
Scinderin Is a Novel Oncogene for Its Correlates with Poor Prognosis, Immune Infiltrates and Matrix Metalloproteinase-2/9 (MMP2/9) in Glioma
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Clinical Samples
2.2. Gene Expression Profiling Interactive Analysis (GEPIA)
2.3. Oncomine and cBioPortal
2.4. Chinese Glioma Genome Atlas (CGGA)
2.5. Gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG)
2.6. Protein–Protein Interaction (PPI) Network and GeneMANIA
2.7. Immune Infiltration Analysis
2.8. The Cancer Genome Atlas (TCGA)
2.9. Tissue Microarray and Immunohistochemistry (IHC)
2.10. Western Blot Assay
2.11. Statistical Analysis
3. Results
3.1. SCIN Is Significantly Upregulated in Glioma
3.2. SCIN Is A Prognostic Factor for Glioma Patients
3.3. Functional Enrichment Analysis of SCIN-Related Genes in Glioma
3.4. SCIN Expression Is Associated Immune Infiltration in Glioma Cells
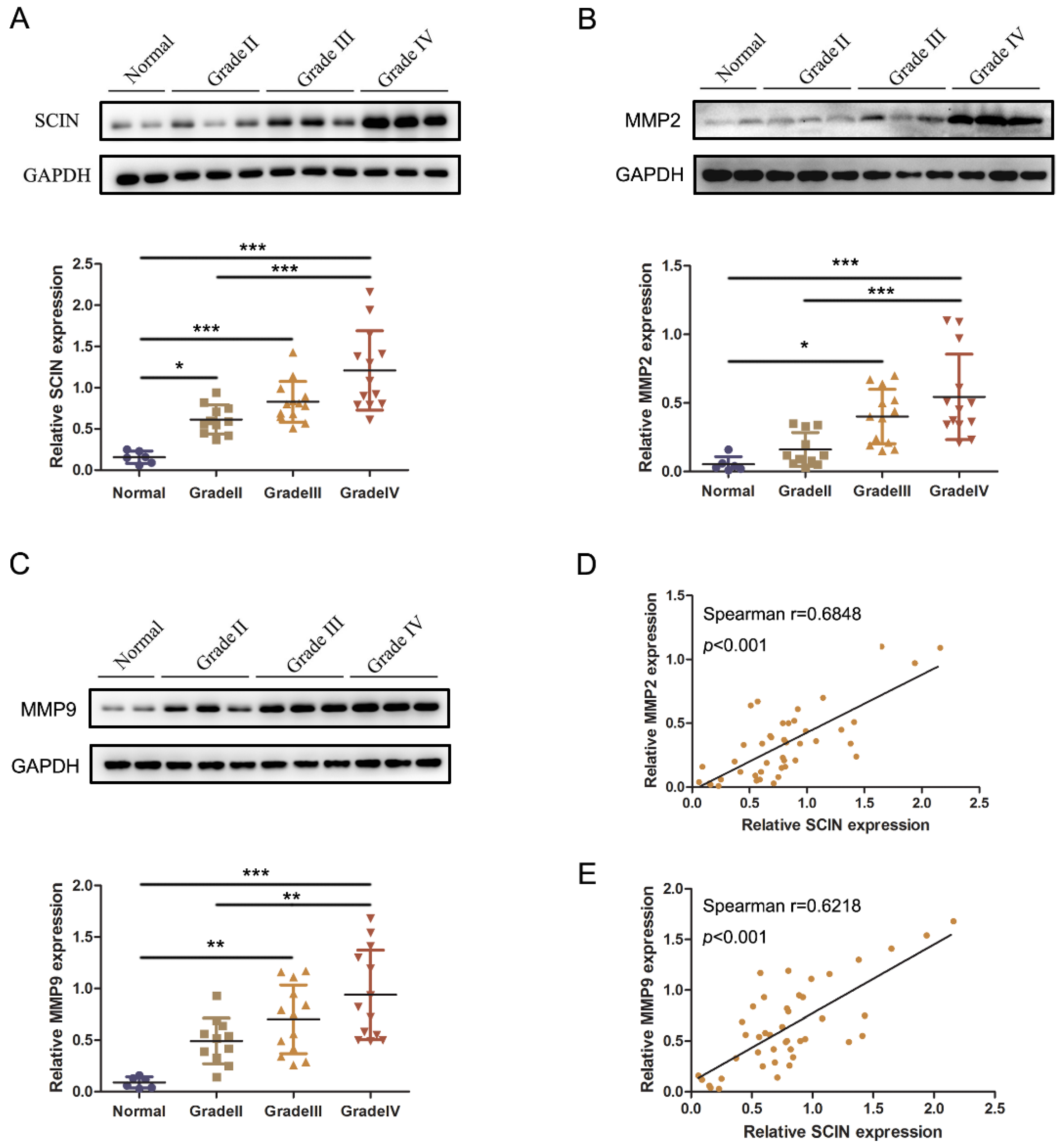
3.5. Co-Overexpressions of SCIN and MMP2/9 Are Associated with A Poor Prognosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K.; Burger, P.C.; Jouvet, A.; Scheithauer, B.W.; Kleihues, P. The 2007 WHO classification of tumours of the central nervous system. Acta Neuropathol. 2007, 114, 97–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostrom, Q.T.; Gittleman, H.; de Blank, P.M.; Finlay, J.L.; Gurney, J.G.; McKean-Cowdin, R.; Stearns, D.S.; Wolff, J.E.; Liu, M.; Wolinsky, Y.; et al. American Brain Tumor Association Adolescent and Young Adult Primary Brain and Central Nervous System Tumors Diagnosed in the United States in 2008–2012. Neuro Oncol. 2016, 18, i1–i50. [Google Scholar] [CrossRef] [Green Version]
- Maekawa, S.; Sakai, H. Inhibition of actin regulatory activity of the 74-kDa protein from bovine adrenal medulla (adseverin) by some phospholipids. J. Biol. Chem. 1990, 265, 10940–10942. [Google Scholar] [CrossRef]
- Del Castillo, A.R.; Lemaire, S.; Tchakarov, L.; Jeyapragasan, M.; Doucet, J.; Vitale, M.; Trifaró, J. Chromaffin cell scinderin, a novel calcium-dependent actin filament-severing protein. EMBO J. 1990, 9, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Shi, D.; Liu, T.; Yu, Z.; Zhou, C. Lentivirus-mediated silencing of SCIN inhibits proliferation of human lung carcinoma cells. Gene 2015, 554, 32–39. [Google Scholar] [CrossRef]
- Wang, D.; Sun, S.Q.; Yu, Y.H.; Wu, W.Z.; Yang, S.L.; Tan, J.M. Suppression of SCIN inhibits human prostate cancer cell proliferation and induces G0/G1 phase arrest. Int. J. Oncol. 2014, 44, 161–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.J.; Liu, J.Y.; Chen, J.; Wu, Y.X.; Yan, P.; Ji, C.D.; Wang, Y.X.; Xiang, D.F.; Zhang, X.; Zhang, P.; et al. Scinderin promotes the invasion and metastasis of gastric cancer cells and predicts the outcome of patients. Cancer Lett. 2016, 376, 110–117. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.; Chen, T.W.; Jiang, Y.B.; Wei, X.B.; Lu, C.D.; Li, J.J.; Xie, D.; Cheng, S.Q. Scinderin suppresses cell proliferation and predicts the poor prognosis of hepatocellular carcinoma. Oncol. Lett. 2020, 19, 2011–2020. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, D.R.; Kalyana-Sundaram, S.; Mahavisno, V.; Varambally, R.; Yu, J.; Briggs, B.B.; Barrette, T.R.; Anstet, M.J.; Kincead-Beal, C.; Kulkarni, P.; et al. Oncomine 3.0: Genes, pathways, and networks in a collection of 18,000 cancer gene expression profiles. Neoplasia 2007, 9, 166–180. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, D.R.; Yu, J.; Shanker, K.; Deshpande, N.; Varambally, R.; Ghosh, D.; Barrette, T.; Pandey, A.; Chinnaiyan, A.M. ONCOMINE: A cancer microarray database and integrated data-mining platform. Neoplasia 2004, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Zhang, K.N.; Wang, Q.; Li, G.; Zeng, F.; Zhang, Y.; Wu, F.; Chai, R.; Wang, Z.; Zhang, C.; et al. Chinese Glioma Genome Atlas (CGGA): A Comprehensive Resource with Functional Genomic Data from Chinese Glioma Patients. Genom. Proteom. Bioinform. 2021, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Martin-Broto, J.; Stacchiotti, S.; Lopez-Pousa, A.; Redondo, A.; Bernabeu, D.; de Alava, E.; Casali, P.G.; Italiano, A.; Gutierrez, A.; Moura, D.S.; et al. Pazopanib for treatment of advanced malignant and dedifferentiated solitary fibrous tumour: A multicentre, single-arm, phase 2 trial. Lancet. Oncol. 2019, 20, 134–144. [Google Scholar] [CrossRef]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef] [Green Version]
- Tomczak, K.; Czerwinska, P.; Wiznerowicz, M. The Cancer Genome Atlas (TCGA): An immeasurable source of knowledge. Contemp. Oncol. 2015, 19, A68–A77. [Google Scholar] [CrossRef] [PubMed]
- Colaprico, A.; Silva, T.C.; Olsen, C.; Garofano, L.; Cava, C.; Garolini, D.; Sabedot, T.S.; Malta, T.M.; Pagnotta, S.M.; Castiglioni, I.; et al. TCGAbiolinks: An R/Bioconductor package for integrative analysis of TCGA data. Nucleic Acids Res. 2016, 44, e71. [Google Scholar] [CrossRef]
- Lin, Q.; Li, J.; Zhu, D.; Niu, Z.; Pan, X.; Xu, P.; Ji, M.; Wei, Y.; Xu, J. Aberrant Scinderin Expression Correlates With Liver Metastasis and Poor Prognosis in Colorectal. Cancer Front. Pharmacol. 2019, 10, 1183. [Google Scholar]
- Chen, X.M.; Guo, J.M.; Chen, P.; Mao, L.G.; Feng, W.Y.; Le, D.H.; Li, K.Q. Suppression of scinderin modulates epithelialmesenchymal transition markers in highly metastatic gastric cancer cell line SGC7901. Mol. Med. Rep. 2014, 10, 2327–2333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, X.; Su, W.; Zhao, H.; Yang, S.; Zeng, T.; Wu, W.; Wang, D. Loss of scinderin decreased expression of epidermal growth factor receptor and promoted apoptosis of castration-resistant prostate cancer cells. FEBS Open Bio. 2018, 8, 743–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jian, W.; Zhang, X.; Wang, J.; Liu, Y.; Hu, C.; Wang, X.; Liu, R. Scinderin-knockdown inhibits proliferation and promotes apoptosis in human breast carcinoma cells. Oncol. Lett. 2018, 16, 3207–3214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.H.; Zhang, W.; Zhou, J.D.; Zhang, T.J.; Ma, J.C.; Xu, Z.J.; Lian, X.Y.; Wu, D.H.; Wen, X.M.; Deng, Z.Q.; et al. Decreased SCIN expression, associated with promoter methylation, is a valuable predictor for prognosis in acute myeloid leukemia. Mol. Carcinog. 2018, 57, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Du, X.; Chen, X.; Chen, C.; Wang, H.; Yang, Y.; Teng, L. MiR-301a-5p/SCIN promotes gastric cancer progression via regulating STAT3 and NF-kappaB signaling. J. Cancer 2021, 12, 5394–5403. [Google Scholar] [CrossRef]
- Schalper, K.A.; Rodriguez-Ruiz, M.E.; Diez-Valle, R.; Lopez-Janeiro, A.; Porciuncula, A.; Idoate, M.A.; Inoges, S.; de Andrea, C.; Lopez-Diaz de Cerio, A.; Tejada, S.; et al. Neoadjuvant nivolumab modifies the tumor immune microenvironment in resectable glioblastoma. Nat. Med. 2019, 25, 470–476. [Google Scholar] [CrossRef]
- Cloughesy, T.F.; Mochizuki, A.Y.; Orpilla, J.R.; Hugo, W.; Lee, A.H.; Davidson, T.B.; Wang, A.C.; Ellingson, B.M.; Rytlewski, J.A.; Sanders, C.M.; et al. Neoadjuvant anti-PD-1 immunotherapy promotes a survival benefit with intratumoral and systemic immune responses in recurrent glioblastoma. Nat. Med. 2019, 25, 477–486. [Google Scholar] [CrossRef]
- Pombo Antunes, A.R.; Scheyltjens, I.; Duerinck, J.; Neyns, B.; Movahedi, K.; Van Ginderachter, J.A. Understanding the glioblastoma immune microenvironment as basis for the development of new immunotherapeutic strategies. Elife 2020, 9, e52176. [Google Scholar] [CrossRef]
- Aldape, K.; Brindle, K.M.; Chesler, L.; Chopra, R.; Gajjar, A.; Gilbert, M.R.; Gottardo, N.; Gutmann, D.H.; Hargrave, D.; Holland, E.C.; et al. Challenges to curing primary brain tumours. Nat. Rev. Clin. Oncol. 2019, 16, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Li, G.H.; Arora, P.D.; Chen, Y.; McCulloch, C.A.; Liu, P. Multifunctional roles of gelsolin in health and diseases. Med. Res. Rev. 2012, 32, 999–1025. [Google Scholar] [CrossRef]
- Miyauchi, E.; Furuta, T.; Ohtsuki, S.; Tachikawa, M.; Uchida, Y.; Sabit, H.; Obuchi, W.; Baba, T.; Watanabe, M.; Terasaki, T.; et al. Identification of blood biomarkers in glioblastoma by SWATH mass spectrometry and quantitative targeted absolute proteomics. PLoS ONE 2018, 13, e0193799. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Furuta, T.; Sabit, H.; Tamai, S.; Jiapaer, S.; Dong, Y.; Kinoshita, M.; Uchida, Y.; Ohtsuki, S.; Terasaki, T.; et al. Gelsolin inhibits malignant phenotype of glioblastoma and is regulated by miR-654-5p and miR-450b-5p. Cancer Sci. 2020, 111, 2413–2422. [Google Scholar] [CrossRef] [PubMed]
- Jin, N.; Lang, M.J.; Weisman, L.S. Phosphatidylinositol 3,5-bisphosphate: Regulation of cellular events in space and time. Biochem. Soc. Trans. 2016, 44, 177–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, J.; Strunk, B.S.; Weisman, L.S. PI5P and PI(3,5)P2: Minor, but Essential Phosphoinositides. Cell Struct. Funct. 2017, 42, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Watakabe, A.; Kobayashi, R.; Helfman, D.M. N-tropomodulin: A novel isoform of tropomodulin identified as the major binding protein to brain tropomyosin. J. Cell Sci. 1996, 109, 2299–2310. [Google Scholar] [CrossRef] [PubMed]
- Weber, A.; Pennise, C.R.; Fowler, V.M. Tropomodulin increases the critical concentration of barbed end-capped actin filaments by converting ADP.P(i)-actin to ADP-actin at all pointed filament ends. J. Biol. Chem. 1999, 274, 34637–34645. [Google Scholar] [CrossRef] [Green Version]
- Binder, Z.A.; Thorne, A.H.; Bakas, S.; Wileyto, E.P.; Bilello, M.; Akbari, H.; Rathore, S.; Ha, S.M.; Zhang, L.; Ferguson, C.J.; et al. Epidermal Growth Factor Receptor Extracellular Domain Mutations in Glioblastoma Present Opportunities for Clinical Imaging and Therapeutic Development. Cancer Cell 2018, 34, 163–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Zhang, H.; Yu, L. HMGA2 promotes glioma invasion and poor prognosis via a long-range chromatin interaction. Cancer Med. 2018, 7, 3226–3239. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Yu, X.; Sun, S.; Zhang, X.; Yang, W.; Zhang, J.; Zhang, X.; Jiang, Z. Increased expression of MMP-2 and MMP-9 indicates poor prognosis in glioma recurrence. Biomed. Pharmacother. 2019, 118, 109369. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Features | Cases | High SCIN Expression | Low SCIN Expression | p Value | |
|---|---|---|---|---|---|
| Age (Mean ± SD, years) | 106 | 52.86 ± 11.488 | 56.81 ± 9.518 | 0.380 | |
| Gender | Male | 49 | 24 | 25 | 0.199 |
| Female | 57 | 35 | 22 | ||
| Tumor size (cm) < 3 | 35 | 16 | 19 | 0.148 | |
| ≥3 | 71 | 43 | 28 | ||
| KPS | <80 | 62 | 38 | 24 | 0.166 |
| ≥80 | 44 | 21 | 23 | ||
| WHO Grade | I | 3 | 1 | 2 | 0.006 * |
| II | 31 | 10 | 21 | ||
| III | 26 | 15 | 11 | ||
| IV | 46 | 33 | 13 | ||
| Survival state | Alive | 40 | 13 | 27 | <0.001 * |
| Death | 66 | 46 | 20 | ||
| MMP2 expression | High | 62 | 42 | 20 | 0.003 * |
| Low | 44 | 17 | 27 | ||
| MMP9 expression | High | 61 | 40 | 21 | 0.017 * |
| Low | 45 | 19 | 26 | ||
| Univariate Analysis | Multivariate Analysis | |||||||
|---|---|---|---|---|---|---|---|---|
| p Value | Hazard Ratio | 95% Confidence Interval | Concordance Index | p Value | Hazard Ratio | 95% Confidence Interval | Concordance Index | |
| SCIN | <0.001 | 2.727 | 1.608–4.625 | 0.622 | 0.034 | 1.857 | 1.047–3.292 | |
| MMP2 | 0.004 | 2.156 | 1.273–3.650 | 0.595 | 0.695 | 0.887 | 0.488–1.613 | |
| MMP9 | 0.001 | 2.467 | 1.442–4.220 | 0.588 | 0.039 | 1.851 | 1.032–3.317 | 0.723 |
| Tumor size | 0.024 | 1.914 | 1.088–3.366 | 0.567 | 0.297 | 1.368 | 0.760–2.463 | |
| WHO Grade | <0.001 | 6.048 | 3.074–11.902 | 0.672 | <0.001 | 5.300 | 2.499–11.240 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, Z.; Bai, L.; Zhang, D.; Zhang, T.; Ren, F. Scinderin Is a Novel Oncogene for Its Correlates with Poor Prognosis, Immune Infiltrates and Matrix Metalloproteinase-2/9 (MMP2/9) in Glioma. Brain Sci. 2022, 12, 1415. https://doi.org/10.3390/brainsci12101415
Wang Y, Li Z, Bai L, Zhang D, Zhang T, Ren F. Scinderin Is a Novel Oncogene for Its Correlates with Poor Prognosis, Immune Infiltrates and Matrix Metalloproteinase-2/9 (MMP2/9) in Glioma. Brain Sciences. 2022; 12(10):1415. https://doi.org/10.3390/brainsci12101415
Chicago/Turabian StyleWang, Yiwei, Zhongyan Li, Lili Bai, Dongyong Zhang, Tianchi Zhang, and Fu Ren. 2022. "Scinderin Is a Novel Oncogene for Its Correlates with Poor Prognosis, Immune Infiltrates and Matrix Metalloproteinase-2/9 (MMP2/9) in Glioma" Brain Sciences 12, no. 10: 1415. https://doi.org/10.3390/brainsci12101415
APA StyleWang, Y., Li, Z., Bai, L., Zhang, D., Zhang, T., & Ren, F. (2022). Scinderin Is a Novel Oncogene for Its Correlates with Poor Prognosis, Immune Infiltrates and Matrix Metalloproteinase-2/9 (MMP2/9) in Glioma. Brain Sciences, 12(10), 1415. https://doi.org/10.3390/brainsci12101415