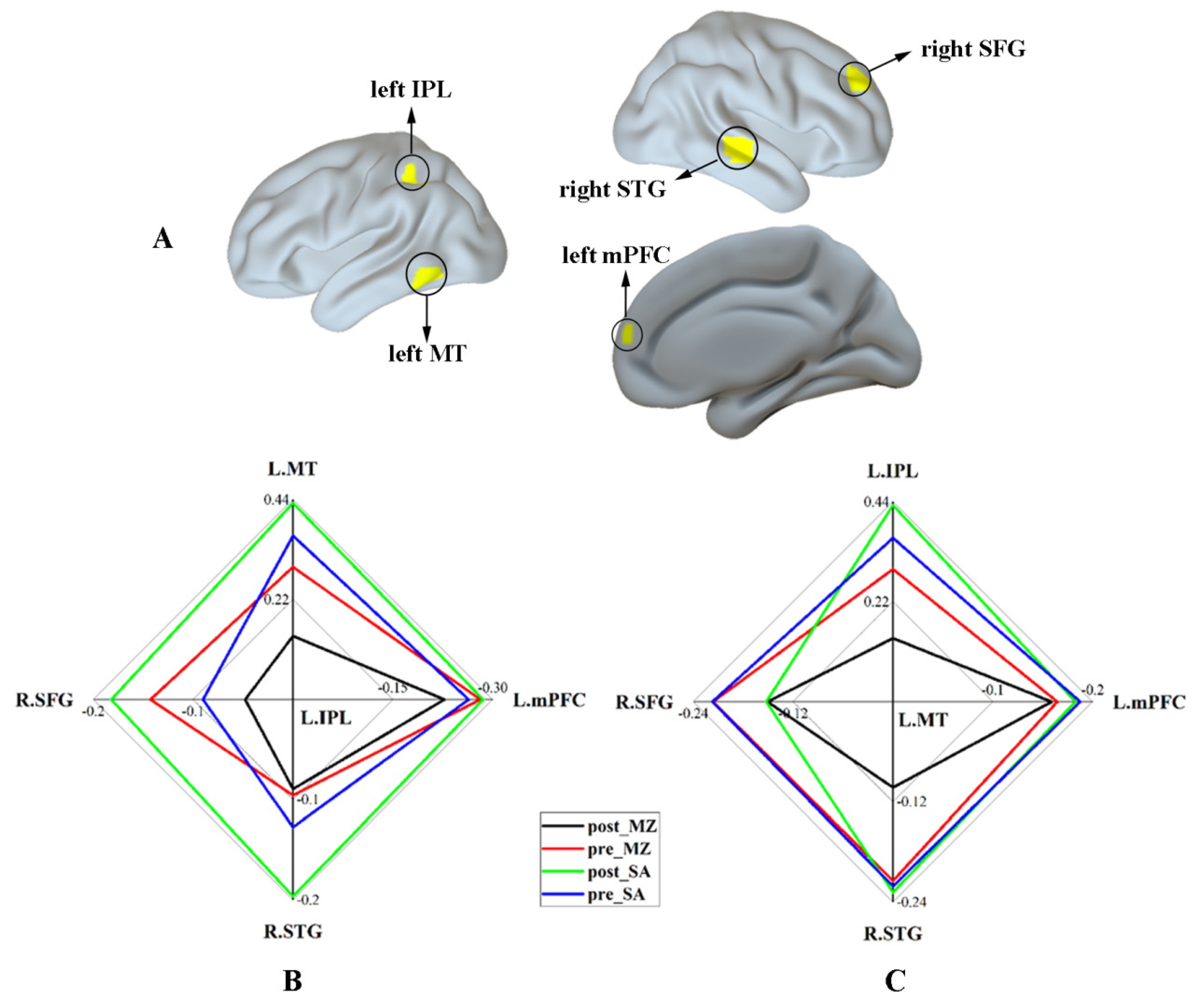
3.2.3. DAN Connectivity Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Veselis, R.A.; Reinsel, R.A.; Feshchenko, V.A. Drug-induced amnesia is a separate phenomenon from sedation: Electrophysiologic evidence. Anesthesiology 2001, 95, 896–907. [Google Scholar] [CrossRef] [PubMed]
- Hemmings, H.C., Jr.; Riegelhaupt, P.M.; Kelz, M.B.; Solt, K.; Eckenhoff, R.G.; Orser, B.A.; Goldstein, P.A. Towards a comprehensive understanding of anesthetic mechanisms of action: A decade of discovery. Trends Pharm. Sci. 2019, 40, 464–481. [Google Scholar] [CrossRef] [PubMed]
- Biswal, B.B. Resting state fMRI: A personal history. NeuroImage 2012, 62, 938–944. [Google Scholar] [CrossRef] [PubMed]
- Fox, M.D.; Greicius, M. Clinical applications of resting state functional connectivity. Front. Syst. Neurosci. 2010, 4, 19. [Google Scholar] [CrossRef] [Green Version]
- Buckner, R.L.; Krienen, F.M.; Yeo, B.T. Opportunities and limitations of intrinsic functional connectivity MRI. Nat. Neurosci. 2013, 16, 832–837. [Google Scholar] [CrossRef] [PubMed]
- Logothetis, N.K.; Pauls, J.; Augath, M.; Trinath, T.; Oeltermann, A. Neurophysiological investigation of the basis of the fMRI signal. Nature 2001, 412, 150–157. [Google Scholar] [CrossRef]
- Fox, M.D.; Raichle, M.E. Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nat. Rev. Neurosci. 2007, 8, 700–711. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, A.A.; Naci, L.; MacDonald, P.A.; Owen, A.M. Anesthesia and neuroimaging: Investigating the neural correlates of unconsciousness. Trends Cogn. Sci. 2015, 19, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Boveroux, P.; Vanhaudenhuyse, A.; Bruno, M.A.; Noirhomme, Q.; Lauwick, S.; Luxen, A.; Degueldre, C.; Plenevaux, A.; Schnakers, C.; Phillips, C.; et al. Breakdown of within-and between-network resting state functional magnetic resonance imaging connectivity during propofol-induced loss of consciousness. Anesthesiology 2010, 113, 1038–1053. [Google Scholar] [CrossRef] [Green Version]
- Mhuircheartaigh, R.N.; Rosenorn-Lanng, D.; Wise, R.; Jbabdi, S.; Rogers, R.; Tracey, I. Cortical and subcortical connectivity changes during decreasing levels of consciousness in humans: A functional magnetic resonance imaging study using propofol. J. Neurosci. 2010, 30, 9095–9102. [Google Scholar] [CrossRef]
- Jordan, D.; Ilg, R.; Riedl, V.; Schorer, A.; Grimberg, S.; Neufang, S.; Omerovic, A.; Berger, S.; Untergehrer, G.; Preibisch, C.; et al. Simultaneous electroencephalographic and functional magnetic resonance imaging indicate impaired cortical top-down processing in association with anesthetic-induced unconsciousness. Anesthesiology 2013, 119, 1031–1042. [Google Scholar] [CrossRef] [Green Version]
- Ranft, A.; Golkowski, D.; Kiel, T.; Riedl, V.; Kohl, P.; Rohrer, G.; Pientka, J.; Berger, S.; Thul, A.; Maurer, M.; et al. Neural correlates of sevoflurane-induced unconsciousness identified by simultaneous functional magnetic resonance imaging and electroencephalography. Anesthesiology 2016, 125, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Bonhomme, V.; Vanhaudenhuyse, A.; Demertzi, A.; Bruno, M.-A.; Jaquet, O.; Bahri, M.A.; Plenevaux, A.; Boly, M.; Boveroux, P.; Soddu, A.; et al. Resting-state network-specific breakdown of functional connectivity during ketamine alteration of consciousness in volunteers. Anesthesiology 2016, 125, 873–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkire, M.T.; Hudetz, A.G.; Tononi, G. Consciousness and anesthesia. Science 2008, 322, 876–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayne, T.; Hohwy, J.; Owen, A.M. Are there levels of consciousness? Trends Cogn. Sci. 2016, 20, 405–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashour, G.A.; Hudetz, A.G. Bottom-up and top-down mechanisms of general anesthetics modulate different dimensions of consciousness. Front. Neural Circuits 2017, 11, 44. [Google Scholar] [CrossRef] [Green Version]
- Koch, C.; Massimini, M.; Boly, M.; Tononi, G. Neural correlates of consciousness: Progress and problems. Nat. Rev. Neurosci. 2016, 17, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Sanders, R.D.; Tononi, G.; Laureys, S.; Sleigh, J.W. Unresponsiveness ≠ unconsciousness. Anesthesiology 2012, 116, 946–959. [Google Scholar] [CrossRef] [Green Version]
- Bonhomme, V.; Staquet, C.; Montupil, J.; Defresne, A.; Kirsch, M.; Martial, C.; Vanhaudenhuyse, A.; Chatelle, C.; Larroque, S.K.; Raimondo, F.; et al. General anesthesia: A probe to explore consciousness. Front. Syst. Neurosci. 2019, 13, 36. [Google Scholar] [CrossRef] [Green Version]
- Vanhaudenhuyse, A.; Demertzi, A.; Schabus, M.; Noirhomme, Q.; Bredart, S.; Boly, M.; Phillips, C.; Soddu, A.; Luxen, A.; Moonen, G.; et al. Two distinct neuronal networks mediate the awareness of environment and of self. J. Cogn. Neurosci. 2011, 23, 570–578. [Google Scholar] [CrossRef] [Green Version]
- Fox, M.D.; Snyder, A.Z.; Vincent, J.L.; Corbetta, M.; van Essen, D.C.; Raichle, M.E. The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proc. Natl. Acad. Sci. USA 2005, 102, 9673–9678. [Google Scholar] [CrossRef] [Green Version]
- Petersen, S.E.; Posner, M.I. The attention system of the human brain: 20 years after. Annu. Rev. Neurosci. 2012, 35, 73–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greicius, M.D.; Kiviniemi, V.; Tervonen, O.; Vainionpää, V.; Alahuhta, S.; Reiss, A.L.; Menon, V. Persistent default-mode network connectivity during light sedation. Hum. Brain Mapp. 2008, 29, 839–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, E.A.; Adapa, R.M.; Absalom, A.R.; Menon, D.K. Changes in resting neural connectivity during propofol sedation. PLoS ONE 2010, 5, e14224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martuzzi, R.; Ramani, R.; Qiu, M.; Rajeevan, N.; Constable, R. Functional connectivity and alterations in baseline brain state in humans. NeuroImage 2010, 49, 823–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, P.; Zhang, H.; Xu, Y.; Jia, W.; Zang, Y.; Li, K. Disruption of cortical integration during midazolam-induced light sedation. Hum. Brain Mapp. 2015, 36, 4247–4261. [Google Scholar] [CrossRef] [Green Version]
- Leech, R.; Kamourieh, S.; Beckmann, C.F.; Sharp, D.J. Fractionating the default mode network: Distinct contributions of the ventral and dorsal posterior cingulate cortex to cognitive control. J. Neurosci. 2011, 31, 3217–3224. [Google Scholar] [CrossRef]
- Di Perri, C.; Bahri, M.A.; Amico, E.; Thibaut, A.; Heine, L.; Antonopoulos, G.; Charland-Verville, V.; Wannez, S.; Gomez, F.; Hustinx, R.; et al. Neural correlates of consciousness in patients who have emerged from a minimally conscious state: A cross-sectional multimodal imaging study. Lancet Neurol. 2016, 15, 830–842. [Google Scholar] [CrossRef]
- Nyhus, E.; Curran, T. Midazolam-induced amnesia reduces memory for details and affects the ERP correlates of recollection and familiarity. J. Cogn. Neurosci. 2012, 24, 416–427. [Google Scholar] [CrossRef] [PubMed]
- Chernik, D.A.; Gillings, D.; Laine, H.; Hendler, J.; Silver, J.M.; Davidson, A.B.; Schwam, E.M.; Siegel, J.L. Validity and reliability of the Observer’s Assessment of Alertness/Sedation Scale: Study with intravenous midazolam. J. Clin. Psychopharmacol. 1990, 10, 244–251. [Google Scholar] [PubMed]
- Whitfield-Gabrieli, S.; Nieto-Castanon, A. Conn: A functional connectivity toolbox for correlated and anticorrelated brain networks. Brain Connect. 2012, 2, 125–141. [Google Scholar] [CrossRef] [Green Version]
- Behzadi, Y.; Restom, K.; Liau, J.; Liu, T.T. A component based noise correction method (CompCor) for BOLD and perfusion based fMRI. NeuroImage 2007, 37, 90–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, K.; Birn, R.M.; Handwerker, D.; Jones, T.B.; Bandettini, P.A. The impact of global signal regression on resting state correlations: Are anti-correlated networks introduced? NeuroImage 2009, 44, 893–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissenbacher, A.; Kasess, C.; Gerstl, F.; Lanzenberger, R.; Moser, E.; Windischberger, C. Correlations and anticorrelations in resting-state functional connectivity MRI: A quantitative comparison of preprocessing strategies. NeuroImage 2009, 47, 1408–1416. [Google Scholar] [CrossRef] [PubMed]
- Demertzi, A.; Antonopoulos, G.; Heine, L.; Voss, H.U.; Crone, J.S.; Angeles, C.D.L.; Bahri, M.A.; di Perri, C.; Vanhaudenhuyse, A.; Charland-Verville, V.; et al. Intrinsic functional connectivity differentiates minimally conscious from unresponsive patients. Brain 2015, 138, 2619–2631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas Yeo, B.T.; Krienen, F.M.; Sepulcre, J.; Sabuncu, M.R.; Lashkari, D.; Hollinshead, M.; Roffman, J.L.; Smoller, J.W.; Zöllei, L.; Polimeni, J.R.; et al. The organization of the human cerebral cortex estimated by intrinsic functional connectivity. J. Neurophysiol. 2011, 106, 1125–1165. [Google Scholar] [CrossRef]
- Raichle, M.E. The restless brain. Brain Connect. 2011, 1, 3–12. [Google Scholar] [CrossRef]
- Power, J.D.; Barnes, K.A.; Snyder, A.Z.; Schlaggar, B.L.; Petersen, S.E. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. NeuroImage 2012, 59, 2142–2154. [Google Scholar] [CrossRef] [Green Version]
- Kiviniemi, V.J.; Haanpää, H.; Kantola, J.-H.; Jauhiainen, J.; Vainionpää, V.; Alahuhta, S.; Tervonen, O. Midazolam sedation increases fluctuation and synchrony of the resting brain BOLD signal. Magn. Reson. Imaging 2005, 23, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Zuo, X.-N.; di Martino, A.; Kelly, C.; Shehzad, Z.E.; Gee, D.; Klein, D.F.; Castellanos, F.; Biswal, B.B.; Milham, M.P. The oscillating brain: Complex and reliable. NeuroImage 2010, 49, 1432–1445. [Google Scholar] [CrossRef] [Green Version]
- Kuhnke, P.; Beaupain, M.C.; Cheung, V.K.; Weise, K.; Kiefer, M.; Hartwigsen, G. Left posterior inferior parietal cortex causally supports the retrieval of action knowledge. NeuroImage 2020, 219, 117041. [Google Scholar] [CrossRef]
- Chen, Q.; Garcea, F.E.; Jacobs, R.; Mahon, B.Z. Abstract representations of object-directed action in the left inferior parietal lobule. Cereb. Cortex 2017, 28, 2162–2174. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, L.D.; Khawaja, F.A.; Pack, C.C.; Butts, D.A. Diverse suppressive influences in area MT and selectivity to complex motion features. J. Neurosci. 2013, 33, 16715–16728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demertzi, A.; Gómez, F.; Crone, J.S.; Vanhaudenhuyse, A.; Tshibanda, L.; Noirhomme, Q.; Thonnard, M.; Charland-Verville, V.; Kirsch, M.; Laureys, S.; et al. Multiple fMRI system-level baseline connectivity is disrupted in patients with consciousness alterations. Cortex 2014, 52, 35–46. [Google Scholar] [CrossRef]
- Martuzzi, R.; Ramani, R.; Qiu, M.; Shen, X.; Papademetris, X.; Constable, R.T. A whole-brain voxel based measure of intrinsic connectivity contrast reveals local changes in tissue connectivity with anesthetic without a priori assumptions on thresholds or regions of interest. NeuroImage 2011, 58, 1044–1050. [Google Scholar] [CrossRef] [Green Version]
- Khalsa, S.; Mayhew, S.D.; Chechlacz, M.; Bagary, M.; Bagshaw, A.P. The structural and functional connectivity of the posterior cingulate cortex: Comparison between deterministic and probabilistic tractography for the investigation of structure—function relationships. NeuroImage 2014, 102, 118–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, J.L.; Patel, G.H.; Fox, M.D.; Snyder, A.Z.; Baker, J.; van Essen, D.C.; Zempel, J.M.; Snyder, L.H.; Corbetta, M.; Raichle, M.E. Intrinsic functional architecture in the anaesthetized monkey brain. Nat. Cell Biol. 2007, 447, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Boly, M.; Tshibanda, L.; Vanhaudenhuyse, A.; Noirhomme, Q.; Schnakers, C.; Ledoux, D.; Boveroux, P.; Garweg, C.; Lambermont, B.; Phillips, C.; et al. Functional connectivity in the default network during resting state is preserved in a vegetative but not in a brain dead patient. Hum. Brain Mapp. 2009, 30, 2393–2400. [Google Scholar] [CrossRef] [Green Version]
- Jobson, D.D.; Hase, Y.; Clarkson, A.N.; Kalaria, R.N. The role of the medial prefrontal cortex in cognition, ageing and dementia. Brain Commun. 2021, 3, fcab125. [Google Scholar] [CrossRef] [PubMed]
- Günseli, E.; Aly, M. Preparation for upcoming attentional states in the hippocampus and medial prefrontal cortex. eLife 2020, 9, 53191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Pisapia, N.; Turatto, M.; Lin, P.; Jovicich, J.; Caramazza, A. Unconscious priming instructions modulate activity in default and executive networks of the human brain. Cereb. Cortex 2012, 22, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Kelly, C.; Uddin, L.; Biswal, B.B.; Castellanos, F.X.; Milham, M.P. Competition between functional brain networks mediates behavioral variability. NeuroImage 2008, 39, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, J.; Wang, Z.; Sun, P.; Li, K.; Liang, P. Dysfunctional interactions between the default mode network and the dorsal attention network in subtypes of amnestic mild cognitive impairment. Aging 2019, 11, 9147–9166. [Google Scholar] [CrossRef] [PubMed]
- Porges, E.C.; Jensen, G.; Foster, B.; Edden, R.A.; Puts, N.A. The trajectory of cortical GABA across the lifespan, an individual participant data meta-analysis of edited MRS studies. eLife 2021, 10, 62575. [Google Scholar] [CrossRef]
- Cuypers, K.; Hehl, M.; van Aalst, J.; Chalavi, S.; Mikkelsen, M.; van Laere, K.; Dupont, P.; Mantini, D.; Swinnen, S.P. Age-related GABAergic differences in the primary sensorimotor cortex: A multimodal approach combining PET, MRS and TMS. NeuroImage 2021, 226, 117536. [Google Scholar] [CrossRef] [PubMed]
- Engin, E.; Benham, R.S.; Rudolph, U. An emerging circuit pharmacology of GABAA receptors. Trends Pharmacol. Sci. 2018, 39, 710–732. [Google Scholar] [CrossRef]
- Barttfeld, P.; Uhrig, L.; Sitt, J.D.; Sigman, M.; Jarraya, B.; Dehaene, S. Signature of consciousness in the dynamics of resting-state brain activity. Proc. Natl. Acad. Sci. USA 2015, 112, 887–892. [Google Scholar] [CrossRef] [Green Version]
- Thirion, B.; Pinel, P.; Meriaux, S.; Roche, A.; Dehaene, S.; Poline, J.B. Analysis of a large fMRI cohort: Statistical and methodological issues for group analyses. NeuroImage 2007, 35, 105–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.; Utevsky, A.V.; Bland, A.R.; Clement, N.; Clithero, J.; Harsch, A.E.; Carter, R.M.; Huettel, S.A. Characterizing individual differences in functional connectivity using dual-regression and seed-based approaches. NeuroImage 2014, 95, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Birn, R.M.; Smith, M.A.; Jones, T.B.; Bandettini, P.A. The respiration response function: The temporal dynamics of fMRI signal fluctuations related to changes in respiration. NeuroImage 2008, 40, 644–654. [Google Scholar] [CrossRef] [Green Version]
- Shmueli, K.; van Gelderen, P.; de Zwart, J.A.; Horovitz, S.G.; Fukunaga, M.; Jansma, J.M.; Duyn, J.H. Low-frequency fluctuations in the cardiac rate as a source of variance in the resting-state fMRI BOLD signal. NeuroImage 2007, 38, 306–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Vlisides, P.E.; Kelz, M.B.; Avidan, M.S.; Mashour, G.A. Dynamic cortical connectivity during general anesthesia in healthy volunteers. Anesthesiology 2019, 130, 870–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Zhang, J.; Wu, J.; Mashour, G.A.; Hudetz, A.G. Temporal circuit of macroscale dynamic brain activity supports human consciousness. Sci. Adv. 2020, 6, eaaz0087. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Xu, Y.; Deshpande, G.; Li, K.; Sun, P.; Liang, P. The Effect of Light Sedation with Midazolam on Functional Connectivity of the Dorsal Attention Network. Brain Sci. 2021, 11, 1107. https://doi.org/10.3390/brainsci11081107
Wang J, Xu Y, Deshpande G, Li K, Sun P, Liang P. The Effect of Light Sedation with Midazolam on Functional Connectivity of the Dorsal Attention Network. Brain Sciences. 2021; 11(8):1107. https://doi.org/10.3390/brainsci11081107
Chicago/Turabian StyleWang, Junkai, Yachao Xu, Gopikrishna Deshpande, Kuncheng Li, Pei Sun, and Peipeng Liang. 2021. "The Effect of Light Sedation with Midazolam on Functional Connectivity of the Dorsal Attention Network" Brain Sciences 11, no. 8: 1107. https://doi.org/10.3390/brainsci11081107
APA StyleWang, J., Xu, Y., Deshpande, G., Li, K., Sun, P., & Liang, P. (2021). The Effect of Light Sedation with Midazolam on Functional Connectivity of the Dorsal Attention Network. Brain Sciences, 11(8), 1107. https://doi.org/10.3390/brainsci11081107