


The Role of Astrocytes in the Mechanism of Perioperative Neurocognitive Disorders


Abstract
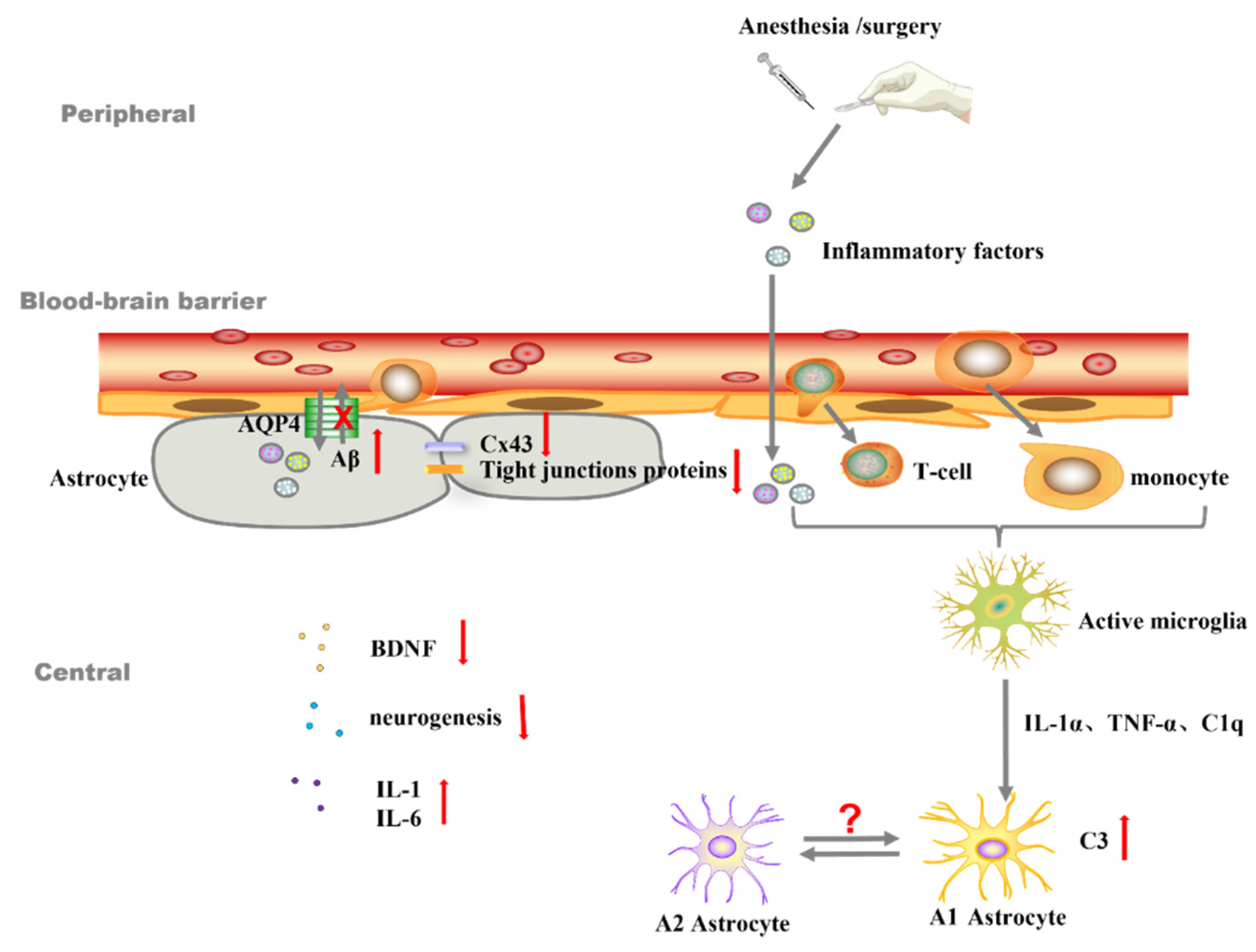
:1. Introduction
2. Different Phenotypes of Reactive Astrocytes and PND
3. Astrocytes Are Involved in the Development of PND Partially by Mediating Neuroinflammation
4. Abnormal Astrocytes Induce PND by Regulating Synapse Formation, Quantity, and Transmission of Neurotransmitters
5. Astrocytes Maintain Homeostasis of Intracerebral Microenvironment
6. Astrocytes Assist Neurons in Trophic Metabolism
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Evered, L.; Silbert, B.; Knopman, D.S.; Scott, D.A.; DeKosky, S.T.; Rasmussen, L.S.; Oh, E.S.; Crosby, G.; Berger, M.; Eckenhoff, R.G.; et al. Recommendations for the Nomenclature of Cognitive Change Associated with Anaesthesia and Surgery-2018. Anesthesiology 2018, 129, 872–879. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Mai, L.; Zhu, X.; Huang, F.; He, H. The Role of Microglia in Perioperative Neurocognitive Disorders. Front. Cell. Neurosci. 2020, 14, 261. [Google Scholar] [CrossRef] [PubMed]
- Hovens, I.B.; Schoemaker, R.G.; van der Zee, E.A.; Heineman, E.; Izaks, G.J.; van Leeuwen, B.L. Thinking through postoperative cognitive dysfunction: How to bridge the gap between clinical and pre-clinical perspectives. Brain Behav. Immun. 2012, 26, 1169–1179. [Google Scholar] [CrossRef] [Green Version]
- Needham, M.J.; Webb, C.E.; Bryden, D.C. Postoperative cognitive dysfunction and dementia: What we need to know and do. Br. J. Anaesth. 2017, 119, i115–i125. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.; Monk, T.; Rasmussen, L.S.; Abildstrom, H.; Houx, P.; Korttila, K.; Kuipers, H.M.; Hanning, C.D.; Siersma, V.D.; Kristensen, D.; et al. Postoperative cognitive dysfunction in middle-aged patients. Anesthesiology 2002, 96, 1351–1357. [Google Scholar] [CrossRef]
- Meybohm, P.; Renner, J.; Broch, O.; Caliebe, D.; Albrecht, M.; Cremer, J.; Haake, N.; Scholz, J.; Zacharowski, K.; Bein, B. Postoperative neurocognitive dysfunction in patients undergoing cardiac surgery after remote ischemic preconditioning: A double-blind randomized controlled pilot study. PLoS ONE 2013, 8, e64743. [Google Scholar] [CrossRef] [Green Version]
- Monk, T.G.; Weldon, B.C.; Garvan, C.W.; Dede, D.E.; van der Aa, M.T.; Heilman, K.M.; Gravenstein, J.S. Predictors of cognitive dysfunction after major noncardiac surgery. Anesthesiology 2008, 108, 18–30. [Google Scholar] [CrossRef] [Green Version]
- Subramaniyan, S.; Terrando, N. Neuroinflammation and Perioperative Neurocognitive Disorders. Anesth. Analg. 2019, 128, 781–788. [Google Scholar] [CrossRef]
- Liu, Y.; Yin, Y. Emerging Roles of Immune Cells in Postoperative Cognitive Dysfunction. Mediat. Inflamm. 2018, 2018, 6215350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H.; Liu, W.; Fang, H. Inflammation caused by peripheral immune cells across into injured mouse blood brain barrier can worsen postoperative cognitive dysfunction induced by isoflurane. BMC Cell Biol. 2018, 19, 23. [Google Scholar] [CrossRef]
- Tan, X.X.; Qiu, L.L.; Sun, J. Research Progress on the Role of Inflammatory Mechanisms in the Development of Postoperative Cognitive Dysfunction. Biomed. Res. Int. 2021, 2021, 3883204. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhang, M.; Zhang, Z.; Dong, W.; Wang, Q.; Ren, J. Ratio of beta-amyloid protein (Abeta) and Tau predicts the postoperative cognitive dysfunction on patients undergoing total hip/knee replacement surgery. Exp. Ther. Med. 2018, 15, 878–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, W.; Lu, K.; Wang, J.; Wu, A.; Yue, Y. Activation of mTOR signaling leads to orthopedic surgery-induced cognitive decline in mice through beta-amyloid accumulation and tau phosphorylation. Mol. Med. Rep. 2016, 14, 3925–3934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, H.; Liu, B.; Chen, Y.; Zhang, J. Learning, memory and synaptic plasticity in hippocampus in rats exposed to sevoflurane. Int. J. Dev. Neurosci. 2016, 48, 38–49. [Google Scholar] [CrossRef]
- Lin, X.; Chen, Y.; Zhang, P.; Chen, G.; Zhou, Y.; Yu, X. The potential mechanism of postoperative cognitive dysfunction in older people. Exp. Gerontol. 2020, 130, 110791. [Google Scholar] [CrossRef]
- Arranz, A.M.; De Strooper, B. The role of astroglia in Alzheimer’s disease: Pathophysiology and clinical implications. Lancet Neurol. 2019, 18, 406–414. [Google Scholar] [CrossRef]
- Shu, X.; Sun, Y.; Sun, X.; Zhou, Y.; Bian, Y.; Shu, Z.; Ding, J.; Lu, M.; Hu, G. The effect of fluoxetine on astrocyte autophagy flux and injured mitochondria clearance in a mouse model of depression. Cell Death Dis. 2019, 10, 577. [Google Scholar] [CrossRef] [Green Version]
- Dietz, A.G.; Goldman, S.A.; Nedergaard, M. Glial cells in schizophrenia: A unified hypothesis. Lancet Psychiatry 2020, 7, 272–281. [Google Scholar] [CrossRef]
- Cai, Z.; Wan, C.Q.; Liu, Z. Astrocyte and Alzheimer’s disease. J. Neurol. 2017, 264, 2068–2074. [Google Scholar] [CrossRef]
- Booth, H.D.E.; Hirst, W.D.; Wade-Martins, R. The Role of Astrocyte Dysfunction in Parkinson’s Disease Pathogenesis. Trends Neurosci. 2017, 40, 358–370. [Google Scholar] [CrossRef]
- Diaz-Castro, B.; Gangwani, M.R.; Yu, X.; Coppola, G.; Khakh, B.S. Astrocyte molecular signatures in Huntington’s disease. Sci. Transl. Med. 2019, 11, eaaw8546. [Google Scholar] [CrossRef] [Green Version]
- Liddelow, S.A.; Barres, B.A. Reactive Astrocytes: Production, Function, and Therapeutic Potential. Immunity 2017, 46, 957–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhauser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; et al. Reactive astrocyte nomenclature, definitions, and future directions. Nat. Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Munch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamanian, J.L.; Xu, L.; Foo, L.C.; Nouri, N.; Zhou, L.; Giffard, R.G.; Barres, B.A. Genomic analysis of reactive astrogliosis. J. Neurosci. 2012, 32, 6391–6410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, C.; Liu, J.; Lin, D.; Zhang, J.; Terrando, N.; Wu, A. Complement activation contributes to perioperative neurocognitive disorders in mice. J. Neuroinflamm. 2018, 15, 254. [Google Scholar] [CrossRef]
- Li, D.; Chen, M.; Meng, T.; Fei, J. Hippocampal microglial activation triggers a neurotoxic-specific astrocyte response and mediates etomidate-induced long-term synaptic inhibition. J. Neuroinflamm. 2020, 17, 109. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Yang, N.; Li, Y.; Li, Y.; Hong, J.; Wang, Q.; Liu, K.; Han, D.; Han, Y.; Mi, X.; et al. Cholecystokinin octapeptide improves hippocampal glutamatergic synaptogenesis and postoperative cognition by inhibiting induction of A1 reactive astrocytes in aged mice. CNS Neurosci. Ther. 2021, 27, 1374–1384. [Google Scholar] [CrossRef]
- Li, Y.; Wu, Z.Y.; Zheng, W.C.; Wang, J.X.; Yue, X.; Song, R.X.; Gao, J.G. Esketamine alleviates postoperative cognitive decline via stimulator of interferon genes/ TANK-binding kinase 1 signaling pathway in aged rats. Brain Res. Bull. 2022, 187, 169–180. [Google Scholar] [CrossRef]
- Fan, Y.Y.; Huo, J. A1/A2 astrocytes in central nervous system injuries and diseases: Angels or devils? Neurochem. Int. 2021, 148, 105080. [Google Scholar] [CrossRef]
- Mulkey, D.K.; Olsen, M.L.; Ou, M.; Cleary, C.M.; Du, G. Putative Roles of Astrocytes in General Anesthesia. Curr. Neuropharmacol. 2022, 20, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Chen, L.; Liao, P.; Huang, L.; Chen, Z.; Liao, D.; Yang, L.; Wang, J.; Yu, G.; Wang, L.; et al. Astroglial dysfunctions drive aberrant synaptogenesis and social behavioral deficits in mice with neonatal exposure to lengthy general anesthesia. PLoS Biol. 2019, 17, e3000086. [Google Scholar] [CrossRef]
- Safavynia, S.A.; Goldstein, P.A. The Role of Neuroinflammation in Postoperative Cognitive Dysfunction: Moving From Hypothesis to Treatment. Front. Psychiatry 2018, 9, 752. [Google Scholar] [CrossRef]
- Kristek, G.; Rados, I.; Kristek, D.; Kapural, L.; Neskovic, N.; Skiljic, S.; Horvat, V.; Mandic, S.; Harsanji-Drenjancevic, I. Influence of postoperative analgesia on systemic inflammatory response and postoperative cognitive dysfunction after femoral fractures surgery: A randomized controlled trial. Reg. Anesth. Pain Med. 2019, 44, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Gao, X.; Wang, Z.; Jiang, X.; Lu, S.; Xu, J.; Qin, G.; Gu, Z.; Huang, D. Preoperative Chronic and Acute Pain Affects Postoperative Cognitive Function Mediated by Neurotransmitters. J. Mol. Neurosci. 2021, 71, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Koyama, T.; Kawano, T.; Iwata, H.; Aoyama, B.; Eguchi, S.; Nishigaki, A.; Yamanaka, D.; Tateiwa, H.; Shigematsu-Locatelli, M.; Locatelli, F.M.; et al. Acute postoperative pain exacerbates neuroinflammation and related delirium-like cognitive dysfunction in rats. J. Anesth. 2019, 33, 482–486. [Google Scholar] [CrossRef]
- Waisman, A.; Liblau, R.S.; Becher, B. Innate and adaptive immune responses in the CNS. Lancet Neurol. 2015, 14, 945–955. [Google Scholar] [CrossRef]
- Linnerbauer, M.; Wheeler, M.A.; Quintana, F.J. Astrocyte Crosstalk in CNS Inflammation. Neuron 2020, 108, 608–622. [Google Scholar] [CrossRef]
- Xu, J.; Dong, H.; Qian, Q.; Zhang, X.; Wang, Y.; Jin, W.; Qian, Y. Astrocyte-derived CCL2 participates in surgery-induced cognitive dysfunction and neuroinflammation via evoking microglia activation. Behav. Brain Res. 2017, 332, 145–153. [Google Scholar] [CrossRef]
- Li, N.; Zhang, X.; Dong, H.; Hu, Y.; Qian, Y. Bidirectional relationship of mast cells-neurovascular unit communication in neuroinflammation and its involvement in POCD. Behav. Brain Res. 2017, 322, 60–69. [Google Scholar] [CrossRef]
- Zhang, X.; Yao, H.; Qian, Q.; Li, N.; Jin, W.; Qian, Y. Cerebral Mast Cells Participate In Postoperative Cognitive Dysfunction by Promoting Astrocyte Activation. Cell Physiol. Biochem. 2016, 40, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Tian, A.; Ma, H.; Zhang, R.; Tan, W.; Wang, X.; Wu, B.; Wang, J.; Wan, C. Interleukin17A Promotes Postoperative Cognitive Dysfunction by Triggering beta-Amyloid Accumulation via the Transforming Growth Factor-beta (TGFbeta)/Smad Signaling Pathway. PLoS ONE 2015, 10, e0141596. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, X.; Yu, W.; Ying, J.; Fang, P.; Zheng, Q.; Feng, X.; Hu, J.; Xiao, F.; Chen, S.; et al. CB2R Activation Regulates TFEB-Mediated Autophagy and Affects Lipid Metabolism and Inflammation of Astrocytes in POCD. Front. Immunol. 2022, 13, 836494. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, Y.; Li, K.; Li, X.; Guo, M.; Peng, M. A Noradrenergic Lesion Attenuates Surgery-Induced Cognitive Impairment in Rats by Suppressing Neuroinflammation. Front. Mol. Neurosci. 2021, 14, 752838. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.R.; Cao, F.; Zhang, Y.; Peng, S. Electroacupuncture reduces astrocyte number and oxidative stress in aged rats with surgery-induced cognitive dysfunction. J. Int. Med. Res. 2019, 47, 3860–3873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Wu, X.; Ye, L.; Bai, Y.; Zhang, H.; Xuan, Z.; Feng, Y.; Zhang, P.; Chen, Y.; Yan, Y.; et al. Edaravone at high concentrations attenuates cognitive dysfunctions induced by abdominal surgery under general anesthesia in aged mice. Metab. Brain Dis. 2020, 35, 373–383. [Google Scholar] [CrossRef]
- Liu, X.; Ying, J.; Wang, X.; Zheng, Q.; Zhao, T.; Yoon, S.; Yu, W.; Yang, D.; Fang, Y.; Hua, F. Astrocytes in Neural Circuits: Key Factors in Synaptic Regulation and Potential Targets for Neurodevelopmental Disorders. Front. Mol. Neurosci. 2021, 14, 729273. [Google Scholar] [CrossRef]
- Chung, W.S.; Allen, N.J.; Eroglu, C. Astrocytes Control Synapse Formation, Function, and Elimination. Cold Spring Harb. Perspect. Biol. 2015, 7, a020370. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Li, L.L.; Sun, Z.T.; Zhang, W.; Han, X.P. Effects of Dexmedetomidine on Postoperative Cognitive Dysfunction and Serum Levels of b-Amyloid and Neuronal Microtubule-Associated Protein in Orthotopic Liver Transplantation Patients. Ann. Transpl. 2016, 21, 508–515. [Google Scholar] [CrossRef]
- Son, S.M.; Nam, D.W.; Cha, M.Y.; Kim, K.H.; Byun, J.; Ryu, H.; Mook-Jung, I. Thrombospondin-1 prevents amyloid beta-mediated synaptic pathology in Alzheimer’s disease. Neurobiol. Aging 2015, 36, 3214–3227. [Google Scholar] [CrossRef]
- Chung, W.S.; Clarke, L.E.; Wang, G.X.; Stafford, B.K.; Sher, A.; Chakraborty, C.; Joung, J.; Foo, L.C.; Thompson, A.; Chen, C.; et al. Astrocytes mediate synapse elimination through MEGF10 and MERTK pathways. Nature 2013, 504, 394–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Kim, J.Y.; Noh, S.; Lee, H.; Lee, S.Y.; Mun, J.Y.; Park, H.; Chung, W.S. Astrocytes phagocytose adult hippocampal synapses for circuit homeostasis. Nature 2021, 590, 612–617. [Google Scholar] [CrossRef] [PubMed]
- Shui, M.; Sun, Y.; Lin, D.; Xue, Z.; Liu, J.; Wu, A.; Wei, C. Anomalous Levels of CD47/Signal Regulatory Protein Alpha in the Hippocampus Lead to Excess Microglial Engulfment in Mouse Model of Perioperative Neurocognitive Disorders. Front. Neurosci. 2022, 16, 788675. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Arboledas, A.; Acharya, M.M.; Tenner, A.J. The Role of Complement in Synaptic Pruning and Neurodegeneration. Immunotargets Ther. 2021, 10, 373–386. [Google Scholar] [CrossRef]
- Eulenburg, V.; Gomeza, J. Neurotransmitter transporters expressed in glial cells as regulators of synapse function. Brain Res. Rev. 2010, 63, 103–112. [Google Scholar] [CrossRef]
- Qu, X.; Xu, C.; Wang, H.; Xu, J.; Liu, W.; Wang, Y.; Jia, X.; Xie, Z.; Xu, Z.; Ji, C.; et al. Hippocampal glutamate level and glutamate aspartate transporter (GLAST) are up-regulated in senior rat associated with isoflurane-induced spatial learning/memory impairment. Neurochem. Res. 2013, 38, 59–73. [Google Scholar] [CrossRef]
- Cooper, A.J. Possible treatment of end-stage hyperammonemic encephalopathy by inhibition of glutamine synthetase. Metab. Brain Dis. 2013, 28, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Skytt, D.M.; Klawonn, A.M.; Stridh, M.H.; Pajecka, K.; Patruss, Y.; Quintana-Cabrera, R.; Bolanos, J.P.; Schousboe, A.; Waagepetersen, H.S. siRNA knock down of glutamate dehydrogenase in astrocytes affects glutamate metabolism leading to extensive accumulation of the neuroactive amino acids glutamate and aspartate. Neurochem. Int. 2012, 61, 490–497. [Google Scholar] [CrossRef]
- Li, Y.; Wang, S.; Ran, K.; Hu, Z.; Liu, Z.; Duan, K. Differential hippocampal protein expression between normal aged rats and aged rats with postoperative cognitive dysfunction: A proteomic analysis. Mol. Med. Rep. 2015, 12, 2953–2960. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, S.; Gharagozloo, M.; Simard, C.; Gris, D. Astrocytes Maintain Glutamate Homeostasis in the CNS by Controlling the Balance between Glutamate Uptake and Release. Cells 2019, 8, 184. [Google Scholar] [CrossRef]
- Bezzi, P.; Domercq, M.; Brambilla, L.; Galli, R.; Schols, D.; De Clercq, E.; Vescovi, A.; Bagetta, G.; Kollias, G.; Meldolesi, J.; et al. CXCR4-activated astrocyte glutamate release via TNFalpha: Amplification by microglia triggers neurotoxicity. Nat. Neurosci. 2001, 4, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Morales, M.; Garcia-Colunga, J. Effects of nicotine on K+ currents and nicotinic receptors in astrocytes of the hippocampal CA1 region. Neuropharmacology 2009, 56, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Takata, N.; Mishima, T.; Hisatsune, C.; Nagai, T.; Ebisui, E.; Mikoshiba, K.; Hirase, H. Astrocyte calcium signaling transforms cholinergic modulation to cortical plasticity in vivo. J. Neurosci. 2011, 31, 18155–18165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Xu, G.; Zhang, X.; Ren, Y.; Yang, T.; Xiao, C.; Zhou, C. Malfunction of astrocyte and cholinergic input is involved in postoperative impairment of hippocampal synaptic plasticity and cognitive function. Neuropharmacology 2022, 217, 109191. [Google Scholar] [CrossRef] [PubMed]
- Coyle, J.T.; Balu, D.; Wolosker, H. D-Serine, the Shape-Shifting NMDA Receptor Co-agonist. Neurochem. Res. 2020, 45, 1344–1353. [Google Scholar] [CrossRef]
- Wolosker, H.; Balu, D.T.; Coyle, J.T. The Rise and Fall of the d-Serine-Mediated Gliotransmission Hypothesis. Trends Neurosci. 2016, 39, 712–721. [Google Scholar] [CrossRef] [Green Version]
- Kartvelishvily, E.; Shleper, M.; Balan, L.; Dumin, E.; Wolosker, H. Neuron-derived D-serine release provides a novel means to activate N-methyl-D-aspartate receptors. J. Biol. Chem. 2006, 281, 14151–14162. [Google Scholar] [CrossRef] [Green Version]
- Perez, E.J.; Tapanes, S.A.; Loris, Z.B.; Balu, D.T.; Sick, T.J.; Coyle, J.T.; Liebl, D.J. Enhanced astrocytic d-serine underlies synaptic damage after traumatic brain injury. J. Clin. Investig. 2017, 127, 3114–3125. [Google Scholar] [CrossRef]
- Cohen-Salmon, M.; Slaoui, L.; Mazare, N.; Gilbert, A.; Oudart, M.; Alvear-Perez, R.; Elorza-Vidal, X.; Chever, O.; Boulay, A.C. Astrocytes in the regulation of cerebrovascular functions. Glia 2021, 69, 817–841. [Google Scholar] [CrossRef]
- Mathiisen, T.M.; Lehre, K.P.; Danbolt, N.C.; Ottersen, O.P. The perivascular astroglial sheath provides a complete covering of the brain microvessels: An electron microscopic 3D reconstruction. Glia 2010, 58, 1094–1103. [Google Scholar] [CrossRef]
- Nagelhus, E.A.; Ottersen, O.P. Physiological roles of aquaporin-4 in brain. Physiol. Rev. 2013, 93, 1543–1562. [Google Scholar] [CrossRef] [Green Version]
- Velagapudi, R.; Subramaniyan, S.; Xiong, C.; Porkka, F.; Rodriguiz, R.M.; Wetsel, W.C.; Terrando, N. Orthopedic Surgery Triggers Attention Deficits in a Delirium-Like Mouse Model. Front. Immunol. 2019, 10, 2675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, X.; Liu, S.; Lian, C.; Li, H.; Li, K.; Li, L.; Zhao, G. Dysfunction of the Glymphatic System as a Potential Mechanism of Perioperative Neurocognitive Disorders. Front. Aging Neurosci. 2021, 13, 659457. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Ming, J.; Liu, S.; Lai, B.; Fang, F.; Cang, J. Sevoflurane enhanced the clearance of Abeta1-40 in hippocampus under surgery via up-regulating AQP-4 expression in astrocyte. Life Sci. 2019, 221, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Harrison, I.F.; Ismail, O.; Machhada, A.; Colgan, N.; Ohene, Y.; Nahavandi, P.; Ahmed, Z.; Fisher, A.; Meftah, S.; Murray, T.K.; et al. Impaired glymphatic function and clearance of tau in an Alzheimer’s disease model. Brain 2020, 143, 2576–2593. [Google Scholar] [CrossRef]
- Dong, R.; Han, Y.; Jiang, L.; Liu, S.; Zhang, F.; Peng, L.; Wang, Z.; Ma, Z.; Xia, T.; Gu, X. Connexin 43 gap junction-mediated astrocytic network reconstruction attenuates isoflurane-induced cognitive dysfunction in mice. J. Neuroinflamm. 2022, 19, 64. [Google Scholar] [CrossRef]
- Pivoriunas, A.; Verkhratsky, A. Astrocyte-Endotheliocyte Axis in the Regulation of the Blood-Brain Barrier. Neurochem. Res. 2021, 46, 2538–2550. [Google Scholar] [CrossRef]
- Wang, B.; Li, S.; Cao, X.; Dou, X.; Li, J.; Wang, L.; Wang, M.; Bi, Y. Blood-brain Barrier Disruption Leads to Postoperative Cognitive Dysfunction. Curr. Neurovasc. Res. 2017, 14, 359–367. [Google Scholar] [CrossRef]
- Hu, N.; Guo, D.; Wang, H.; Xie, K.; Wang, C.; Li, Y.; Wang, C.; Wang, C.; Yu, Y.; Wang, G. Involvement of the blood-brain barrier opening in cognitive decline in aged rats following orthopedic surgery and high concentration of sevoflurane inhalation. Brain Res. 2014, 1551, 13–24. [Google Scholar] [CrossRef]
- Horng, S.; Therattil, A.; Moyon, S.; Gordon, A.; Kim, K.; Argaw, A.T.; Hara, Y.; Mariani, J.N.; Sawai, S.; Flodby, P.; et al. Astrocytic tight junctions control inflammatory CNS lesion pathogenesis. J. Clin. Investig. 2017, 127, 3136–3151. [Google Scholar] [CrossRef]
- Sweeney, M.D.; Zhao, Z.; Montagne, A.; Nelson, A.R.; Zlokovic, B.V. Blood-Brain Barrier: From Physiology to Disease and Back. Physiol. Rev. 2019, 99, 21–78. [Google Scholar] [CrossRef] [PubMed]
- Spampinato, S.F.; Bortolotto, V.; Canonico, P.L.; Sortino, M.A.; Grilli, M. Astrocyte-Derived Paracrine Signals: Relevance for Neurogenic Niche Regulation and Blood-Brain Barrier Integrity. Front. Pharmacol. 2019, 10, 1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, J.I.; Dodelet-Devillers, A.; Kebir, H.; Ifergan, I.; Fabre, P.J.; Terouz, S.; Sabbagh, M.; Wosik, K.; Bourbonniere, L.; Bernard, M.; et al. The Hedgehog pathway promotes blood-brain barrier integrity and CNS immune quiescence. Science 2011, 334, 1727–1731. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Li, Z.; Ma, L.; Yang, N.; Guo, X. Isoflurane-Induced Postoperative Neurovascular and Cognitive Dysfunction Is Associated with VEGF Overexpression in Aged Rats. J. Mol. Neurosci. 2019, 69, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.F.; Wang, P.; Jiang, Y.X.; Zhang, X.P.; Shi, W.; Luo, Y.W. Effects of Apolipoprotein Epsilon epsilon4 allele on early postoperative cognitive dysfunction after anesthesia. Anaesthesist 2021, 70, 60–67. [Google Scholar] [CrossRef]
- Jackson, R.J.; Meltzer, J.C.; Nguyen, H.; Commins, C.; Bennett, R.E.; Hudry, E.; Hyman, B.T. APOE4 derived from astrocytes leads to blood-brain barrier impairment. Brain 2021, 145, 3582–3593. [Google Scholar] [CrossRef]
- Methia, N.; Andre, P.; Hafezi-Moghadam, A.; Economopoulos, M.; Thomas, K.L.; Wagner, D.D. ApoE deficiency compromises the blood brain barrier especially after injury. Mol. Med. 2001, 7, 810–815. [Google Scholar] [CrossRef]
- Belanger, M.; Allaman, I.; Magistretti, P.J. Brain energy metabolism: Focus on astrocyte-neuron metabolic cooperation. Cell Metab. 2011, 14, 724–738. [Google Scholar] [CrossRef] [Green Version]
- Morita, M.; Ikeshima-Kataoka, H.; Kreft, M.; Vardjan, N.; Zorec, R.; Noda, M. Metabolic Plasticity of Astrocytes and Aging of the Brain. Int. J. Mol. Sci. 2019, 20, 941. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, S. Metabolic Contribution and Cerebral Blood Flow Regulation by Astrocytes in the Neurovascular Unit. Cells 2022, 11, 813. [Google Scholar] [CrossRef]
- Acosta, C.; Anderson, H.D.; Anderson, C.M. Astrocyte dysfunction in Alzheimer disease. J. Neurosci. Res. 2017, 95, 2430–2447. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Iwamoto, T.; Kimura, S.; Nakao, S. Persistent isoflurane-induced hypotension causes hippocampal neuronal damage in a rat model of chronic cerebral hypoperfusion. J. Anesth. 2018, 32, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Martensson, J.; Gogenur, I.; Asghar, M.S. Exploring Postoperative Cognitive Dysfunction and Delirium in Noncardiac Surgery Using MRI: A Systematic Review. Neural Plast. 2018, 2018, 1281657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, S.A.; DeMuth, T.M.; Miller, K.N.; Pugh, T.D.; Polewski, M.A.; Colman, R.J.; Eliceiri, K.W.; Beasley, T.M.; Johnson, S.C.; Anderson, R.M. Regional metabolic heterogeneity of the hippocampus is nonuniformly impacted by age and caloric restriction. Aging Cell 2016, 15, 100–110. [Google Scholar] [CrossRef] [Green Version]
- Femenia, T.; Gimenez-Cassina, A.; Codeluppi, S.; Fernandez-Zafra, T.; Katsu-Jimenez, Y.; Terrando, N.; Eriksson, L.I.; Gomez-Galan, M. Disrupted Neuroglial Metabolic Coupling after Peripheral Surgery. J. Neurosci. 2018, 38, 452–464. [Google Scholar] [CrossRef] [Green Version]
- Hadjihambi, A.; Karagiannis, A.; Theparambil, S.M.; Ackland, G.L.; Gourine, A.V. The effect of general anaesthetics on brain lactate release. Eur. J. Pharmacol. 2020, 881, 173188. [Google Scholar] [CrossRef]
- Suzuki, A.; Stern, S.A.; Bozdagi, O.; Huntley, G.W.; Walker, R.H.; Magistretti, P.J.; Alberini, C.M. Astrocyte-neuron lactate transport is required for long-term memory formation. Cell 2011, 144, 810–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, L.A.; Korol, D.L.; Gold, P.E. Lactate produced by glycogenolysis in astrocytes regulates memory processing. PLoS ONE 2011, 6, e28427. [Google Scholar] [CrossRef]
- Zhang, J.; Tan, H.; Jiang, W.; Zuo, Z. Amantadine alleviates postoperative cognitive dysfunction possibly by increasing glial cell line-derived neurotrophic factor in rats. Anesthesiology 2014, 121, 773–785. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.; Zhu, T.; Chen, C.; Zhang, G.; Zhang, J.; Wang, L.; Zhang, L.; Wang, M.; Wang, X. Serum glial cell line-derived neurotrophic factor levels and postoperative cognitive dysfunction after surgery for rheumatic heart disease. J. Thorac. Cardiovasc. Surg. 2018, 155, 958–965.e1. [Google Scholar] [CrossRef]
- Pertusa, M.; Garcia-Matas, S.; Mammeri, H.; Adell, A.; Rodrigo, T.; Mallet, J.; Cristofol, R.; Sarkis, C.; Sanfeliu, C. Expression of GDNF transgene in astrocytes improves cognitive deficits in aged rats. Neurobiol. Aging 2008, 29, 1366–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gui, L.; Lei, X.; Zuo, Z. Decrease of glial cell-derived neurotrophic factor contributes to anesthesia- and surgery-induced learning and memory dysfunction in neonatal rats. J. Mol. Med. 2017, 95, 369–379. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| The Implication of Astrocytes in PND | References | |
|---|---|---|
| phenotypes of reactive astrocytes |
| [27,28,29] |
| [31,32] | |
| Mediating neuroinflammation |
| [32] |
| [37,38] | |
| [39] | |
| [40] | |
| [41] | |
| [42] | |
| [43] | |
| Regulating synapse formation, quantity, and transmission of neurotransmitters |
| [46,47] |
| [26,49,50] | |
| [52,53,54,55,56] | |
| [60,61] | |
| [64,65] | |
| Maintain homeostasis of intracerebral microenvironment |
| [69,70] |
| [73] | |
| [75,76,77] | |
| [78,79,80,81,82,83] | |
| Assist neurons in trophic metabolism |
| [88,89,90] |
| [91,92,93] | |
| [97,98] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Y.; Lin, X.; Liu, X.; Yu, K.; Miao, H.; Li, T. The Role of Astrocytes in the Mechanism of Perioperative Neurocognitive Disorders. Brain Sci. 2022, 12, 1435. https://doi.org/10.3390/brainsci12111435
Cao Y, Lin X, Liu X, Yu K, Miao H, Li T. The Role of Astrocytes in the Mechanism of Perioperative Neurocognitive Disorders. Brain Sciences. 2022; 12(11):1435. https://doi.org/10.3390/brainsci12111435
Chicago/Turabian StyleCao, Ying, Xiaowan Lin, Xiao Liu, Kang Yu, Huihui Miao, and Tianzuo Li. 2022. "The Role of Astrocytes in the Mechanism of Perioperative Neurocognitive Disorders" Brain Sciences 12, no. 11: 1435. https://doi.org/10.3390/brainsci12111435
APA StyleCao, Y., Lin, X., Liu, X., Yu, K., Miao, H., & Li, T. (2022). The Role of Astrocytes in the Mechanism of Perioperative Neurocognitive Disorders. Brain Sciences, 12(11), 1435. https://doi.org/10.3390/brainsci12111435