
The Semantics of Natural Objects and Tools in the Brain: A Combined Behavioral and MEG Study

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment 1—Behavioral Study
2.1.1. Participants
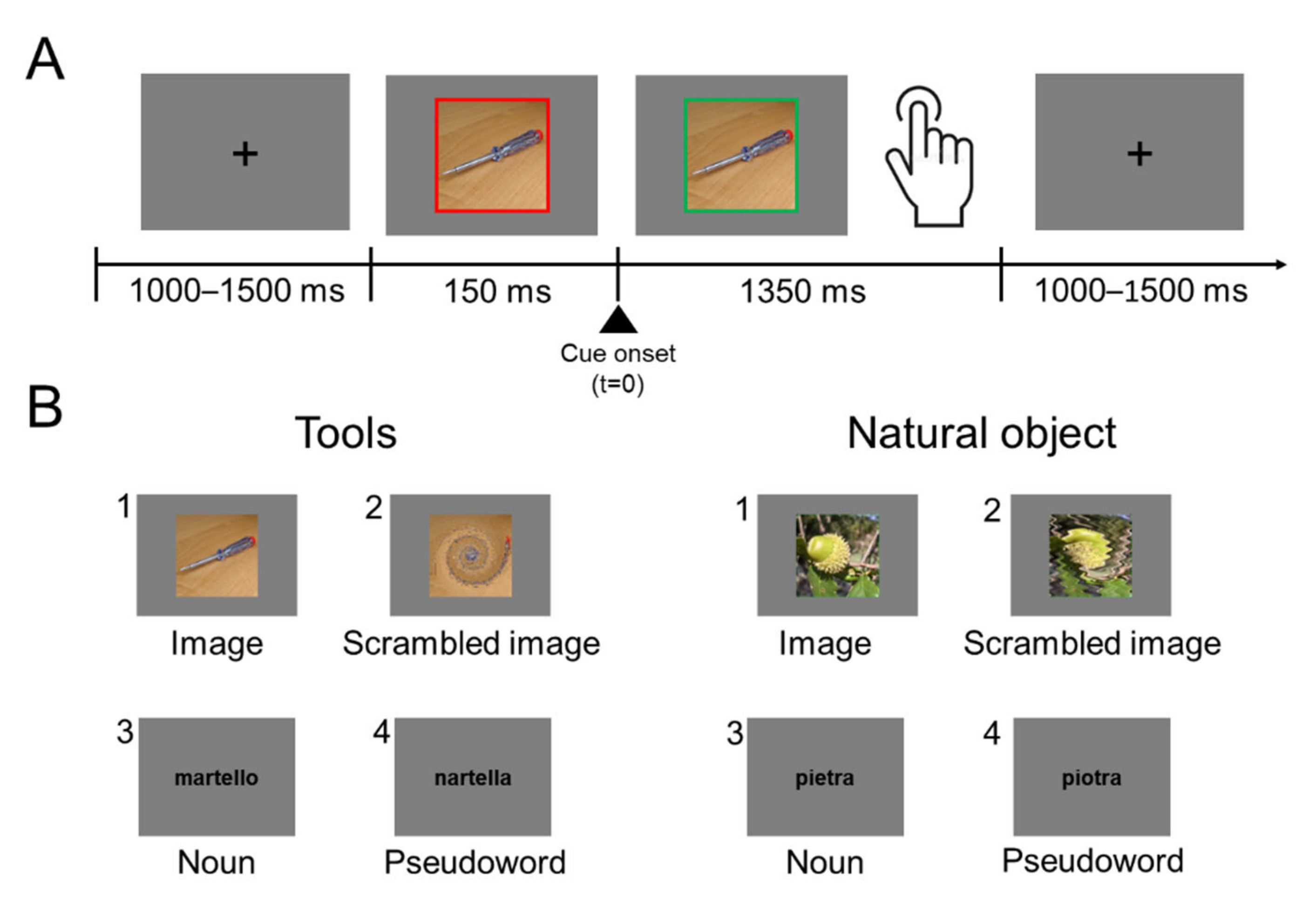
2.1.2. Apparatus, Procedure and Stimuli
2.1.3. Data Analysis
2.2. Experiment 2—MEG Study
2.2.1. Participants
2.2.2. Task
2.2.3. MEG Data Acquisition and Pre-Processing
2.3. Data Analysis
2.3.1. Sensors Analysis
2.3.2. Source Analysis
2.3.3. Statistical Analysis
3. Results
3.1. Experiment 1—Behavioral Study
3.2. Experiment 2—MEG Study
3.2.1. Behavioral Data
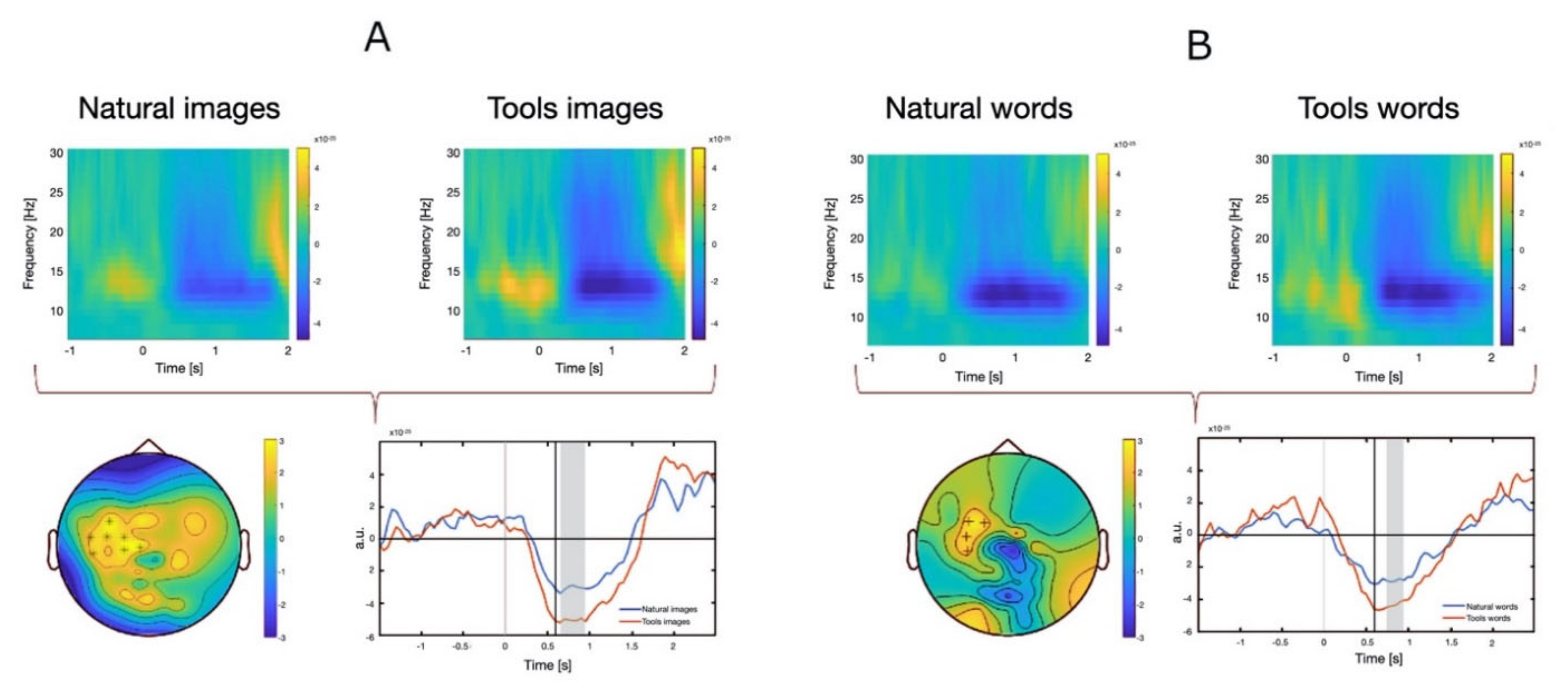
3.2.2. MEG Data
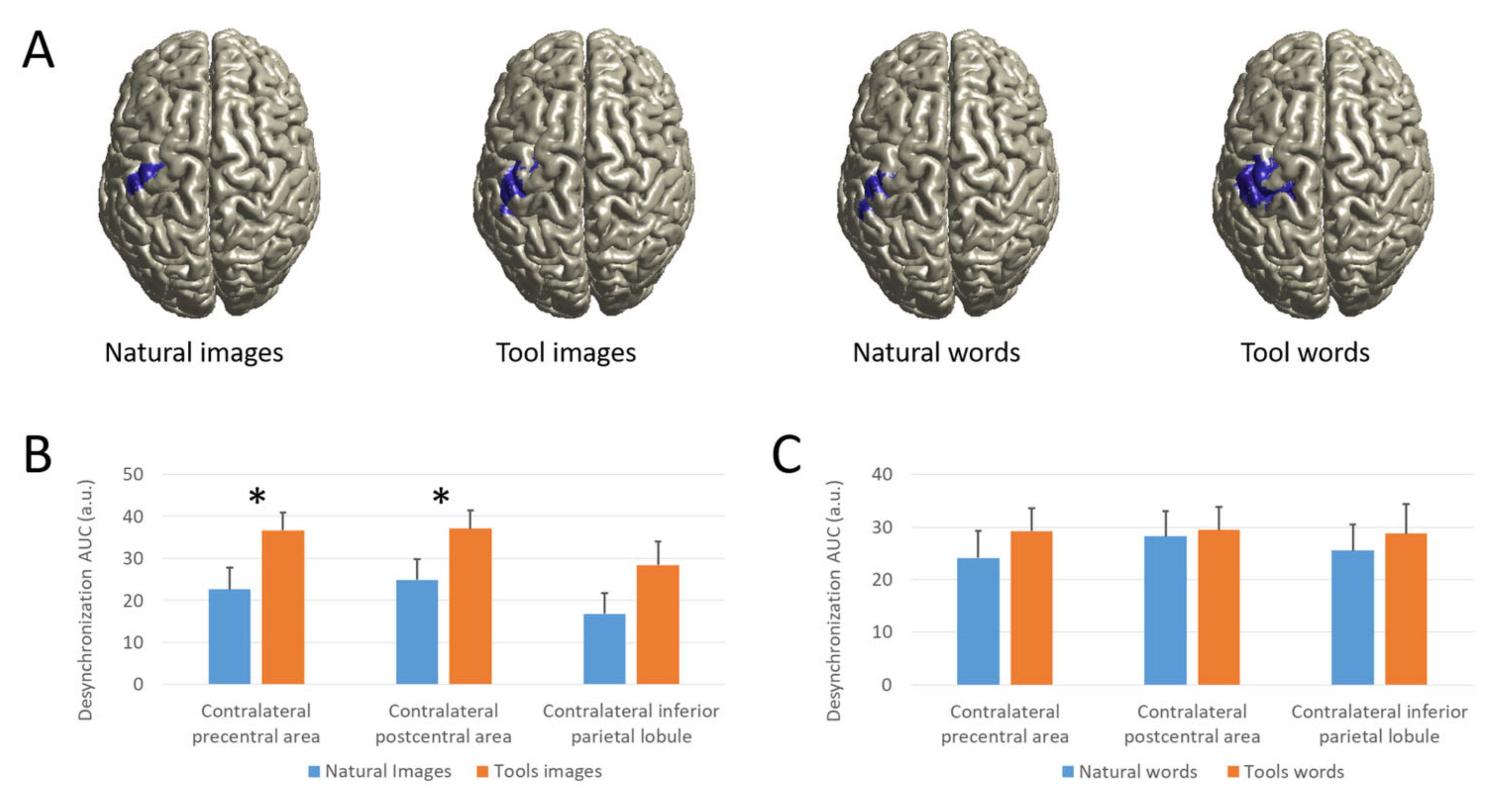
3.2.3. Source Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barsalou, L.W. Grounded cognition. Annu. Rev. Psychol. 2008, 59, 617–645. [Google Scholar] [CrossRef] [Green Version]
- Buccino, G.; Colagè, I.; Gobbi, N.; Bonaccorso, G. Grounding meaning in experience: A broad perspective on embodied language. Neurosci. Biobehav. Rev. 2016, 69, 69–78. [Google Scholar] [CrossRef]
- Fischer, M.H.; Zwaan, R.A. Embodied language: A review of the role of the motor system in language comprehension. Q. J. Exp. Psychol. 2008, 61, 825–850. [Google Scholar] [CrossRef] [PubMed]
- Gallese, V. Mirror neurons and the social nature of language: The neural exploitation hypothesis. Soc. Neurosci. 2008, 3, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Kousta, S.T.; Vigliocco, G.; Vinson, D.P.; Andrews, M.; Del Campo, E. The Representation of Abstract Words: Why Emotion Matters. J. Exp. Psychol. Gen. 2011, 140, 14–34. [Google Scholar] [CrossRef] [PubMed]
- Pulvermüller, F. A brain perspective on language mechanisms: From discrete neuronal ensembles to serial order. Prog. Neurobiol. 2002, 67, 85–111. [Google Scholar] [CrossRef]
- Vigliocco, G.; Kousta, S.T.; Della Rosa, P.A.; Vinson, D.P.; Tettamanti, M.; Devlin, J.T.; Cappa, S.F. The Neural Representation of Abstract Words: The Role of Emotion. Cereb. Cortex 2014, 24, 1767–1777. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, A. Disembodying cognition. Lang. Cogn. 2011, 2, 79–116. [Google Scholar] [CrossRef] [Green Version]
- Mahon, B.Z.; Caramazza, A. The orchestration of the sensory-motor systems: Clues from neuropsychology. Cogn. Neuropsychol. 2005, 22, 480–494. [Google Scholar] [CrossRef]
- Mahon, B.Z.; Caramazza, A. A critical look at the embodied cognition hypothesis and a new proposal for grounding conceptual content. J. Physiol. Paris 2008, 102, 59–70. [Google Scholar] [CrossRef]
- Pylyshyn, Z. Return of the mental image: Are there really pictures in the brain? Trends Cogn. Sci. 2003, 7, 113–118. [Google Scholar] [CrossRef]
- Desai, R.H.; Binder, J.R.; Conant, L.L.; Seidenberg, M.S. Activation of Sensory–Motor Areas in Sentence Comprehension. Cereb. Cortex 2010, 20, 468–478. [Google Scholar] [CrossRef]
- Fernandino, L.; Binder, J.R.; Desai, R.H.; Pendl, S.L.; Humphries, C.J.; Gross, W.L.; Conant, L.L.; Seidenberg, M.S. Concept Representation Reflects Multimodal Abstraction: A Framework for Embodied Semantics. Cereb. Cortex 2016, 26, 2018–2034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahon, B.Z.; Kemmerer, D. Interactions between language, thought, and perception: Cognitive and neural perspectives. Cogn. Neuropsychol. 2020, 37, 235–240. [Google Scholar] [CrossRef]
- Martin, A. The representation of object concepts in the brain. Annu. Rev. Psychol. 2007, 58, 25–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, A. GRAPES-Grounding representations in action, perception, and emotion systems: How object properties and categories are represented in the human brain. Psychon. Bull. Rev. 2016, 23, 979–990. [Google Scholar] [CrossRef]
- González, J.; Barros-Loscertales, A.; Pulvermüller, F.; Meseguer, V.; Sanjuán, A.; Belloch, V.; Ávila, C. Reading cinnamon activates olfactory brain regions. NeuroImage 2006, 32, 906–912. [Google Scholar] [CrossRef] [PubMed]
- Barrós-Loscertales, A.; González, J.; Pulvermüller, F.; Ventura-Campos, N.; Bustamante, J.C.; Costumero, V.; Parcet, M.A.; Ávila, C. Reading salt activates gustatory brain regions: FMRI evidence for semantic grounding in a novel sensory modality. Cereb. Cortex 2012, 22, 2554–2563. [Google Scholar] [CrossRef] [PubMed]
- Ponz, A.; Montant, M.; Liegeois-Chauvel, C.; Silva, C.; Braun, M.; Jacobs, A.M.; Ziegler, J.C. Emotion processing in words: A test of the neural re-use hypothesis using surface and intracranial EEG. Soc. Cogn. Affect. Neurosci. 2014, 9, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Mahon, B.Z. What is embodied about cognition? Lang. Cogn. Neurosci. 2015, 30, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Binder, J.R.; Desai, R.H.; Graves, W.W.; Conant, L.L. Where is the semantic system? A critical review and meta-analysis of 120 functional neuroimaging studies. Cereb. Cortex 2009, 19, 2767–2796. [Google Scholar] [CrossRef]
- Goodale, M.A.; Milner, A.D. Separate visual pathways for perception and action. Trends Neurosci. 1992, 15, 20–25. [Google Scholar] [CrossRef]
- Goodale, M.A.; Milner, A.D. Two visual streams: Interconnections do not imply duplication of function. Cogn. Neurosci. 2010, 1, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Milner, A.D.; Goodale, M.A. Two visual systems re-viewed. Neuropsychologia 2008, 46, 774–785. [Google Scholar] [CrossRef] [PubMed]
- Buccino, G.; Sato, M.; Cattaneo, L.; Rodà, F.; Riggio, L. Broken affordances, broken objects: A TMS study. Neuropsychologia 2009, 47, 3074–3078. [Google Scholar] [CrossRef]
- Chao, L.L.; Martin, A. Representation of manipulable man-made objects in the dorsal stream. NeuroImage 2000, 12, 478–484. [Google Scholar] [CrossRef] [Green Version]
- Rizzolatti, G.; Matelli, M. Two different streams form the dorsal visual system: Anatomy and functions. Exp. Brain Res. 2003, 153, 146–157. [Google Scholar] [CrossRef]
- Binkofski, F.; Buxbaum, L.J. Two action systems in the human brain. Brain Lang. 2013, 127, 222–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakreida, K.; Effnert, I.; Thill, S.; Menz, M.M.; Jirak, D.; Eickhoff, C.R.; Ziemke, T.; Eickhoff, S.B.; Borghi, A.M.; Binkofski, F. Affordance processing in segregated parieto-frontal dorsal stream sub-pathways. Neurosci. Biobehav. Rev. 2016, 69, 89–112. [Google Scholar] [CrossRef]
- Binkofski, F.; Buccino, G. The role of the parietal cortex in sensorimotor transformations and action coding. Handb. Clin. Neurol. 2018, 151, 467–479. [Google Scholar]
- Gonzalez Rothi, L.J.; Ochipa, C.; Heilman, K.M. A Cognitive Neuropsychological Model of Limb Praxis. Cogn. Neuropsychol. 1991, 8, 443–458. [Google Scholar] [CrossRef]
- Hodges, J.R.; Spatt, J.; Patterson, K. “What” and “how”: Evidence for the dissociation of object knowledge and mechanical problem-solving skills in the human brain. Proc. Natl. Acad. Sci. USA 1999, 96, 9444–9448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson-Frey, S.H. The neural bases of complex tool use in humans. Trends Cogn. Sci. 2004, 8, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Kalénine, S.; Buxbaum, L.J.; Coslett, H.B. Critical brain regions for action recognition: Lesion symptom mapping in left hemisphere stroke. Brain 2010, 133, 3269–3280. [Google Scholar] [CrossRef]
- Negri, G.A.L.; Rumiati, R.; Zadini, A.; Ukmar, M.; Mahon, B.; Caramazza, A. What is the role of motor simulation in action and object recognition? Evidence from apraxia. Cogn. Neuropsychol. 2007, 24, 795–816. [Google Scholar] [CrossRef] [PubMed]
- Cohen, N.R.; Cross, E.S.; Tunik, E.; Grafton, S.T.; Culham, J.C. Ventral and dorsal stream contributions to the online control of immediate and delayed grasping: A TMS approach. Neuropsychologia 2009, 47, 1553–1562. [Google Scholar] [CrossRef]
- Whitwell, R.L.; Milner, A.D.; Goodale, M.A. The two visual systems hypothesis: New challenges and insights from visual form agnosic patient DF. Front. Neurol. 2014, 5, 255. [Google Scholar] [CrossRef] [Green Version]
- Van Polanen, V.; Davare, M. Interactions between dorsal and ventral streams for controlling skilled grasp. Neuropsychologia 2015, 79, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Kopiske, K.K.; Bruno, N.; Hesse, C.; Schenk, T.; Franz, V.H. The functional subdivision of the visual brain: Is there a real illusion effect on action? A multi-lab replication study. Cortex 2016, 79, 130–152. [Google Scholar] [CrossRef] [Green Version]
- Uccelli, S.; Pisu, V.; Riggio, L.; Bruno, N. The Uznadze illusion reveals similar effects of relative size on perception and action. Exp. Brain Res. 2019, 237, 953–965. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, G.; Riggio, L. Influence of colour on object motor representation. Neuropsychologia 2022, 164, 108103. [Google Scholar] [CrossRef]
- Boronat, C.B.; Buxbaum, L.J.; Coslett, H.B.; Tang, K.; Saffran, E.M.; Kimberg, D.Y.; Detre, J.A. Distinctions between manipulation and function knowledge of objects: Evidence from functional magnetic resonance imaging. Cogn. Brain Res. 2005, 23, 361–373. [Google Scholar] [CrossRef]
- Brandi, M.L.; Wohlschläger, A.; Sorg, C.; Hermsdörfer, J. The Neural Correlates of Planning and Executing Actual Tool Use. J. Neurosci. 2014, 34, 13183–13194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Garcea, F.E.; Mahon, B.Z. The Representation of Object-Directed Action and Function Knowledge in the Human Brain. Cereb. Cortex 2016, 26, 1609–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Garcea, F.E.; Jacobs, R.A.; Mahon, B.Z. Abstract Representations of Object-Directed Action in the Left Inferior Parietal Lobule. Cereb. Cortex 2018, 28, 2162–2174. [Google Scholar] [CrossRef]
- Gallivan, J.P.; Adam McLean, D.; Valyear, K.F.; Culham, J.C. Decoding the neural mechanisms of human tool use. ELife 2013, 2, e00425. [Google Scholar] [CrossRef]
- Hermsdörfer, J.; Terlinden, G.; Mühlau, M.; Goldenberg, G.; Wohlschläger, A.M. Neural representations of pantomimed and actual tool use: Evidence from an event-related fMRI study. NeuroImage 2007, 36, T109–T118. [Google Scholar] [CrossRef] [PubMed]
- Kellenbach, M.L.; Brett, M.; Patterson, K. Actions Speak Louder Than Functions: The Importance of Manipulability and Action in Tool Representation. J. Cogn. Neurosci. 2003, 15, 30–46. [Google Scholar] [CrossRef]
- Marques, J.F.; Canessa, N.; Cappa, S. Neural differences in the processing of true and false sentences: Insights into the nature of “truth” in language comprehension. Cortex 2009, 45, 759–768. [Google Scholar] [CrossRef]
- Rumiati, R.I.; Weiss, P.H.; Shallice, T.; Ottoboni, G.; Noth, J.; Zilles, K.; Fink, G.R. Neural basis of pantomiming the use of visually presented objects. NeuroImage 2004, 21, 1224–1231. [Google Scholar] [CrossRef]
- Buchwald, M.; Przybylski, L.; Króliczak, G. Decoding Brain States for Planning Functional Grasps of Tools: A Functional Magnetic Resonance Imaging Multivoxel Pattern Analysis Study. J. Int. Neuropsychol. Soc. 2018, 24, 1013–1025. [Google Scholar] [CrossRef]
- Grafton, S.T.; Fadiga, L.; Arbib, M.A.; Rizzolatti, G. Premotor cortex activation during observation and naming of familiar tools. NeuroImage 1997, 6, 231–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Króliczak, G.; Frey, S.H. A Common Network in the Left Cerebral Hemisphere Represents Planning of Tool Use Pantomimes and Familiar Intransitive Gestures at the Hand-Independent Level. Cereb. Cortex 2009, 19, 2396–2410. [Google Scholar] [CrossRef] [Green Version]
- Peeters, R.; Simone, L.; Nelissen, K.; Fabbri-Destro, M.; Vanduffel, W.; Rizzolatti, G.; Orban, G.A. The Representation of Tool Use in Humans and Monkeys: Common and Uniquely Human Features. J. Neurosci. 2009, 29, 11523–11539. [Google Scholar] [CrossRef] [Green Version]
- Vigliocco, G.; Vinson, D.P.; Druks, J.; Barber, H.; Cappa, S.F. Nouns and verbs in the brain: A review of behavioural, electrophysiological, neuropsychological and imaging studies. Neurosci. Biobehav. Rev. 2011, 35, 407–426. [Google Scholar] [CrossRef] [PubMed]
- Marino, B.F.M.; Sirianni, M.; Volta, R.D.; Magliocco, F.; Silipo, F.; Quattrone, A.; Buccino, G. Viewing photos and reading nouns of natural graspable objects similarly modulate motor responses. Front. Hum. Neurosci. 2014, 8, 968. [Google Scholar] [CrossRef] [PubMed]
- Buccino, G.; Marino, B.F.; Bulgarelli, C.; Mezzadri, M. Fluent speakers of a second language process graspable nouns expressed in L2 like in their native language. Front. Psychol. 2017, 8, 1306. [Google Scholar] [CrossRef] [Green Version]
- Buccino, G.; Dalla Volta, R.; Arabia, G.; Morelli, M.; Chiriaco, C.; Lupo, A.; Silipo, F.; Quattrone, A. Processing graspable object images and their nouns is impaired in Parkinson’s disease patients. Cortex 2018, 100, 32–39. [Google Scholar] [CrossRef]
- Zhang, Z.; Sun, Y.; Humphreys, G.W. Perceiving object affordances through visual and linguistic pathways: A comparative study. Sci. Rep. 2016, 6, 26806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bub, D.N.; Masson, M.E.J.; Kumar, R. Time course of motor affordances evoked by pictured objects and words. J. Exp. Psychol. Hum. Percept. Perform. 2018, 44, 53–68. [Google Scholar] [CrossRef]
- Horoufchin, H.; Bzdok, D.; Buccino, G.; Borghi, A.M.; Binkofski, F. Action and object words are differentially anchored in the sensory motor system—A perspective on cognitive embodiment. Sci. Rep. 2018, 8, 6583. [Google Scholar] [CrossRef]
- Harpaintner, M.; Sim, E.J.; Trumpp, N.M.; Ulrich, M.; Kiefer, M. The grounding of abstract concepts in the motor and visual system: An fMRI study. Cortex 2020, 124, 1–22. [Google Scholar] [CrossRef]
- Klepp, A.; Niccolai, V.; Buccino, G.; Schnitzler, A.; Biermann-Ruben, K. Language-motor interference reflected in MEG beta oscillations. NeuroImage 2015, 109, 438–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulenger, V.; Roy, A.C.; Paulignan, Y.; Deprez, V.; Jeannerod, M.; Nazir, T.A. Cross-talk between language processes and overt motor behavior in the first 200 msec of processing. J. Cogn. Neurosci. 2006, 18, 1607–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buccino, G.; Riggio, L.; Melli, G.; Binkofski, F.; Gallese, V.; Rizzolatti, G. Listening to action-related sentences modulates the activity of the motor system: A combined TMS and behavioral study. Cogn. Brain Res. 2005, 24, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Dalla Volta, R.; Gianelli, C.; Campione, G.C.; Gentilucci, M. Action word understanding and overt motor behavior. Exp. Brain Res. 2009, 196, 403–412. [Google Scholar] [CrossRef] [PubMed]
- De Vega, M.; Moreno, V.; Castillo, D. The comprehension of action-related sentences may cause interference rather than facilitation on matching actions. Psychol. Res. 2013, 77, 20–30. [Google Scholar] [CrossRef]
- De Vega, M.; León, I.; Hernández, J.A.; Valdés, M.; Padrón, I.; Ferstl, E.C. Action Sentences Activate Sensory Motor Regions in the Brain Independently of Their Status of Reality. J. Cogn. Neurosci. 2014, 26, 1363–1376. [Google Scholar] [CrossRef]
- Niccolai, V.; Klepp, A.; Indefrey, P.; Schnitzler, A.; Biermann-Ruben, K. Semantic discrimination impacts tDCS modulation of verb processing. Sci. Rep. 2017, 7, 17162. [Google Scholar] [CrossRef] [Green Version]
- Pulvermüller, F.; Assadollahi, R.; Elbert, T. Neuromagnetic evidence for early semantic access in word recognition. Eur. J. Neurosci. 2001, 13, 201–205. [Google Scholar] [CrossRef]
- Sato, M.; Mengarelli, M.; Riggio, L.; Gallese, V.; Buccino, G. Task related modulation of the motor system during language processing. Brain Lang. 2008, 105, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Pfurtscheller, G.; Lopes Da Silva, F.H. Event-related EEG/MEG synchronization and desynchronization: Basic principles. Clin. Neurophysiol. 1999, 110, 1842–1857. [Google Scholar] [CrossRef]
- Hari, R.; Kujala, M.V. Brain basis of human social interaction: From concepts to brain imaging. Physiol. Rev. 2009, 89, 453–479. [Google Scholar] [CrossRef] [Green Version]
- Moreno, I.; de Vega, M.; León, I. Understanding action language modulates oscillatory mu and beta rhythms in the same way as observing actions. Brain Cogn. 2013, 82, 236–242. [Google Scholar] [CrossRef]
- Brinkman, L.; Stolk, A.; Dijkerman, H.C.; de Lange, F.P.; Toni, I. Distinct Roles for Alpha- and Beta-Band Oscillations during Mental Simulation of Goal-Directed Actions. J. Neurosci. 2014, 34, 14783–14792. [Google Scholar] [CrossRef]
- De Lange, F.P.; Roelofs, K.; Toni, I. Motor imagery: A window into the mechanisms and alterations of the motor system. Cortex 2008, 44, 494–506. [Google Scholar] [CrossRef] [Green Version]
- Schnitzler, A.; Salenius, S.; Salmelin, R.; Jousmäki, V.; Hari, R. Involvement of Primary Motor Cortex in Motor Imagery: A Neuromagnetic Study. NeuroImage 1997, 6, 201–208. [Google Scholar] [CrossRef]
- Weiss, S.; Mueller, H. “Too Many betas do not Spoil the Broth”: The Role of Beta Brain Oscillations in Language Processing. Front. Psychol. 2012, 3, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing (3.6.3); R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Taulu, S.; Simola, J. Spatiotemporal signal space separation method for rejecting nearby interference in MEG measurements. Phys. Med. Biol. 2006, 51, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Gross, J.; Kujala, J.; Hamalainen, M.; Timmermann, L.; Schnitzler, A.; Salmelin, R. Dynamic imaging of coherent sources: Studying neural interactions in the human brain. Proc. Natl. Acad. Sci. USA 2001, 98, 694–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Veen, B.D.; van Drongelen, W.; Yuchtman, M.; Suzuki, A. Localization of brain electrical activity via linearly constrained minimum variance spatial filtering. IEEE Trans. Biomed. Eng. 1997, 44, 867–880. [Google Scholar] [CrossRef]
- Friston, K.J.; Holmes, A.P.; Worsley, K.J.; Poline, J.P.; Frith, C.D.; Frackowiak, R.S.J. Statistical parametric maps in functional imaging: A general linear approach. Hum. Brain Mapp. 1994, 2, 189–210. [Google Scholar] [CrossRef]
- Oostenveld, R.; Fries, P.; Maris, E.; Schoffelen, J.M. FieldTrip: Open Source Software for Advanced Analysis of MEG, EEG, and Invasive Electrophysiological Data. Comput. Intell. Neurosci. 2011, 2011, 156869. [Google Scholar] [CrossRef]
- Hari, R.; Forss, N.; Avikainen, S.; Kirveskari, E.; Salenius, S.; Rizzolatti, G. Activation of human primary motor cortex during action observation: A neuromagnetic study. Proc. Natl. Acad. Sci. USA 1998, 95, 15061–15065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devereux, B.J.; Clarke, A.; Marouchos, A.; Tyle, L.K. Representational similarity analysis reveals commonalities and differences in the semantic processing of words and objects. J. Neurosci. 2013, 33, 18906–18916. [Google Scholar] [CrossRef] [Green Version]
- Shinkareva, S.V.; Malave, V.L.; Mason, R.A.; Mitchell, T.M.; Just, M.A. Commonality of neural representations of words and pictures. NeuroImage 2011, 54, 2418–2425. [Google Scholar] [CrossRef]
- Simanova, I.; Hagoort, P.; Oostenveld, R.; Van Gerven, M.A.J. Modality-independent decoding of semantic information from the human brain. Cereb. Cortex 2014, 24, 426–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganis, G.; Kutas, M.; Sereno, M.I. The Search for “Common Sense”: An Electrophysiological Study of the Comprehension of Words and Pictures in Reading. J. Cogn. Neurosci. 1996, 8, 89–106. [Google Scholar] [CrossRef] [PubMed]
- Van Doren, L.; Dupont, P.; De Grauwe, S.; Peeters, R.; Vandenberghe, R. The amodal system for conscious word and picture identification in the absence of a semantic task. NeuroImage 2010, 49, 3295–3307. [Google Scholar] [CrossRef]
- Vandenberghe, R.; Price, C.; Wise, R.; Josephs, O.; Frackowiak, R.S.J. Functional anatomy of a common semantic system for words and pictures. Nature 1996, 383, 254–256. [Google Scholar] [CrossRef] [Green Version]
- Garofalo, G.; Magliocco, F.; Silipo, F.; Riggio, L.; Buccino, G. What matters is the undelying experiences similar motor responses during processing observed hand actions and hand related verbs. J. Neuropsychol. 2021; In press. [Google Scholar]
- Santana, E.J.; De Vega, M. An ERP study of motor compatibility effects in action language. Brain Res. 2013, 1526, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Chersi, F.; Thill, S.; Ziemke, T.; Borghi, A.M. Sentence processing: Linking language to motor chains. Front. Neurorobotics 2010, 4, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, A.M.; Ibáñez, A. A touch with words: Dynamic synergies between manual actions and language. Neurosci. Biobehav. Rev. 2016, 68, 59–95. [Google Scholar] [CrossRef] [PubMed]
- Marino, B.F.M.; Gough, P.M.; Gallese, V.; Riggio, L.; Buccino, G. How the motor system handles nouns: A behavioral study. Psychol. Res. 2013, 77, 64–73. [Google Scholar] [CrossRef]
- lriki, A.; Tanaka, M.; Iwamura, Y. Coding of modified body schema during tool use by macaque postcentral neurones. NeuroReport 1996, 7, 2325–2330. [Google Scholar] [CrossRef] [PubMed]
- Maravita, A.; Iriki, A. Tools for the body (schema). Trends Cogn. Sci. 2004, 8, 79–86. [Google Scholar] [CrossRef]
- Nelissen, K.; Vanduffel, W. Grasping-related functional MRI brain responses in the macaque monkey. J. Neurosci. 2011, 31, 8220–8229. [Google Scholar] [CrossRef] [Green Version]
- Errante, A.; Ziccarelli, S.; Mingolla, G.; Fogassi, L. Grasping and Manipulation: Neural Bases and Anatomical Circuitry in Humans. Neuroscience 2021, 458, 203–212. [Google Scholar] [CrossRef]
- Buxbaum, L.J.; Veramontil, T.; Schwartz, M.F. Function and manipulation tool knowledge in apraxia: Knowing ‘what for’ but not ‘how’. Neurocase 2000, 6, 83–97. [Google Scholar]
- De Renzi, E.; Lucchelli, F. Ideational Apraxia. Brain 1988, 111, 1173–1185. [Google Scholar] [CrossRef] [PubMed]
- Heilman, K.M.; Schwartz, H.D.; Geschwind, N. Defective motor learning in ideomotor apraxia. Neurology 1975, 25, L–1018. [Google Scholar] [CrossRef] [PubMed]
- Caminiti, R.; Chafee, M.V.; Battaglia-Mayer, A.; Averbeck, B.B.; Crowe, D.A.; Georgopoulos, A.P. Understanding the parietal lobe syndrome from a neurophysiological and evolutionary perspective. Eur. J. Neurosci. 2010, 31, 2320–2340. [Google Scholar] [CrossRef] [PubMed]
- Gough, P.M.; Riggio, L.; Chersi, F.; Sato, M.; Fogassi, L.; Buccino, G. Nouns referring to tools and natural objects differentially modulate the motor system. Neuropsychologia 2012, 50, 19–25. [Google Scholar] [CrossRef]
- Desai, R.H.; Herter, T.; Riccardi, N.; Rorden, C.; Fridriksson, J. Concepts within reach: Action performance predicts action language processing in stroke. Neuropsychologia 2015, 71, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Raichle, M.E.; MacLeod, A.M.; Snyder, A.Z.; Powers, W.J.; Gusnard, D.A.; Shulman, G.L. A default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raichle, M.E. The Brain’s Default Mode Network. Annu. Rev. Neurosci. 2015, 38, 433–447. [Google Scholar] [CrossRef] [Green Version]
- Mars, R.B.; Neubert, F.X.; Noonan, M.A.P.; Sallet, J.; Toni, I.; Rushworth, M.F.S. On the relationship between the “default mode network” and the “social brain”. Front. Hum. Neurosci. 2012, 6, 189. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, J.P.; Macrae, C.N.; Banaji, M.R. Dissociable Medial Prefrontal Contributions to Judgments of Similar and Dissimilar Others. Neuron 2006, 50, 655–663. [Google Scholar] [CrossRef] [Green Version]
- Wen, T.; Mitchell, D.J.; Duncan, J. The Functional Convergence and Heterogeneity of Social, Episodic, and Self-Referential Thought in the Default Mode Network. Cereb. Cortex 2020, 30, 5915–5929. [Google Scholar] [CrossRef] [PubMed]
- Goldman-Rakic, P.S. Regional and cellular fractionation of working memory. Proc. Natl. Acad. Sci. 1996, 93, 13473–13480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buccino, G.; Vogt, S.; Ritzl, A.; Fink, G.R.; Zilles, K.; Freund, H.J.; Rizzolatti, G. Neural circuits underlying imitation learning of hand actions: An event-related fMRI study. Neuron 2004, 42, 323–334. [Google Scholar] [CrossRef]
- Vogt, S.; Buccino, G.; Wohlschläger, A.M.; Canessa, N.; Shah, N.J.; Zilles, K.; Eickhoff, S.B.; Freund, H.J.; Rizzolatti, G.; Fink, G.R. Prefrontal involvement in imitation learning of hand actions: Effects of practice and expertise. NeuroImage 2007, 37, 1371–1383. [Google Scholar] [CrossRef] [Green Version]
- Di Tella, S.; Blasi, V.; Cabinio, M.; Bergsland, N.; Buccino, G.; Baglio, F. How Do We Motorically Resonate in Aging? A Compensatory Role of Prefrontal Cortex. Front. Aging Neurosci. 2021, 13, 412. [Google Scholar] [CrossRef]
- Binkofski, F.; Buccino, G. The role of ventral premotor cortex in action execution and action understanding. J. Physiol. Paris 2006, 99, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Petrides, M.; Cadoret, G.; Mackey, S. Orofacial somatomotor responses in the macaque monkey homologue of Broca’s area. Nature 2005, 435, 1235–1238. [Google Scholar] [CrossRef]
- Tettamanti, M.; Buccino, G.; Saccuman, M.C.; Gallese, V.; Danna, M.; Scifo, P.; Fazio, F.; Rizzolatti, G.; Cappa, S.F.; Perani, D. Listening to action-related sentences activates fronto-parietal motor circuits. J. Cogn. Neurosci. 2005, 17, 273–281. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Noun | Image | |||||
|---|---|---|---|---|---|---|
| Mean (ms) | Standard Deviation (ms) | Standard Error (ms) | Mean (ms) | Standard Deviation (ms) | Standard Error (ms) | |
| Natural | 708 | 80.91 | 15.57 | 720 | 94.61 | 18.21 |
| Tool | 666 | 76.03 | 14.63 | 686 | 93.27 | 17.95 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Visani, E.; Sebastiano, D.R.; Duran, D.; Garofalo, G.; Magliocco, F.; Silipo, F.; Buccino, G. The Semantics of Natural Objects and Tools in the Brain: A Combined Behavioral and MEG Study. Brain Sci. 2022, 12, 97. https://doi.org/10.3390/brainsci12010097
Visani E, Sebastiano DR, Duran D, Garofalo G, Magliocco F, Silipo F, Buccino G. The Semantics of Natural Objects and Tools in the Brain: A Combined Behavioral and MEG Study. Brain Sciences. 2022; 12(1):97. https://doi.org/10.3390/brainsci12010097
Chicago/Turabian StyleVisani, Elisa, Davide Rossi Sebastiano, Dunja Duran, Gioacchino Garofalo, Fabio Magliocco, Francesco Silipo, and Giovanni Buccino. 2022. "The Semantics of Natural Objects and Tools in the Brain: A Combined Behavioral and MEG Study" Brain Sciences 12, no. 1: 97. https://doi.org/10.3390/brainsci12010097
APA StyleVisani, E., Sebastiano, D. R., Duran, D., Garofalo, G., Magliocco, F., Silipo, F., & Buccino, G. (2022). The Semantics of Natural Objects and Tools in the Brain: A Combined Behavioral and MEG Study. Brain Sciences, 12(1), 97. https://doi.org/10.3390/brainsci12010097