
Functional Connectivity States of Alpha Rhythm Sources in the Human Cortex at Rest: Implications for Real-Time Brain State Dependent EEG-TMS



Abstract
:1. Introduction
2. Methods
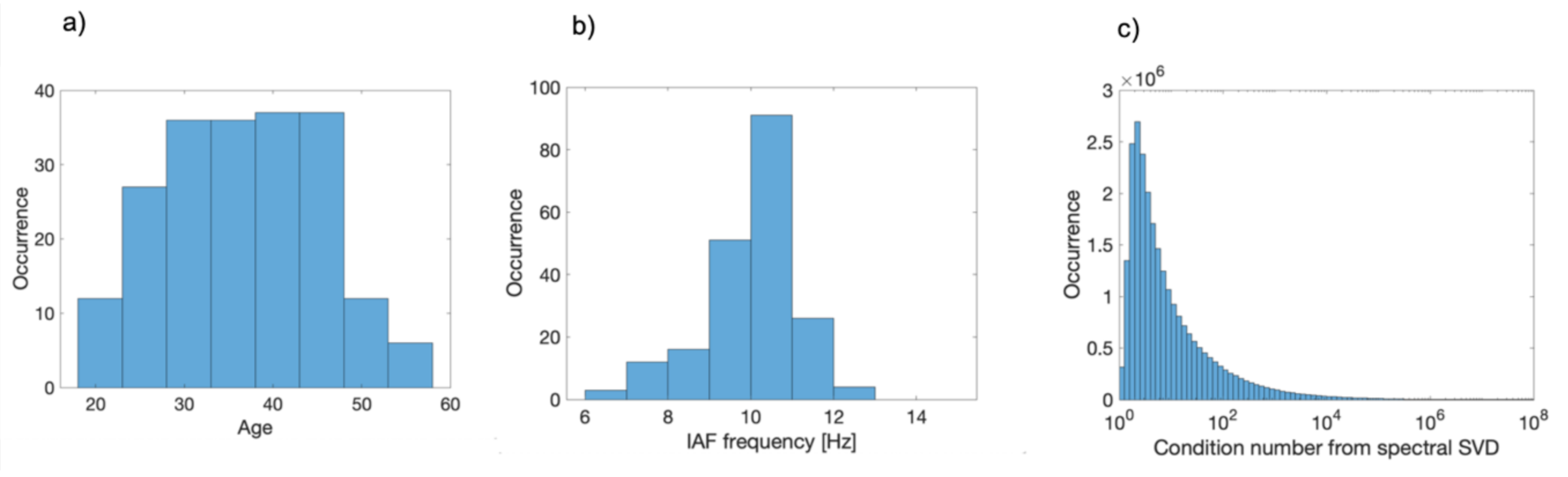
2.1. Dataset and Acquisition
2.2. MEG Data Preprocessing
2.3. Anatomical Data Processing
2.4. MEG Source Reconstruction Based on Individual Anatomies
2.5. Spectral Analysis
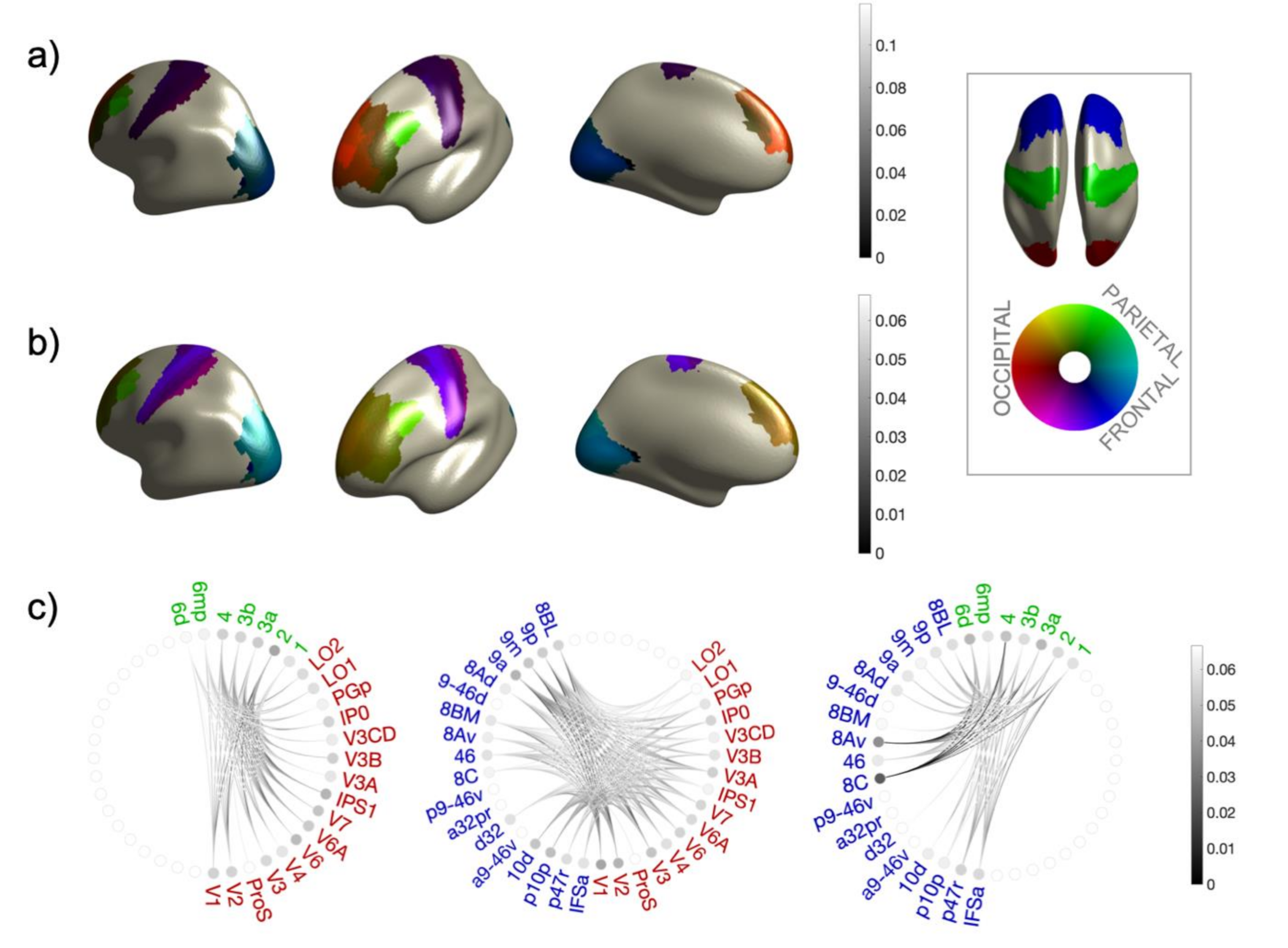
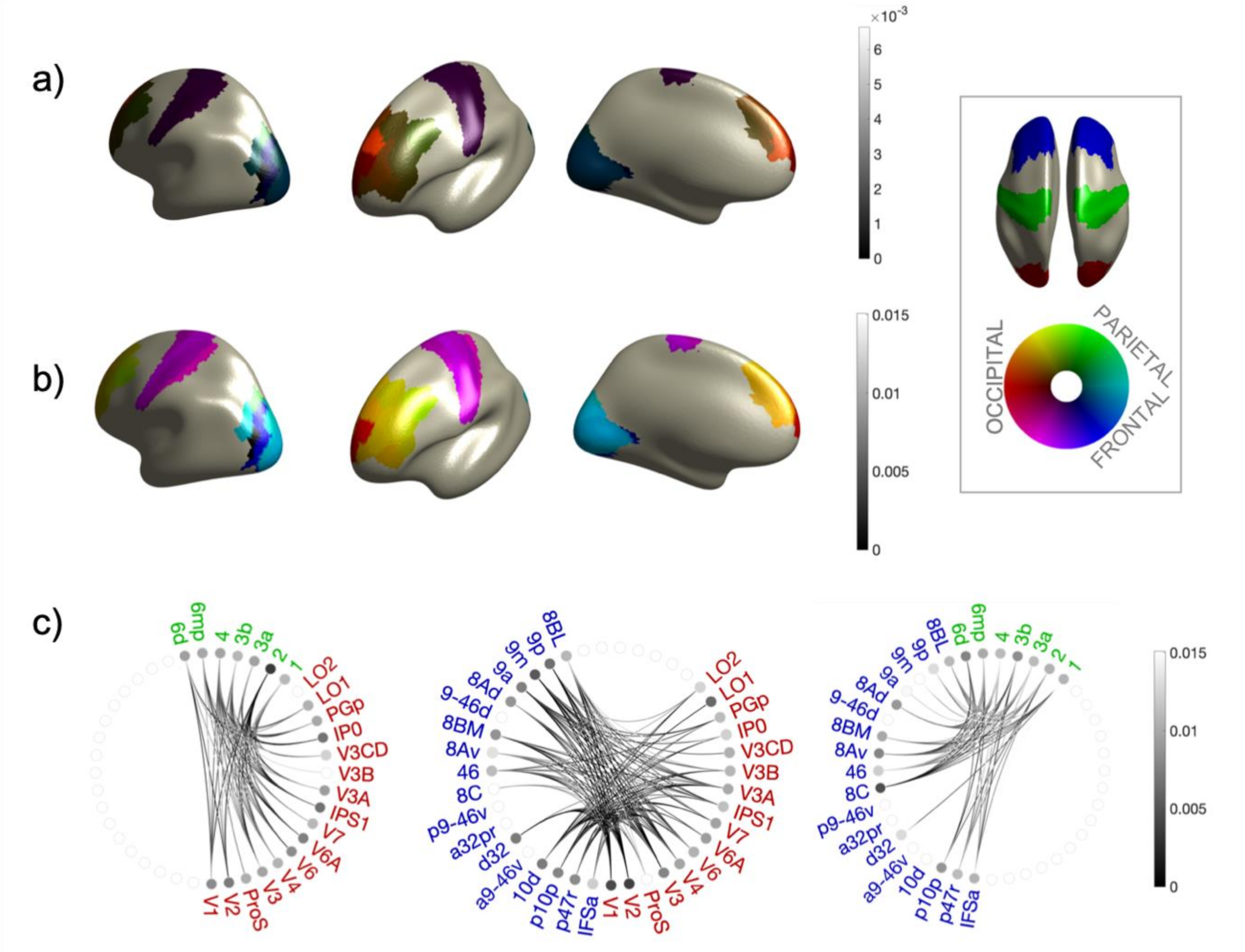
2.6. Connectivity Analysis and Group Statistical Validation
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berger, H. Über das elektroenkephalogramm des menschen. Arch. Psychiatr. Nervenkrankh. 1929, 87, 527–570. [Google Scholar] [CrossRef]
- Da Silva, F.L.; Vos, J.E.; Mooibroek, J.; Van Rotterdam, A. Relative contributions of intracortical and thalamo-cortical processes in the generation of alpha rhythms, revealed by partial coherence analysis. Electroencephalogr. Clin. Neurophysiol. 1980, 50, 449–456. [Google Scholar] [CrossRef]
- Vijayan, S.; Kopell, N.J. Thalamic model of awake alpha oscillations and implications for stimulus processing. Proc. Natl. Acad. Sci. USA 2012, 109, 18553–18558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolk, A.; Brinkman, L.; Vansteensel, M.J.; Aarnoutse, E.; Leijten, F.S.; Dijkerman, C.H.; Knight, R.T.; de Lange, F.P.; Toni, I. Electrocorticographic dissociation of alpha and beta rhythmic activity in the human sensorimotor system. eLife 2019, 8, e48065. [Google Scholar] [CrossRef]
- Klimesch, W.; Sauseng, P.; Hanslmayr, S. EEG alpha oscillations: The inhibition–timing hypothesis. Brain Res. Rev. 2007, 53, 63–88. [Google Scholar] [CrossRef]
- Jensen, O.; Mazaheri, A. Shaping functional architecture by oscillatory alpha activity: Gating by inhibition. Front. Hum. Neurosci. 2010, 4, 186. [Google Scholar] [CrossRef] [Green Version]
- Mathewson, K.E.; Gratton, G.; Fabiani, M.; Beck, D.M.; Ro, T. To see or not to see: Prestimulus α phase predicts visual awareness. J. Neurosci. 2009, 29, 2725–2732. [Google Scholar] [CrossRef] [Green Version]
- Zrenner, C.; Desideri, D.; Belardinelli, P.; Ziemann, U. Real-time EEG-defined excitability states determine efficacy of TMS-induced plasticity in human motor cortex. Brain Stimul. 2018, 11, 374–389. [Google Scholar] [CrossRef]
- Bergmann, T.O.; Lieb, A.; Zrenner, C.; Ziemann, U. Pulsed facilitation of corticospinal excitability by the sensorimotor μ-alpha rhythm. J. Neurosci. 2019, 39, 10034–10043. [Google Scholar] [CrossRef] [Green Version]
- Desideri, D.; Zrenner, C.; Ziemman, U.; Belardinelli, P. Phase of sensorimotor μ-oscillation modulates cortical responses to transcranial magnetic stimulation of the human motor cortex. J. Physiol. 2019, 597, 5671–5686. [Google Scholar] [CrossRef]
- Mazaheri, A.; Jensen, O. Rhythmic pulsing: Linking ongoing brain activity with evoked responses. Front. Hum. Neurosci. 2010, 4, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dugué, L.; Marque, P.; VanRullen, R. The phase of ongoing oscillations mediates the causal relation between brain excitation and visual perception. J. Neurosci. 2011, 31, 11889–11893. [Google Scholar] [CrossRef] [PubMed]
- Stefanou, M.I.; Desideri, D.; Belardinelli, P.; Zrenner, C.; Ziemann, U. Phase synchronicity of μ-rhythm determines efficacy of interhemispheric communication between human motor cortices. J. Neurosci. 2018, 38, 10525–10534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaworonkow, N.; Gordon, P.C.; Belardinelli, P.; Ziemann, U.; Bergmann, T.O.; Zrenner, C. μ-rhythm extracted with personalized EEG filters correlates with corticospinal excitability in real-time phase-triggered EEG-TMS. Front. Neurosci. 2018, 12, 954. [Google Scholar] [CrossRef]
- Zrenner, B.; Zrenner, C.; Gordon, P.C.; Belardinelli, P.; McDermott, E.J.; Soekadar, S.R.; Fallgatter, A.J.; Ziemann, U.; Müller-Dahlhaus, F. Brain oscillation-synchronized stimulation of the left dorsolateral prefrontal cortex in depression using real-time EEG-triggered TMS. Brain Stimul. 2020, 13, 197–205. [Google Scholar] [CrossRef] [Green Version]
- Gordon, P.C.; Dörre, S.; Belardinelli, P.; Stenroos, M.; Zrenner, B.; Ziemann, U.; Zrenner, C. Prefrontal theta-phase synchronized brain stimulation with real-time EEG-Triggered TMS. Front. Hum. Neurosci. 2021, 335. [Google Scholar] [CrossRef]
- Fries, P. Rhythms for cognition: Communication through coherence. Neuron 2015, 88, 220–235. [Google Scholar] [CrossRef] [Green Version]
- Nieminen, J.O.; Sinisalo, H.; Souza, V.H.; Malmi, M.; Yuryev, M.; Tervo, A.E.; Stenroos, M.; Milardovich, D.; Korhonen, J.T.; Koponen, L.M.; et al. Multi-locus transcranial magnetic stimulation system for electronically targeted brain stimulation. Brain Stimul. 2022, 15, 116–124. [Google Scholar] [CrossRef]
- Vinck, M.; van Wingerden, M.; Womelsdorf, T.; Fries, P.; Pennartz, C.M.A. The pairwise phase consistency: A bias-free measure of rhythmic neuronal synchronization. Neuroimage 2010, 51, 112–122. [Google Scholar] [CrossRef]
- Vinck, M.; Oostenveld, R.; van Wingerden, M.; Battaglia, F.; Pennartz, C.M.A. An improved index of phase-synchronization for electrophysiological data in the presence of volume-conduction, noise and sample-size bias. Neuroimage 2011, 55, 1548–1565. [Google Scholar] [CrossRef]
- Bastos, A.M.; Schoffelen, J.M. A tutorial review of functional connectivity analysis methods and their interpretational pitfalls. Front. Syst. Neurosci. 2016, 9, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, J.; Kluger, D.S.; Abbasi, O.; Chalas, N.; Steingräber, N.; Daube, C.; Schoffelen, J.-M. Comparison of undirected frequency-domain connectivity measures for cerebro-peripheral analysis. Neuroimage 2021, 245, 118660. [Google Scholar] [CrossRef] [PubMed]
- Pezzulo, G.; Zorzi, M.; Corbetta, M. The secret life of predictive brains: What’s spontaneous activity for? Trends Cogn. Sci. 2021, 25, 730–743. [Google Scholar] [CrossRef] [PubMed]
- Folstein, M.F.; Folstein, S.E.; McHugh, P.R. ‘Mini-mental state’ a practicalmethod for grading the cognitive state of patients for the clinician. J. Psychiatr. Res. 1975, 12, 189–198. [Google Scholar] [CrossRef]
- Taylor, J.R.; Williams, N.; Cusack, R.; Auer, T.; Shafto, M.A.; Dixon, M.; Tyler, L.K.; Henson, R.N. The Cambridge Centre for ageing and neuroscience (Cam-CAN) data repository: Structural and functional MRI, MEG, and cognitive data from a cross-sectional adult lifespan sample. Neuroimage 2017, 144, 262–269. [Google Scholar]
- Oostenveld, R.; Fries, P.; Maris, E.; Schoffelen, J.-M. FieldTrip: Open source software for advanced analysis of MEG, EEG, and invasive electrophysiological data. Comput. Intell. Neurosci. 2011, 2011, 868305. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J. Statistical Parametric Mapping: The Analysis of Funtional Brain Images, 1st ed.; Elsevier: Amsterdam, The Netherlands; Academic Press: Boston, MA, USA, 2007. [Google Scholar]
- Taulu, S.; Simola, J. Spatiotemporal signal space separation method for rejecting nearby interference in MEG measurements. Phys. Med. Biol. 2006, 51, 1759–1768. [Google Scholar] [CrossRef]
- Taulu, S.; Simola, J.; Kajola, M. Applications of the signal space separation method. IEEE Trans. Signal Process. 2005, 53, 3359–3372. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.-W.; Girolami, M.; Sejnowski, T.J. Independent component analysis using an extended infomax algorithm for mixed subgaussian and supergaussian sources. Neural Comput. 1999, 11, 417–441. [Google Scholar] [CrossRef]
- Hyvärinen, A.; Karhunen, J.; Oja, E. Independent Component Analysis; Wiley: Hoboken, NJ, USA, 2001. [Google Scholar]
- Fischl, B.; Sereno, M.I.; Tootell, R.B.H.; Dale, A.M. High-resolution intersubject averaging and a coordinate system for the cortical surface. Hum. Brain Mapp. 1999, 8, 272–284. [Google Scholar] [CrossRef]
- Glasser, M.F.; Coalson, T.S.; Robinson, E.C.; Hacker, C.D.; Harwell, J.; Yacoub, E.; Ugurbil, K.; Andersson, J.; Beckmann, C.F.; Jenkinson, M.; et al. A multi-modal parcellation of human cerebral cortex. Nature 2016, 536, 171–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilmoniemi, R.; Sarvas, J. Brain Signals: Physics and Mathematics of MEG and EEG; MIT Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Hämäläinen, M.; Hari, R.; Ilmoniemi, R.J.; Knuutila, J.; Lounasmaa, O.V. Magnetoencephalography—Theory, instrumentation, and applications to noninvasive studies of the working human brain. Rev. Mod. Phys. 1993, 65, 413–497. [Google Scholar] [CrossRef] [Green Version]
- Gross, J.; Kujala, J.; Hamalainen, M.; Timmermann, L.; Schnitzler, A.; Salmelin, R. Dynamic imaging of coherent sources: Studying neural interactions in the human brain. Proc. Natl. Acad. Sci. USA 2001, 98, 694–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markett, S.; Reuter, M.; Montag, C.; Voigt, G.; Lachmann, B.; Rudorf, S.; Elger, C.E.; Weber, B. Assessing the function of the fronto-parietal attention network: Insights from resting-state fMRI and the attentional network test. Hum. Brain Mapp. 2014, 35, 1700–1709. [Google Scholar] [CrossRef]
- Capotosto, P.; Babiloni, C.; Romani, G.L.; Corbetta, M. Frontoparietal cortex controls spatial attention through modulation of anticipatory alpha rhythms. J. Neurosci. 2009, 29, 5863–5872. [Google Scholar] [CrossRef] [Green Version]
- Hamm, J.P.; Dyckman, K.A.; McDowell, J.E.; Clementz, B.A. Pre-cue fronto-occipital alpha phase and distributed cortical oscillations predict failures of cognitive control. J. Neurosci. 2012, 32, 7034–7041. [Google Scholar] [CrossRef] [Green Version]
- Brancaccio, A.; Tabarelli, D.; Belardinelli, P. A new framework to interpret individual inter-hemispheric compensatory communication after stroke. J. Pers. Med. 2022, 12, 59. [Google Scholar] [CrossRef]
- Fox, M.D.; Corbetta, M.; Snyder, A.Z.; Vincent, J.L.; Raichle, M.E. Spontaneous neuronal activity distinguishes human dorsal and ventral attention systems. Proc. Natl. Acad. Sci. USA 2006, 103, 10046–10051. [Google Scholar] [CrossRef] [Green Version]
- Mantini, D.; Perrucci, M.G.; Del Gratta, C.; Romani, G.L.; Corbetta, M. Electrophysiological signatures of resting state networks in the human brain. Proc. Natl. Acad. Sci. USA 2007, 104, 13170–13175. [Google Scholar] [CrossRef] [Green Version]
- Smitha, K.A.; Akhil Raja, K.; Arun, K.M.; Rajesh, P.G.; Thomas, B.; Kapilamoorthy, T.R.; Kesavadas, C. Resting state fMRI: A review on methods in resting state connectivity analysis and resting state networks. Neuroradiol. J. 2017, 30, 305–317. [Google Scholar] [CrossRef]
- Zrenner, C.; Belardinelli, P.; Müller-Dahlhaus, F.; Ziemann, U. Closed-loop neuroscience and non-invasive brain stimulation: A tale of two loops. Front. Cell. Neurosci. 2016, 10, 92. [Google Scholar] [CrossRef] [PubMed]
- Momi, D.; Ozdemir, R.A.; Tadayon, E.; Boucher, P.; Shafi, M.M.; Pascual-Leone, A.; Santarnecchi, E. Network-level macroscale structural connectivity predicts propagation of transcranial magnetic stimulation. Neuroimage 2021, 229, 117698. [Google Scholar] [CrossRef] [PubMed]
- Maris, E.; Fries, P.; van Ede, F. Diverse phase relations among neuronal rhythms and their potential function. Trends Neurosci. 2016, 39, 86–99. [Google Scholar] [CrossRef] [PubMed]
- Ermolova, M.; Metsomaa, J.; Zrenner, C.; Kozák, G.; Marzetti, L.; Ziemann, U. Spontaneous phase-coupling within cortico-cortical networks: How time counts for brain-state-dependent stimulation. Brain Stimul. 2021, 14, 404–406. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| ROI | Sector | MNI Coordinates of Centroid (mm) | ||
|---|---|---|---|---|
| x | y | z | ||
| V1 | Occipital | −13.1 | −82.0 | 1.5 |
| V2 | Occipital | −12.4 | −81.5 | 3.6 |
| ProS | Occipital | −18.5 | −52.2 | 0.1 |
| V3 | Occipital | −18.3 | −86.2 | 5.4 |
| V4 | Occipital | −29.7 | −82.5 | −3.9 |
| V6 | Occipital | −13.9 | −78.0 | 27.2 |
| V6A | Occipital | −18.6 | −84.3 | 38.1 |
| V7 | Occipital | −23.8 | −81.9 | 26.6 |
| IPS1 | Occipital | −22.6 | −71.7 | 33.0 |
| V3A | Occipital | −17.2 | −88.4 | 23.0 |
| V3B | Occipital | −28.2 | −78.9 | 16.3 |
| V3CD | Occipital | −35.3 | −85.7 | 12.3 |
| IP0 | Occipital | −30.4 | −73.5 | 25.5 |
| PGp | Occipital | −39.8 | −80.1 | 22.1 |
| LO1 | Occipital | −37.8 | −82.9 | 4.2 |
| LO2 | Occipital | −42.7 | −83.3 | −4.9 |
| 1 | Parietal | −47.1 | −24.5 | 52.3 |
| 2 | Parietal | −35.4 | −34.4 | 49.7 |
| 3a | Parietal | −34.3 | −21.8 | 41.8 |
| 3b | Parietal | −36.8 | −24.1 | 51.6 |
| 4 | Parietal | −26.7 | −19.7 | 53.8 |
| 6mp | Parietal | −14.1 | −13.2 | 65.7 |
| 6d | Parietal | −34.9 | −12.7 | 61.9 |
| 8BL | Frontal | −11.6 | 35.1 | 50.8 |
| 9p | Frontal | −18.9 | 44.0 | 36.4 |
| 9m | Frontal | −7.7 | 51.0 | 21.8 |
| 9a | Frontal | −19.7 | 53.2 | 23.8 |
| 8Ad | Frontal | −23.3 | 24.7 | 41.2 |
| 9–46d | Frontal | −28.7 | 42.1 | 21.4 |
| 8BM | Frontal | −6.3 | 29.5 | 43.1 |
| 8Av | Frontal | −37.1 | 18.0 | 47.4 |
| 46 | Frontal | −36.6 | 35.6 | 28.3 |
| 8C | Frontal | −40.3 | 16.1 | 35.0 |
| p9–46v | Frontal | −43.3 | 29.2 | 26.3 |
| a32pr | Frontal | −10.2 | 28.1 | 28.6 |
| d32 | Frontal | −10.0 | 38.5 | 21.1 |
| a9–46v | Frontal | −37.1 | 47.7 | 8.8 |
| 10d | Frontal | −12.1 | 62.9 | 8.4 |
| p10p | Frontal | −23.6 | 55.0 | 5.2 |
| p47r | Frontal | −41.2 | 40.3 | 1.5 |
| IFSa | Frontal | −42.0 | 31.2 | 13.2 |
| Occipital | Parietal | Frontal | ||||
|---|---|---|---|---|---|---|
| V1 | 0.00005 | 0.01813 | 0.04801 | 0.03330 | ||
| V2 | 0.00026 | 0.02336 | 0.03238 | 0.03052 | ||
| ProS | ||||||
| V3 | 0.00054 | 0.02260 | 0.01536 | 0.02715 | ||
| V4 | 0.00007 | 0.01750 | 0.00167 | 0.01769 | ||
| V6 | 0.01140 | 0.03235 | 0.01202 | 0.02595 | ||
| V6A | 0.00591 | 0.02071 | 0.01167 | 0.01895 | ||
| V7 | 0.00787 | 0.02024 | 0.01866 | 0.02350 | ||
| IPS1 | 0.01689 | 0.03146 | 0.00290 | 0.01733 | ||
| V3A | 0.00240 | 0.02260 | 0.01683 | 0.02353 | ||
| V3B | 0.00526 | 0.02268 | 0.00203 | 0.01546 | ||
| V3CD | 0.00017 | 0.01639 | 0.00426 | 0.01660 | ||
| IP0 | 0.00256 | 0.02089 | 0.00459 | 0.01869 | ||
| PGp | 0.00036 | 0.01662 | 0.00583 | 0.01964 | ||
| LO1 | 0.00054 | 0.01236 | ||||
| LO2 | 0.00007 | 0.01632 | 0.00008 | 0.01108 | ||
| 1 | 0.00147 | 0.01513 | 0.01000 | 0.02590 | ||
| 2 | 0.01323 | 0.03352 | 0.00150 | 0.02274 | ||
| 3a | 0.00182 | 0.02163 | 0.01169 | 0.06285 | ||
| 3b | 0.00166 | 0.01980 | 0.00928 | 0.04439 | ||
| 4 | 0.00153 | 0.01990 | 0.01354 | 0.06664 | ||
| 6mp | 0.00026 | 0.01459 | 0.00653 | 0.03106 | ||
| 6d | 0.00037 | 0.01261 | 0.02895 | 0.05629 | ||
| 8BL | 0.09001 | 0.02671 | 0.00231 | 0.01952 | ||
| 9p | 0.08669 | 0.02861 | 0.00182 | 0.01623 | ||
| 9m | 0.10542 | 0.03381 | 0.00006 | 0.01323 | ||
| 9a | 0.11954 | 0.02970 | 0.00015 | 0.01537 | ||
| 8Ad | 0.03460 | 0.01914 | 0.00374 | 0.01783 | ||
| 9–46d | 0.05946 | 0.02154 | 0.00038 | 0.01336 | ||
| 8BM | 0.04143 | 0.02319 | 0.00015 | 0.01470 | ||
| 8Av | 0.00697 | 0.01461 | 0.06058 | 0.04594 | ||
| 46 | 0.01625 | 0.01679 | 0.00248 | 0.01423 | ||
| 8C | 0.00071 | 0.01432 | 0.10138 | 0.05798 | ||
| p9–46v | 0.00158 | 0.01213 | 0.01557 | 0.01848 | ||
| a32pr | 0.00488 | 0.01591 | 0.00003 | 0.00649 | ||
| d32 | 0.02711 | 0.02249 | 0.00003 | 0.00840 | ||
| a9–46v | 0.04548 | 0.01925 | 0.00283 | 0.01968 | ||
| 10d | 0.07100 | 0.02592 | 0.00028 | 0.01980 | ||
| p10p | 0.05559 | 0.02151 | 0.00038 | 0.01635 | ||
| p47r | 0.01739 | 0.01740 | 0.01290 | 0.02046 | ||
| IFSa | 0.00425 | 0.01535 | 0.00776 | 0.01874 | ||
| Occipital | Parietal | Frontal | ||||
|---|---|---|---|---|---|---|
| V1 | 0.00004 | 0.01007 | 0.00103 | 0.01402 | ||
| V2 | 0.00002 | 0.01189 | 0.00103 | 0.01511 | ||
| ProS | 0.00008 | 0.00814 | ||||
| V3 | 0.00004 | 0.01173 | 0.00041 | 0.01337 | ||
| V4 | 0.00012 | 0.01177 | ||||
| V6 | 0.00007 | 0.01102 | 0.00013 | 0.01063 | ||
| V6A | 0.00014 | 0.00900 | 0.00013 | 0.01001 | ||
| V7 | 0.00019 | 0.01040 | ||||
| IPS1 | 0.00023 | 0.01053 | ||||
| V3A | 0.00005 | 0.01201 | 0.00014 | 0.01081 | ||
| V3B | 0.00016 | 0.01120 | ||||
| V3CD | ||||||
| IP0 | 0.00016 | 0.01141 | 0.00003 | 0.00919 | ||
| PGp | 0.00009 | 0.01005 | 0.00009 | 0.01133 | ||
| LO1 | ||||||
| LO2 | 0.00017 | 0.00917 | ||||
| 1 | 0.00010 | 0.00946 | 0.00006 | 0.01010 | ||
| 2 | 0.00022 | 0.01309 | 0.00006 | 0.01011 | ||
| 3a | 0.00008 | 0.01093 | 0.00022 | 0.01174 | ||
| 3b | 0.00013 | 0.01032 | 0.00011 | 0.01131 | ||
| 4 | 0.00015 | 0.01078 | 0.00017 | 0.01302 | ||
| 6mp | 0.00007 | 0.00970 | 0.00009 | 0.00931 | ||
| 6d | 0.00018 | 0.01071 | 0.00027 | 0.01096 | ||
| 8BL | 0.00266 | 0.01300 | 0.00008 | 0.01005 | ||
| 9p | 0.00664 | 0.01340 | 0.00006 | 0.00831 | ||
| 9m | 0.00505 | 0.01418 | 0.00003 | 0.00788 | ||
| 9a | 0.00261 | 0.01296 | ||||
| 8Ad | 0.00063 | 0.01018 | 0.00010 | 0.00953 | ||
| 9–46d | 0.00206 | 0.01211 | 0.00002 | 0.00974 | ||
| 8BM | 0.00133 | 0.01180 | 0.00002 | 0.00744 | ||
| 8Av | 0.00011 | 0.00886 | 0.00074 | 0.01142 | ||
| 46 | 0.00043 | 0.01072 | 0.00010 | 0.00949 | ||
| 8C | 0.00003 | 0.00789 | 0.00090 | 0.01296 | ||
| p9–46v | 0.00017 | 0.00939 | 0.00025 | 0.01076 | ||
| a32pr | 0.00051 | 0.01047 | 0.00002 | 0.00623 | ||
| d32 | 0.00134 | 0.01224 | 0.00003 | 0.00763 | ||
| a9–46v | 0.00119 | 0.01163 | 0.00005 | 0.00786 | ||
| 10d | 0.00269 | 0.01193 | ||||
| p10p | 0.00150 | 0.01111 | 0.00012 | 0.00804 | ||
| p47r | 0.00051 | 0.01068 | 0.00007 | 0.00797 | ||
| IFSa | 0.00029 | 0.00949 | 0.00011 | 0.00918 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tabarelli, D.; Brancaccio, A.; Zrenner, C.; Belardinelli, P. Functional Connectivity States of Alpha Rhythm Sources in the Human Cortex at Rest: Implications for Real-Time Brain State Dependent EEG-TMS. Brain Sci. 2022, 12, 348. https://doi.org/10.3390/brainsci12030348
Tabarelli D, Brancaccio A, Zrenner C, Belardinelli P. Functional Connectivity States of Alpha Rhythm Sources in the Human Cortex at Rest: Implications for Real-Time Brain State Dependent EEG-TMS. Brain Sciences. 2022; 12(3):348. https://doi.org/10.3390/brainsci12030348
Chicago/Turabian StyleTabarelli, Davide, Arianna Brancaccio, Christoph Zrenner, and Paolo Belardinelli. 2022. "Functional Connectivity States of Alpha Rhythm Sources in the Human Cortex at Rest: Implications for Real-Time Brain State Dependent EEG-TMS" Brain Sciences 12, no. 3: 348. https://doi.org/10.3390/brainsci12030348
APA StyleTabarelli, D., Brancaccio, A., Zrenner, C., & Belardinelli, P. (2022). Functional Connectivity States of Alpha Rhythm Sources in the Human Cortex at Rest: Implications for Real-Time Brain State Dependent EEG-TMS. Brain Sciences, 12(3), 348. https://doi.org/10.3390/brainsci12030348