
The Effects of Intraoperative Hypothermia on Postoperative Cognitive Function in the Rat Hippocampus and Its Possible Mechanisms




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
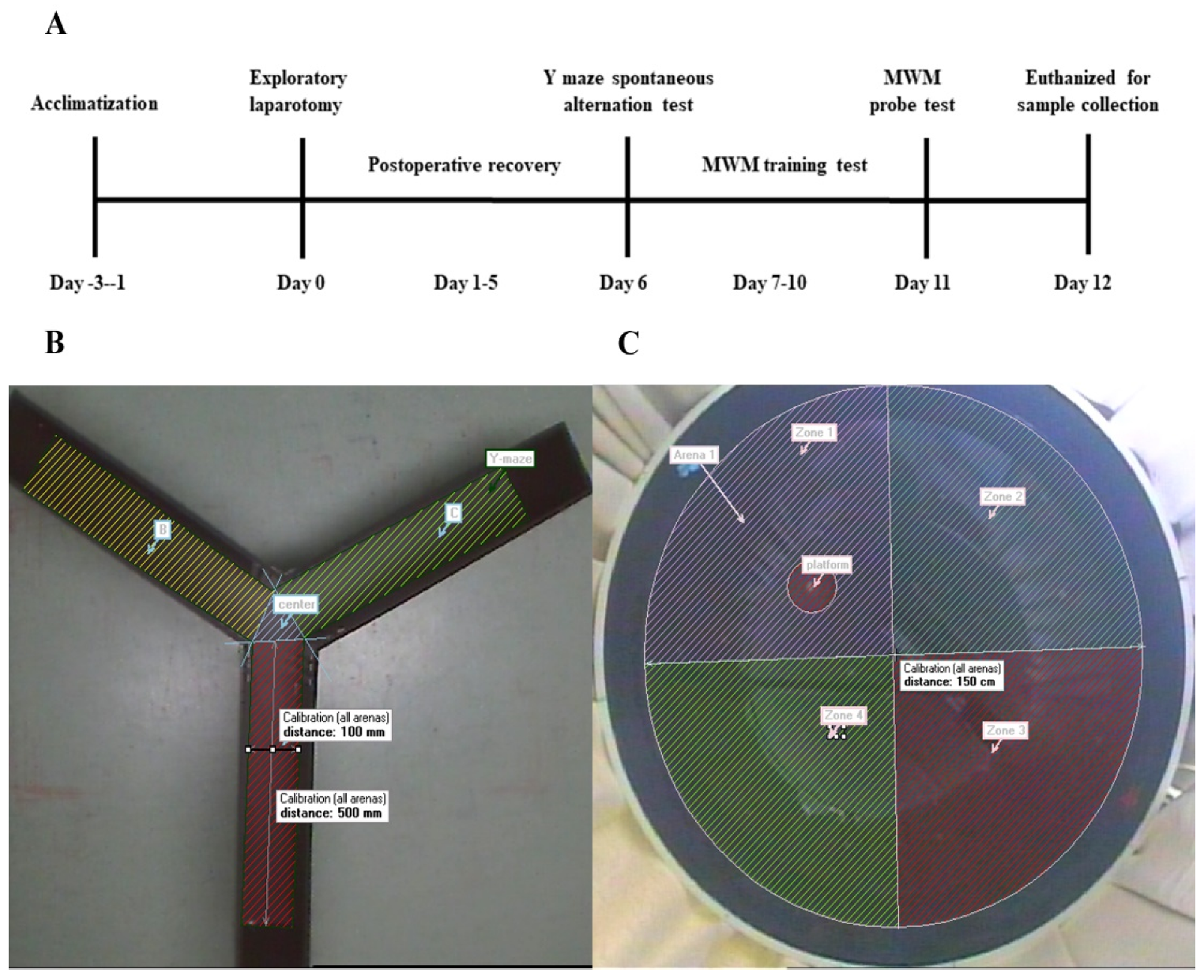
2.2. Protocols
2.3. Surgery
2.4. Behavioural Tests
2.5. Y-Maze Test
2.6. MWM Test
2.7. Haematoxylin-Eosin (HE) Staining
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
3.1. Intraoperative Hypothermia Impaired Rats’ Spatial Working Memory
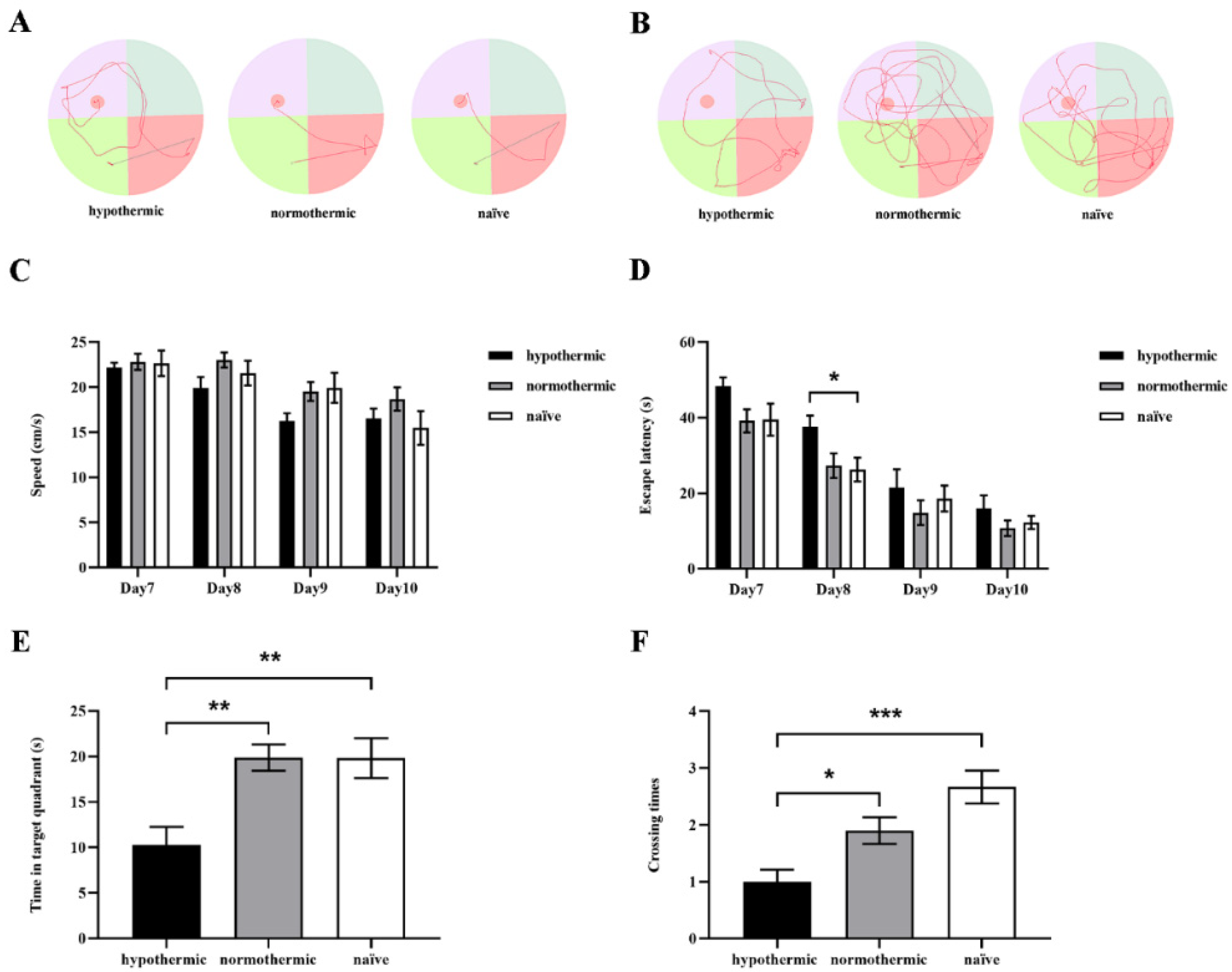
3.2. Intraoperative Hypothermia Reduced Rats’ Postoperative Spatial Learning and Memory Ability
3.3. Intraoperative Hypothermia Increased Hippocampal Neuron Injuries in Postoperative Rats
3.4. Intraoperative Hypothermia Suppressed the Expression of Synaptic Plasticity-Related Proteins in the Hippocampal Region of Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sari, S.; Aksoy, S.M.; But, A. The incidence of inadvertent perioperative hypothermia in patients undergoing general anesthesia and an examination of risk factors. Int. J. Clin. Pract. 2021, 75, e14103. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Lei, Y.; Xu, S.; Si, Y.; Li, S.; Xia, Z.; Shi, Y.; Gu, X.; Yu, J.; Xu, G.; et al. Intraoperative hypothermia and its clinical outcomes in patients undergoing general anesthesia: National study in China. PLoS ONE 2017, 12, e0177221. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Honar, H.; Sessler, D.I.; Dalton, J.E.; Yang, D.; Panjasawatwong, K.; Deroee, A.F.; Salmasi, V.; Saager, L.; Kurz, A. Intraoperative core temperature patterns, transfusion requirement, and hospital duration in patients warmed with forced air. Anesthesiology 2015, 122, 276–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmied, H.; Kurz, A.; Sessler, D.I.; Kozek, S.R.A. Mild hypothermia increases blood loss and transfusion requirements during total hip arthroplasty. Lancet 1996, 347, 289–292. [Google Scholar] [CrossRef]
- Frank, S.; Fleisher, L.; Breslow, M.; Higgins, M.; Olson, K.; Kelly, S.; Beattie, C. Perioperative maintenance of normothermia reduces the incidence of morbid cardiac events. A randomized clinical trial. JAMA 1997, 277, 1127–1134. [Google Scholar] [CrossRef]
- Kurz, A.; Sessler, D.I.; Lenhardt, R. Perioperative normothermia to reduce the incidence of surgical-wound infection and shorten hospitalization. Study of wound infection and temperature group. N. Engl. J. Med. 1996, 334, 1209–1215. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Xiang, Z.; Deng, X.; Fan, T.; Fu, R.; Geng, W.; Guo, R.; He, N.; Li, C.; Li, L.; et al. Incidence of Inadvertent Intraoperative Hypothermia and Its Risk Factors in Patients Undergoing General Anesthesia in Beijing: A Prospective Regional Survey. PLoS ONE 2015, 10, e0136136. [Google Scholar] [CrossRef] [Green Version]
- Yi, J.; Zhan, L.; Lei, Y.; Xu, S.; Si, Y.; Li, S.; Xia, Z.; Shi, Y.; Gu, X.; Yu, J.; et al. Establishment and Validation of a Prediction Equation to Estimate Risk of Intraoperative Hypothermia in Patients Receiving General Anesthesia. Sci. Rep. 2017, 7, 13927. [Google Scholar] [CrossRef] [Green Version]
- Needham, M.J.; Webb, C.E.; Bryden, D.C. Postoperative cognitive dysfunction and dementia: What we need to know and do. Br. J. Anaesth. 2017, 119, i115–i125. [Google Scholar] [CrossRef] [Green Version]
- Evered, L.; Silbert, B.; Knopman, D.S.; Scott, D.A.; DeKosky, S.T.; Rasmussen, L.S.; Oh, E.S.; Crosby, G.; Berger, M.; Eckenhoff, R.G.; et al. Recommendations for the nomenclature of cognitive change associated with anaesthesia and surgery-2018. Br. J. Anaesth. 2018, 121, 1005–1012. [Google Scholar] [CrossRef] [Green Version]
- Skvarc, D.R.; Berk, M.; Byrne, L.K.; Dean, O.M.; Dodd, S.; Lewis, M.; Marriott, A.; Moore, E.M.; Morris, G.; Page, R.S.; et al. Post-Operative Cognitive Dysfunction: An exploration of the inflammatory hypothesis and novel therapies. Neurosci. Biobehav. Rev. 2018, 84, 116–133. [Google Scholar] [CrossRef]
- Androsova, G.; Krause, R.; Winterer, G.; Schneider, R. Biomarkers of postoperative delirium and cognitive dysfunction. Front. Aging Neurosci. 2015, 7, 112. [Google Scholar] [CrossRef]
- Chen, C.; Gao, R.; Li, M.; Wang, Q.; Chen, H.; Zhang, S.; Mao, X.; Behensky, A.; Zhang, Z.; Gan, L.; et al. Extracellular RNAs-TLR3 signaling contributes to cognitive decline in a mouse model of postoperative cognitive dysfunction. Brain Behav. Immun. 2019, 80, 439–451. [Google Scholar] [CrossRef]
- Oberman, K.; Hovens, I.; de Haan, J.; Falcao-Salles, J.; van Leeuwen, B.; Schoemaker, R. Acute pre-operative ibuprofen improves cognition in a rat model for postoperative cognitive dysfunction. J. Neuroinflam. 2021, 18, 156. [Google Scholar] [CrossRef]
- Netto, M.B.; de Oliveira Junior, A.N.; Goldim, M.; Mathias, K.; Fileti, M.E.; da Rosa, N.; Laurentino, A.O.; de Farias, B.X.; Costa, A.B.; Rezin, G.T.; et al. Oxidative stress and mitochondrial dysfunction contributes to postoperative cognitive dysfunction in elderly rats. Brain Behav. Immun. 2018, 73, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Zhang, S.; Zhou, H.; Tao, X.; Ni, Y.; Pei, D.; Kang, S.; Yan, W.; Lu, J. Role of mTOR-Regulated Autophagy in Synaptic Plasticity Related Proteins Downregulation and the Reference Memory Deficits Induced by Anesthesia/Surgery in Aged Mice. Front. Aging Neurosci. 2021, 13, 628541. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.-Y.; Xiong, B.-R.; Zhang, W.; Zhou, W.-C.; Yang, H.; Gao, F.; Xiang, H.-B.; Manyande, A.; Tian, X.-B.; Tian, Y.-K. PGE2-EP3 signaling exacerbates hippocampus-dependent cognitive impairment after laparotomy by reducing expression levels of hippocampal synaptic plasticity-related proteins in aged mice. CNS Neurosci. Ther. 2018, 24, 917–929. [Google Scholar] [CrossRef]
- Pacheco Estefan, D.; Sanchez-Fibla, M.; Duff, A.; Principe, A.; Rocamora, R.; Zhang, H.; Axmacher, N.; Verschure, P. Coordinated representational reinstatement in the human hippocampus and lateral temporal cortex during episodic memory retrieval. Nat. Commun. 2019, 10, 2255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellmund, J.L.S.; Gardenfors, P.; Moser, E.I.; Doeller, C.F. Navigating cognition: Spatial codes for human thinking. Science 2018, 362, eaat6766. [Google Scholar] [CrossRef] [Green Version]
- Borgomaneri, S.; Battaglia, S.; Sciamanna, G.; Tortora, F.; Laricchiuta, D. Memories are not written in stone: Re-writing fear memories by means of non-invasive brain stimulation and optogenetic manipulations. Neurosci. BioBehav. Rev. 2021, 127, 334–352. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, S.; Garofalo, S.; di Pellegrino, G.; Starita, F. Revaluing the Role of vmPFC in the Acquisition of Pavlovian Threat Conditioning in Humans. J. Neurosci. 2020, 44, 8491–8500. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, S.; Harrison, B.J.; Fullana, M.A. Does the human ventromedial prefrontal cortex support fear learning, fear extinction or both? A commentary on subregional contributions. Mol. Psychiatry 2021, 1–3. [Google Scholar] [CrossRef]
- Borgomaneri, S.; Battaglia, S.; Garofalo, S.; Tortora, F.; Avenanti, A.; di Pellegrino, G. State-Dependent TMS over Prefrontal Cortex Disrupts Fear-Memory Reconsolidation and Prevents the Return of Fear. Curr. Biol. 2020, 30, 3672–3679.e4. [Google Scholar] [CrossRef] [PubMed]
- Mansvelder, H.D.; Verhoog, M.B.; Goriounova, N.A. Synaptic plasticity in human cortical circuits: Cellular mechanisms of learning and memory in the human brain? Curr. Opin. Neurobiol. 2019, 54, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Borgomaneri, S.; Serio, G.; Battaglia, S. Please, don’t do it! Fifteen years of progress of non-invasive brain stimulation in action inhibition. Cortex 2020, 132, 404–422. [Google Scholar] [CrossRef]
- Borgomaneri, S.; Vitale, F.; Battaglia, S.; Avenanti, A. Early Right Motor Cortex Response to Happy and Fearful Facial Expressions: A TMS Motor-Evoked Potential Study. Brain Sci. 2021, 11, 1203. [Google Scholar] [CrossRef]
- Battaglia, S.; Serio, G.; Scarpazza, C.; D’Ausilio, A.; Borgomaneri, S. Frozen in (e)motion: How reactive motor inhibition is influenced by the emotional content of stimuli in healthy and psychiatric populations. Behav. Res. Ther. 2021, 146, 103963. [Google Scholar] [CrossRef]
- Wall, M.J.; Collins, D.R.; Chery, S.L.; Allen, Z.D.; Pastuzyn, E.D.; George, A.J.; Nikolova, V.D.; Moy, S.S.; Philpot, B.D.; Shepherd, J.D.; et al. The Temporal Dynamics of Arc Expression Regulate Cognitive Flexibility. Neuron 2018, 98, 1124–1132.e7. [Google Scholar] [CrossRef] [Green Version]
- Plath, N.; Ohana, O.; Dammermann, B.; Errington, M.L.; Schmitz, D.; Gross, C.; Mao, X.; Engelsberg, A.; Mahlke, C.; Welzl, H.; et al. Arc/Arg3.1 is essential for the consolidation of synaptic plasticity and memories. Neuron 2006, 52, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Castro-Gomez, S.; Grendel, J.; Graf, S.; Susens, U.; Binkle, L.; Mensching, D.; Isbrandt, D.; Kuhl, D.; Ohana, O. Arc/Arg3.1 mediates a critical period for spatial learning and hippocampal networks. Proc. Natl. Acad. Sci. USA 2018, 115, 12531–12536. [Google Scholar] [CrossRef] [Green Version]
- Benito, E.; Barco, A. CREB’s control of intrinsic and synaptic plasticity: Implications for CREB-dependent memory models. Trends Neurosci. 2010, 33, 230–240. [Google Scholar] [CrossRef] [Green Version]
- Takeo, S.; Niimura, M.; Miyake-Takagi, K.; Nagakura, A.; Fukatsu, T.; Ando, T.; Takagi, N.; Tanonaka, K.; Hara, J. A possible mechanism for improvement by a cognition-enhancer nefiracetam of spatial memory function and cAMP-mediated signal transduction system in sustained cerebral ischaemia in rats. Br. J. Pharmacol. 2003, 138, 642–654. [Google Scholar] [CrossRef] [Green Version]
- Landeira, B.S.; Santana, T.; Araujo, J.A.M.; Tabet, E.I.; Tannous, B.A.; Schroeder, T.; Costa, M.R. Activity-Independent Effects of CREB on Neuronal Survival and Differentiation during Mouse Cerebral Cortex Development. Cereb. Cortex 2018, 28, 538–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakamoto, K.; Karelina, K.; Obrietan, K. CREB: A multifaceted regulator of neuronal plasticity and protection. J. Neurochem. 2011, 116, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Stewart, M.; Lau, P.; Banks, G.; Bains, R.S.; Castroflorio, E.; Oliver, P.L.; Dixon, C.L.; Kruer, M.C.; Kullmann, D.M.; Acevedo-Arozena, A.; et al. Loss of Frrs1l disrupts synaptic AMPA receptor function, and results in neurodevelopmental, motor, cognitive and electrographical abnormalities. Dis. Model. Mech. 2019, 12, dmm036806. [Google Scholar] [CrossRef] [Green Version]
- Palmer, C.L.; Cotton, L.; Henley, J.M. The molecular pharmacology and cell biology of alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors. Pharmacol. Rev. 2005, 57, 253–277. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Watson, J.F.; Matthews, P.M.; Cais, O.; Greger, I.H. Gating and modulation of a hetero-octameric AMPA glutamate receptor. Nature 2021, 594, 454–458. [Google Scholar] [CrossRef]
- Diering, G.H.; Huganir, R.L. The AMPA Receptor Code of Synaptic Plasticity. Neuron 2018, 100, 314–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.H.; Seo, S.Y.; Oh, J.H.; Ryu, I.S.; Kim, J.; Lee, D.K.; Ryu, Y.; Choe, E.S. Activation of Protein Kinase G After Repeated Cocaine Administration Is Necessary for the Phosphorylation of alpha-Amino-3-Hydroxy-5-Methyl-4-Isoxazolepropionic Acid Receptor GluA1 at Serine 831 in the Rat Nucleus Accumbens. Front. Mol. Neurosci. 2018, 11, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meilin, S.; Machicao, F.; Elmlinger, M. Treatment with Actovegin improves spatial learning and memory in rats following transient forebrain ischaemia. J. Cell. Mol. Med. 2014, 18, 1623–1630. [Google Scholar] [CrossRef] [Green Version]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [Green Version]
- Qiu, L.L.; Pan, W.; Luo, D.; Zhang, G.F.; Zhou, Z.Q.; Sun, X.Y.; Yang, J.J.; Ji, M.H. Dysregulation of BDNF/TrkB signaling mediated by NMDAR/Ca(2+)/calpain might contribute to postoperative cognitive dysfunction in aging mice. J. Neuroinflam. 2020, 17, 23. [Google Scholar] [CrossRef] [Green Version]
- Ghafouri, S.; Fathollahi, Y.; Javan, M.; Shojaei, A.; Asgari, A.; Mirnajafi-Zadeh, J. Effect of low frequency stimulation on impaired spontaneous alternation behavior of kindled rats in Y-maze test. Epilepsy Res. 2016, 126, 37–44. [Google Scholar] [CrossRef]
- Neves, G.; Cooke, S.F.; Bliss, T.V.P. Synaptic plasticity, memory and the hippocampus: A neural network approach to causality. Nat. Rev. Neurosci. 2008, 9, 65–75. [Google Scholar] [CrossRef]
- Zenke, F.; Agnes, E.J.; Gerstner, W. Diverse synaptic plasticity mechanisms orchestrated to form and retrieve memories in spiking neural networks. Nat. Commun. 2015, 6, 6922. [Google Scholar] [CrossRef] [PubMed]
- Vutskits, L.; Xie, Z. Lasting impact of general anaesthesia on the brain: Mechanisms and relevance. Nat. Rev. Neurosci. 2016, 17, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Alam, A.; Hana, Z.; Jin, Z.; Suen, K.C.; Ma, D. Surgery, neuroinflammation and cognitive impairment. EBioMedicine 2018, 37, 547–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Li, Z.; Cao, Y.; Fan, D.; Chui, D.; Guo, X. Increased extrasynaptic GluN2B expression is involved in cognitive impairment after isoflurane anesthesia. Exp. Ther. Med. 2016, 12, 161–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colbran, R.J. Thematic Minireview Series: Molecular Mechanisms of Synaptic Plasticity. J. Biol. Chem. 2015, 290, 28594–28595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; He, Z.; Lv, H.; Chen, W.; Chen, J. Calpain-2 plays a pivotal role in the inhibitory effects of propofol against TNF-alpha-induced autophagy in mouse hippocampal neurons. J. Cell. Mol. Med. 2020, 24, 9287–9299. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, Q.X.; Hou, Y.Y.; Lu, B.; Yu, C.; Chen, J.; Ling, Q.L.; Cao, J.; Chi, Z.Q.; Xu, L.; et al. Actin polymerization-dependent increase in synaptic Arc/Arg3.1 expression in the amygdala is crucial for the expression of aversive memory associated with drug withdrawal. J. Neurosci. 2012, 32, 12005–12017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bramham, C.R.; Alme, M.N.; Bittins, M.; Kuipers, S.D.; Nair, R.R.; Pai, B.; Panja, D.; Schubert, M.; Soule, J.; Tiron, A.; et al. The Arc of synaptic memory. Exp. Brain Res. 2010, 200, 125–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchida, S.; Teubner, B.J.W.; Hevi, C.; Hara, K.; Kobayashi, A.; Dave, R.M.; Shintaku, T.; Jaikhan, P.; Yamagata, H.; Suzuki, T.; et al. CRTC1 Nuclear Translocation Following Learning Modulates Memory Strength via Exchange of Chromatin Remodeling Complexes on the Fgf1 Gene. Cell Rep. 2017, 18, 352–366. [Google Scholar] [CrossRef]
- Shi, M.; Ding, J.; Li, L.; Bai, H.; Li, X.; Lan, L.; Fan, H.; Gao, L. Effects of Ketamine on Learning and Memory in the Hippocampus of Rats through ERK, CREB, and Arc. Brain Sci. 2020, 11, 27. [Google Scholar] [CrossRef]
- Lv, X.F.; Sun, L.L.; Cui, C.L.; Han, J.S. NAc Shell Arc/Arg3.1 Protein Mediates Reconsolidation of Morphine CPP by Increased GluR1 Cell Surface Expression: Activation of ERK-Coupled CREB is Required. Int. J. Neuropsychopharmacol. 2015, 18, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepherd, J.D.; Rumbaugh, G.; Wu, J.; Chowdhury, S.; Plath, N.; Kuhl, D.; Huganir, R.L.; Worley, P.F. Arc/Arg3.1 mediates homeostatic synaptic scaling of AMPA receptors. Neuron 2006, 52, 475–484. [Google Scholar] [CrossRef] [Green Version]
- Roth, R.H.; Cudmore, R.H.; Tan, H.L.; Hong, I.; Zhang, Y.; Huganir, R.L. Cortical Synaptic AMPA Receptor Plasticity during Motor Learning. Neuron 2020, 105, 895–908.e5. [Google Scholar] [CrossRef]
- Li, Y.; Pehrson, A.L.; Waller, J.A.; Dale, E.; Sanchez, C.; Gulinello, M. A critical evaluation of the activity-regulated cytoskeleton-associated protein (Arc/Arg3.1)’s putative role in regulating dendritic plasticity, cognitive processes, and mood in animal models of depression. Front. Neurosci. 2015, 9, 279. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, G.; Li, T.; Huang, Y. The Effects of Intraoperative Hypothermia on Postoperative Cognitive Function in the Rat Hippocampus and Its Possible Mechanisms. Brain Sci. 2022, 12, 96. https://doi.org/10.3390/brainsci12010096
Xu G, Li T, Huang Y. The Effects of Intraoperative Hypothermia on Postoperative Cognitive Function in the Rat Hippocampus and Its Possible Mechanisms. Brain Sciences. 2022; 12(1):96. https://doi.org/10.3390/brainsci12010096
Chicago/Turabian StyleXu, Guangyan, Tianjia Li, and Yuguang Huang. 2022. "The Effects of Intraoperative Hypothermia on Postoperative Cognitive Function in the Rat Hippocampus and Its Possible Mechanisms" Brain Sciences 12, no. 1: 96. https://doi.org/10.3390/brainsci12010096
APA StyleXu, G., Li, T., & Huang, Y. (2022). The Effects of Intraoperative Hypothermia on Postoperative Cognitive Function in the Rat Hippocampus and Its Possible Mechanisms. Brain Sciences, 12(1), 96. https://doi.org/10.3390/brainsci12010096