
Prediction of Chronological Age in Healthy Elderly Subjects with Machine Learning from MRI Brain Segmentation and Cortical Parcellation


Abstract
:1. Introduction
2. Methods
2.1. Study Participants
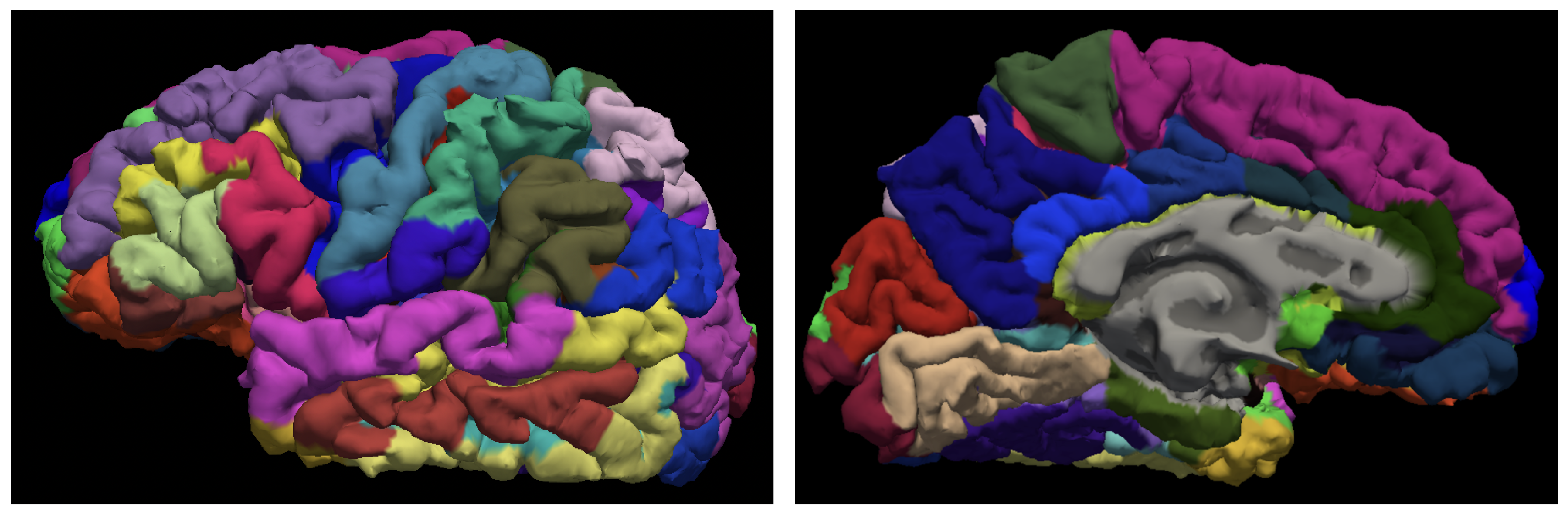
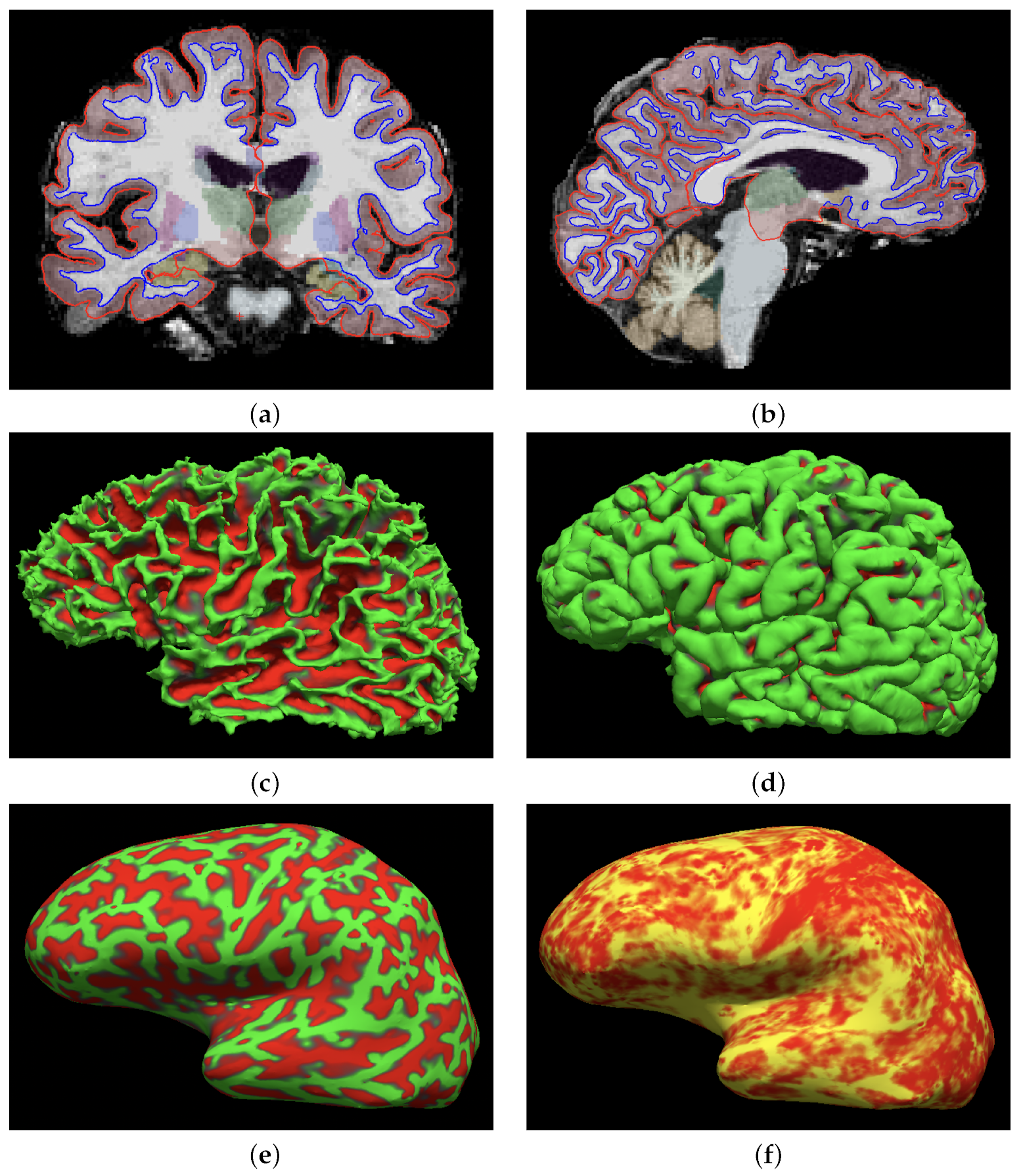
2.2. MRI Data Acquisition and Preprocessing
2.3. Anomaly Detection with the Isolation Forest Algorithm
2.4. Statistical Analysis
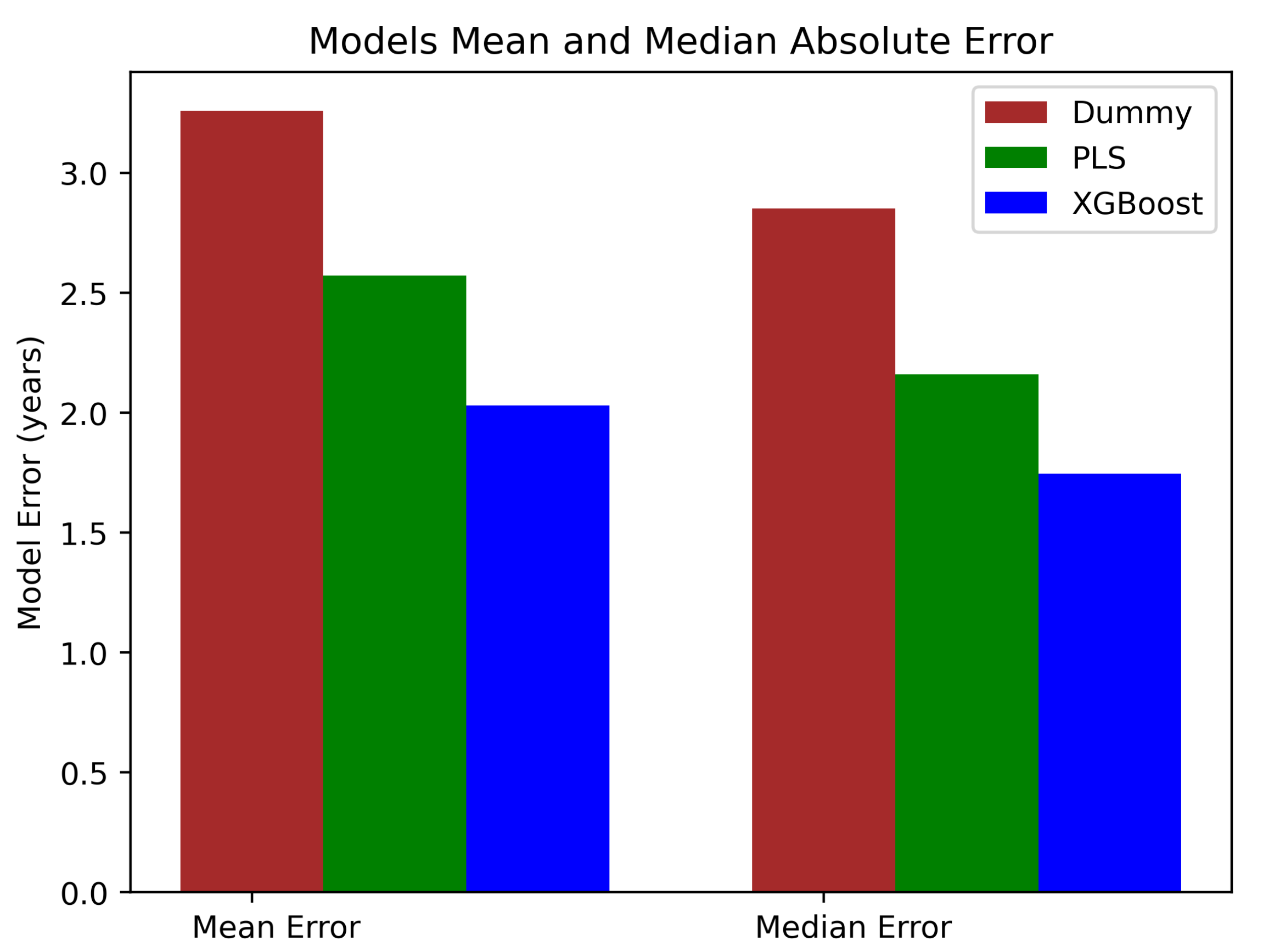
2.5. Age Prediction Analysis
3. Results
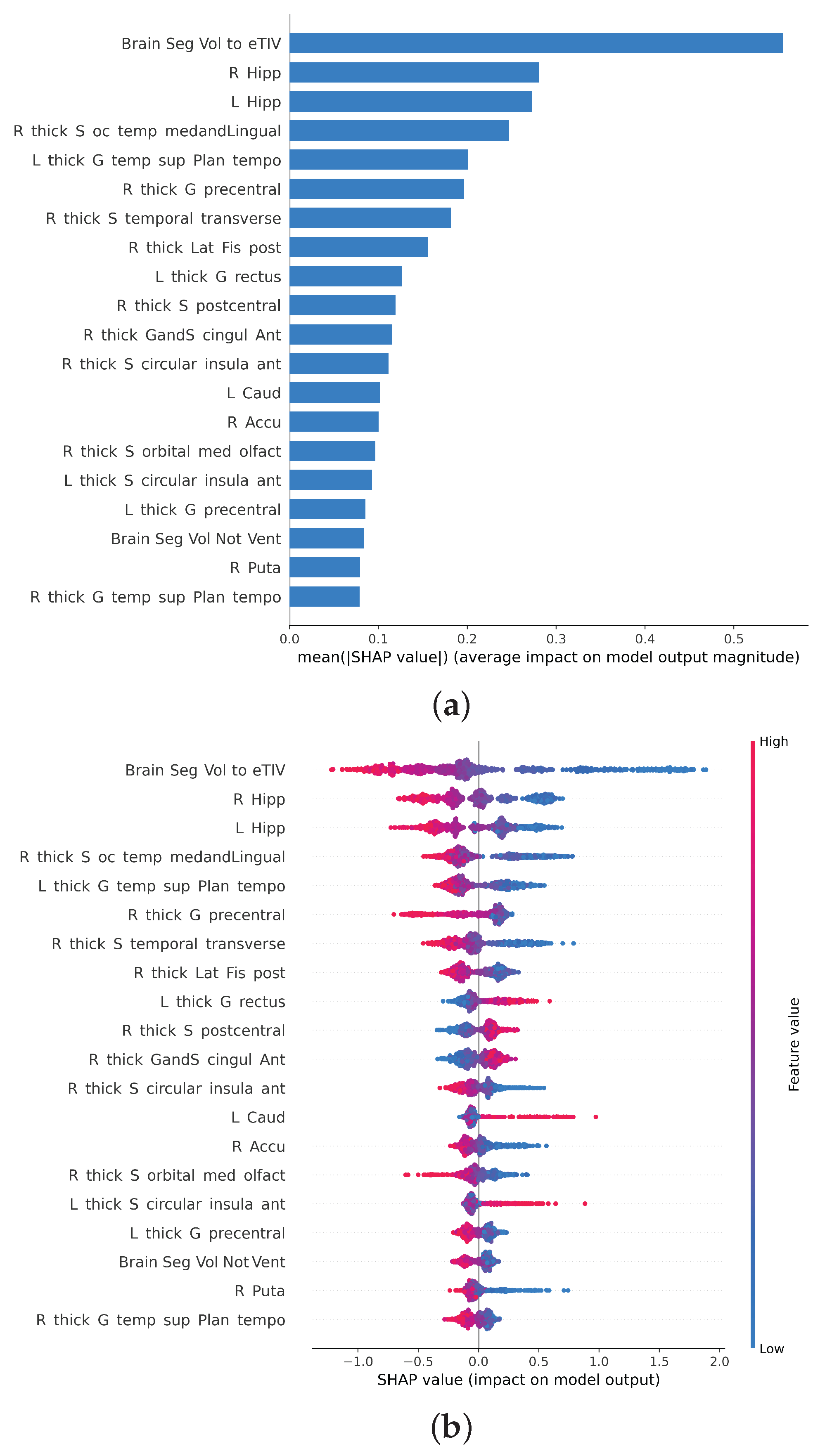
3.1. Feature Importance
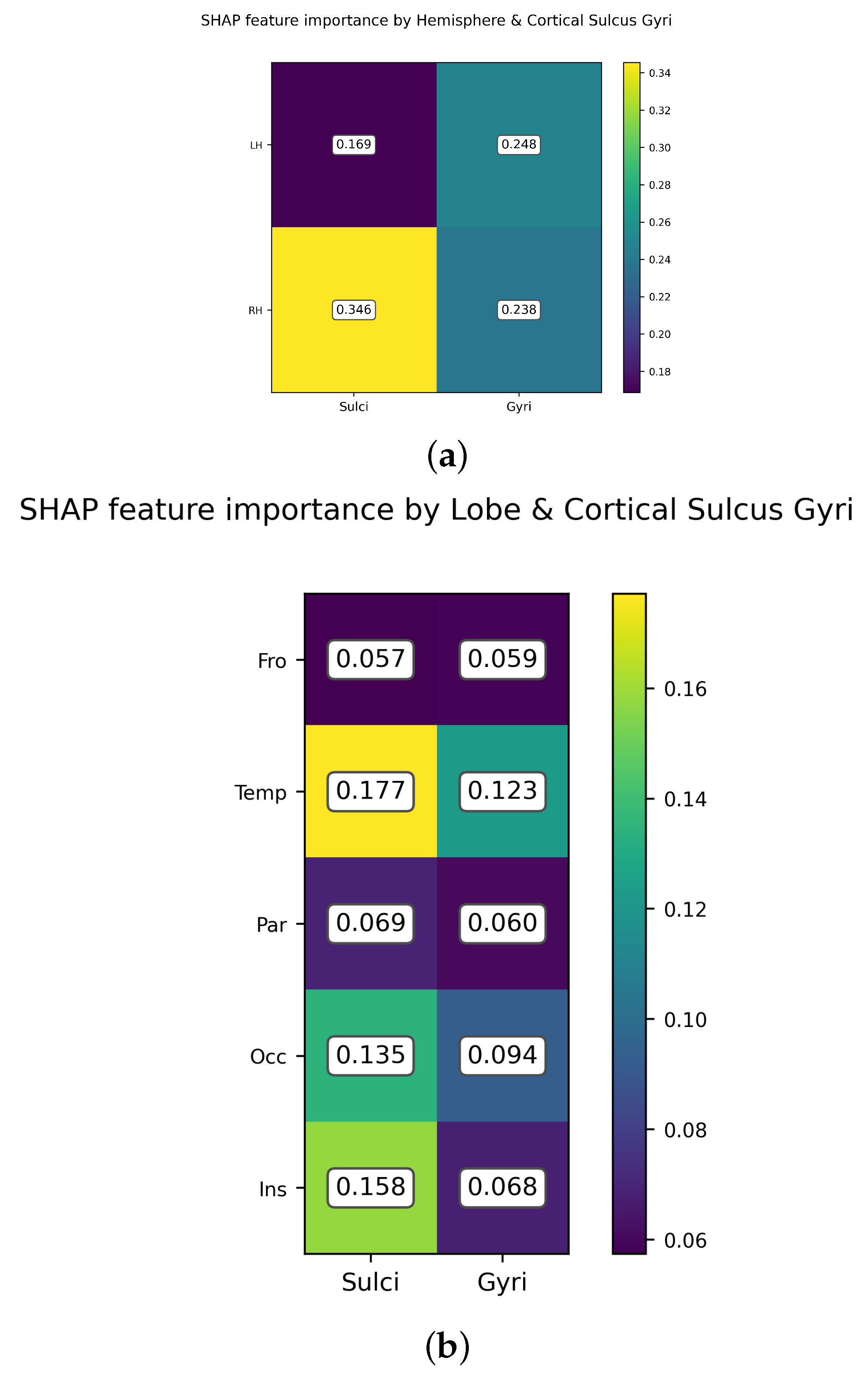
3.2. Feature Importance of Cortical Gyri and Sulci
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Granziera, C.; Wuerfel, J.; Barkhof, F.; Calabrese, M.; De Stefano, N.; Enzinger, C.; Evangelou, N.; Filippi, M.; Geurts, J.J.; Reich, D.S.; et al. Quantitative magnetic resonance imaging towards clinical application in multiple sclerosis. Brain 2021, 144, 1296–1311. [Google Scholar] [CrossRef] [PubMed]
- Harisinghani, M.G.; O’Shea, A.; Weissleder, R. Advances in clinical MRI technology. Sci. Transl. Med. 2019, 11, eaba2591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakkamakul, S.; Witzel, T.; Huang, S.; Boulter, D.; Borja, M.J.; Schaefer, P.; Rosen, B.; Heberlein, K.; Ratai, E.; Gonzalez, G.; et al. Ultrafast brain MRI: Clinical deployment and comparison to conventional brain MRI at 3T. J. Neuroimaging 2016, 26, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Duan, G.; Wu, K.; Anderson, S.W.; Zhang, X. Intelligent metamaterials based on nonlinearity for magnetic resonance imaging. Adv. Mater. 2019, 31, 1905461. [Google Scholar] [CrossRef]
- Gudigar, A.; Raghavendra, U.; Hegde, A.; Kalyani, M.; Ciaccio, E.J.; Acharya, U.R. Brain pathology identification using computer aided diagnostic tool: A systematic review. Comput. Methods Programs Biomed. 2020, 187, 105205. [Google Scholar] [CrossRef]
- Gauriau, R.; Bizzo, B.C.; Kitamura, F.C.; Landi Junior, O.; Ferraciolli, S.F.; Macruz, F.B.; Sanchez, T.A.; Garcia, M.R.; Vedolin, L.M.; Domingues, R.C.; et al. A Deep Learning-Based Model for Detecting Abnormalities on Brain MRI for Triaging: Preliminary Results from a Multi-Site Experience. Radiol. Artif. Intell. 2021, 3, e200184. [Google Scholar] [CrossRef]
- MacDonald, M.E.; Pike, G.B. MRI of healthy brain aging: A review. NMR Biomed. 2021, 3, e4564. [Google Scholar] [CrossRef]
- Royle, N.A.; Booth, T.; Hernández, M.C.V.; Penke, L.; Murray, C.; Gow, A.J.; Maniega, S.M.; Starr, J.; Bastin, M.E.; Deary, I.J.; et al. Estimated maximal and current brain volume predict cognitive ability in old age. Neurobiol. Aging 2013, 34, 2726–2733. [Google Scholar] [CrossRef] [Green Version]
- Salat, D.H.; Buckner, R.L.; Snyder, A.Z.; Greve, D.N.; Desikan, R.S.; Busa, E.; Morris, J.C.; Dale, A.M.; Fischl, B. Thinning of the cerebral cortex in aging. Cereb. Cortex 2004, 14, 721–730. [Google Scholar] [CrossRef] [Green Version]
- Bennett, I.J.; Greenia, D.E.; Maillard, P.; Sajjadi, S.A.; DeCarli, C.; Corrada, M.M.; Kawas, C.H. Age-related white matter integrity differences in oldest-old without dementia. Neurobiol. Aging 2017, 56, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Shams, S.; Martola, J.; Cavallin, L.; Granberg, T.; Shams, M.; Aspelin, P.; Wahlund, L.; Kristoffersen-Wiberg, M. SWI or T2*: Which MRI sequence to use in the detection of cerebral microbleeds? The Karolinska Imaging Dementia Study. Am. J. Neuroradiol. 2015, 36, 1089–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prins, N.D.; Scheltens, P. White matter hyperintensities, cognitive impairment and dementia: An update. Nat. Rev. Neurol. 2015, 11, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Grant, R.; Condon, B.; Lawrence, A.; Hadley, D.; Patterson, J.; Bone, I.; Teasdale, G. Human cranial CSF volumes measured by MRI: Sex and age influences. Magn. Reson. Imaging 1987, 5, 465–468. [Google Scholar] [CrossRef]
- Maruszak, A.; Thuret, S. Why looking at the whole hippocampus is not enough—a critical role for anteroposterior axis, subfield and activation analyses to enhance predictive value of hippocampal changes for Alzheimer’s disease diagnosis. Front. Cell. Neurosci. 2014, 8, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McRae-McKee, K.; Evans, S.; Hadjichrysanthou, C.; Wong, M.; De Wolf, F.; Anderson, R. Combining hippocampal volume metrics to better understand Alzheimer’s disease progression in at-risk individuals. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Whitwell, J.L.; Wiste, H.J.; Weigand, S.D.; Rocca, W.A.; Knopman, D.S.; Roberts, R.O.; Boeve, B.F.; Petersen, R.C.; Jack, C.R.; Initiative, A.D.N.; et al. Comparison of imaging biomarkers in the Alzheimer disease neuroimaging initiative and the Mayo Clinic Study of Aging. Arch. Neurol. 2012, 69, 614–622. [Google Scholar] [CrossRef] [Green Version]
- Nobis, L.; Manohar, S.G.; Smith, S.M.; Alfaro-Almagro, F.; Jenkinson, M.; Mackay, C.E.; Husain, M. Hippocampal volume across age: Nomograms derived from over 19,700 people in UK Biobank. NeuroImage Clin. 2019, 23, 101904. [Google Scholar] [CrossRef]
- Serbruyns, L.; Leunissen, I.; Huysmans, T.; Cuypers, K.; Meesen, R.L.; van Ruitenbeek, P.; Sijbers, J.; Swinnen, S.P. Subcortical volumetric changes across the adult lifespan: Subregional thalamic atrophy accounts for age-related sensorimotor performance declines. Cortex 2015, 65, 128–138. [Google Scholar] [CrossRef]
- Gomez-Ramirez, J.; Quilis-Sancho, J.; Fernandez-Blazquez, M.A. A Comparative Analysis of MRI Automated Segmentation of Subcortical Brain Volumes in a Large Dataset of Elderly Subjects. Neuroinformatics 2021, 1–10. [Google Scholar] [CrossRef]
- McGinnis, S.M.; Brickhouse, M.; Pascual, B.; Dickerson, B.C. Age-related changes in the thickness of cortical zones in humans. Brain Topogr. 2011, 24, 279–291. [Google Scholar] [CrossRef] [Green Version]
- Sowell, E.R.; Peterson, B.S.; Thompson, P.M.; Welcome, S.E.; Henkenius, A.L.; Toga, A.W. Mapping cortical change across the human life span. Nat. Neurosci. 2003, 6, 309. [Google Scholar] [CrossRef] [PubMed]
- Shaw, M.E.; Sachdev, P.S.; Anstey, K.J.; Cherbuin, N. Age-related cortical thinning in cognitively healthy individuals in their 60s: The PATH Through Life study. Neurobiol. Aging 2016, 39, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, M.; Li, Y.; Huang, Y.; Chen, X. A novel brain network construction method for exploring age-related functional reorganization. Comput. Intell. Neurosci. 2016, 2016, 2429691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jónsson, B.A.; Bjornsdottir, G.; Thorgeirsson, T.; Ellingsen, L.M.; Walters, G.B.; Gudbjartsson, D.; Stefansson, H.; Stefansson, K.; Ulfarsson, M. Brain age prediction using deep learning uncovers associated sequence variants. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mendes, S.L.; Pinaya, W.H.L.; Pan, P.; Sato, J.R. Estimating Gender and Age from Brain Structural MRI of Children and Adolescents: A 3D Convolutional Neural Network Multitask Learning Model. Comput. Intell. Neurosci. 2021, 2021, 5550914. [Google Scholar] [CrossRef]
- Aycheh, H.M.; Seong, J.K.; Shin, J.H.; Na, D.L.; Kang, B.; Seo, S.W.; Sohn, K.A. Biological brain age prediction using cortical thickness data: A large scale cohort study. Front. Aging Neurosci. 2018, 10, 252. [Google Scholar] [CrossRef]
- Destrieux, C.; Fischl, B.; Dale, A.; Halgren, E. Automatic parcellation of human cortical gyri and sulci using standard anatomical nomenclature. NeuroImage 2010, 53, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kondo, C.; Ito, K.; Wu, K.; Sato, K.; Taki, Y.; Fukuda, H.; Aoki, T. An age estimation method using brain local features for T1-weighted images. In Proceedings of the 2015 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Milan, Italy, 25–29 August 2015; pp. 666–669. [Google Scholar]
- Cherubini, A.; Caligiuri, M.E.; Péran, P.; Sabatini, U.; Cosentino, C.; Amato, F. Importance of multimodal MRI in characterizing brain tissue and its potential application for individual age prediction. IEEE J. Biomed. Health Inform. 2016, 20, 1232–1239. [Google Scholar] [CrossRef]
- MacDonald, M.E.; Williams, R.J.; Forkert, N.D.; Berman, A.J.; McCreary, C.R.; Frayne, R.; Pike, G.B. Interdatabase variability in cortical thickness measurements. Cereb. Cortex 2019, 29, 3282–3293. [Google Scholar] [CrossRef]
- Cole, J.H.; Ritchie, S.J.; Bastin, M.E.; Hernández, V.; Muñoz Maniega, S.; Royle, N.; Corley, J.; Pattie, A.; Harris, S.E.; Zhang, Q.; et al. Brain age predicts mortality. Mol. Psychiatry 2018, 23, 1385–1392. [Google Scholar] [CrossRef]
- Beheshti, I.; Nugent, S.; Potvin, O.; Duchesne, S. Bias-adjustment in neuroimaging-based brain age frameworks: A robust scheme. NeuroImage Clin. 2019, 24, 102063. [Google Scholar] [CrossRef] [PubMed]
- Ham, S.; Lee, S.J.V. Advances in transcriptome analysis of human brain aging. Exp. Mol. Med. 2020, 52, 1787–1797. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, D.G.; Nalls, M.A.; Gibbs, J.R.; Arepalli, S.; van der Brug, M.; Chong, S.; Moore, M.; Longo, D.L.; Cookson, M.R.; Traynor, B.J.; et al. Distinct DNA methylation changes highly correlated with chronological age in the human brain. Hum. Mol. Genet. 2011, 20, 1164–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, S.; Zhang, Y.; Langfelder, P.; Kahn, R.S.; Boks, M.P.; van Eijk, K.; van den Berg, L.H.; Ophoff, R.A. Aging effects on DNA methylation modules in human brain and blood tissue. Genome Biol. 2012, 13, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannum, G.; Guinney, J.; Zhao, L.; Zhang, L.; Hughes, G.; Sadda, S.; Klotzle, B.; Bibikova, M.; Fan, J.B.; Gao, Y.; et al. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol. Cell 2013, 49, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Sayed, N.; Huang, Y.; Nguyen, K.; Krejciova-Rajaniemi, Z.; Grawe, A.P.; Gao, T.; Tibshirani, R.; Hastie, T.; Alpert, A.; Cui, L.; et al. An inflammatory aging clock (iAge) based on deep learning tracks multimorbidity, immunosenescence, frailty and cardiovascular aging. Nat. Aging 2021, 1, 598–615. [Google Scholar] [CrossRef] [PubMed]
- Shapley, L.S. A value for n-person games. Contrib. Theory Games 1953, 2, 307–317. [Google Scholar]
- Gómez-Ramírez, J.; Ávila-Villanueva, M.; Fernández-Blázquez, M.Á. Selecting the most important self-assessed features for predicting conversion to Mild Cognitive Impairment with Random Forest and Permutation-based methods. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Blasco, R.; Ruiz-Sánchez de León, J.M.; Ávila-Villanueva, M.; Valentí-Soler, M.; Gómez-Ramírez, J.; Fernández-Blázquez, M.A. Transition from mild cognitive impairment to normal cognition: Determining the predictors of reversion with multi-state Markov models. Alzheimer Dement. 2021. [Google Scholar] [CrossRef]
- McCarthy, C.S.; Ramprashad, A.; Thompson, C.; Botti, J.A.; Coman, I.L.; Kates, W.R. A comparison of FreeSurfer-generated data with and without manual intervention. Front. Neurosci. 2015, 9, 379. [Google Scholar] [CrossRef]
- Fischl, B. FreeSurfer. NeuroImage 2012, 62, 774–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Ramírez, J.; González-Rosa, J.J. Intra-and interhemispheric symmetry of subcortical brain structures: A volumetric analysis in the aging human brain. Brain Struct. Funct. 2022, 227, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Breiman, L. Bagging predictors. Mach. Learn. 1996, 24, 123–140. [Google Scholar] [CrossRef] [Green Version]
- Domingues, R.; Filippone, M.; Michiardi, P.; Zouaoui, J. A comparative evaluation of outlier detection algorithms: Experiments and analyses. Pattern Recognit. 2018, 74, 406–421. [Google Scholar] [CrossRef]
- Alaverdyan, Z. Unsupervised Representation Learning for Anomaly Detection on Neuroimaging. Application to Epilepsy Lesion Detection on Brain MRI. Ph.D. Thesis, Université de Lyon, Lyon, France, 2019. [Google Scholar]
- Shen, L.; Saykin, A.J.; Kim, S.; Firpi, H.A.; West, J.D.; Risacher, S.L.; McDonald, B.C.; McHugh, T.L.; Wishart, H.A.; Flashman, L.A. Comparison of manual and automated determination of hippocampal volumes in MCI and early AD. Brain Imaging Behav. 2010, 4, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Malone, I.B.; Leung, K.K.; Clegg, S.; Barnes, J.; Whitwell, J.L.; Ashburner, J.; Fox, N.C.; Ridgway, G.R. Accurate automatic estimation of total intracranial volume: A nuisance variable with less nuisance. NeuroImage 2015, 104, 366–372. [Google Scholar] [CrossRef] [Green Version]
- Wolf, H.; Julin, P.; Gertz, H.J.; Winblad, B.; Wahlund, L.O. Intracranial volume in mild cognitive impairment, Alzheimer’s disease and vascular dementia: Evidence for brain reserve? Int. J. Geriatr. Psychiatry 2004, 19, 995–1007. [Google Scholar] [CrossRef]
- Caspi, Y.; Brouwer, R.M.; Schnack, H.G.; van de Nieuwenhuijzen, M.E.; Cahn, W.; Kahn, R.S.; Niessen, W.J.; van der Lugt, A.; Pol, H.H. Changes in the intracranial volume from early adulthood to the sixth decade of life: A longitudinal study. NeuroImage 2020, 220, 116842. [Google Scholar] [CrossRef]
- Heinen, R.; Bouvy, W.H.; Mendrik, A.M.; Viergever, M.A.; Biessels, G.J.; De Bresser, J. Robustness of automated methods for brain volume measurements across different MRI field strengths. PLoS ONE 2016, 11, e0165719. [Google Scholar]
- Gómez-Ramírez, J.; Fernandez-Blazquez, M.A.; González-Rosa, J. The aging human brain: A causal analysis of the effect of sex and age on brain volume. bioRxiv 2020. [Google Scholar] [CrossRef]
- Garthwaite, P.H. An interpretation of partial least squares. J. Am. Stat. Assoc. 1994, 89, 122–127. [Google Scholar] [CrossRef]
- Chen, T.; He, T.; Benesty, M.; Khotilovich, V.; Tang, Y.; Cho, H. Xgboost: Extreme Gradient Boosting; R Package Version 0.4-2, 2015; Volume 1, pp. 1–4. [Google Scholar]
- Ritzberger, K. Foundations of Non-Cooperative Game Theory; OUP Catalogue; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Tripathi, S.; Hemachandra, N.; Trivedi, P. Interpretable feature subset selection: A Shapley value based approach. In Proceedings of the 2020 IEEE International Conference on Big Data (Big Data), Atlanta, GA, USA, 10–13 December 2020; pp. 5463–5472. [Google Scholar]
- Leonard, C.M.; Towler, S.; Welcome, S.; Halderman, L.K.; Otto, R.; Eckert, M.A.; Chiarello, C. Size matters: Cerebral volume influences sex differences in neuroanatomy. Cereb. Cortex 2008, 18, 2920–2931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lotze, M.; Domin, M.; Gerlach, F.H.; Gaser, C.; Lueders, E.; Schmidt, C.O.; Neumann, N. Novel findings from 2838 adult brains on sex differences in gray matter brain volume. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef]
- Mole, J.P.; Fasano, F.; Evans, J.; Sims, R.; Kidd, E.; Aggleton, J.P.; Metzler-Baddeley, C. APOE-ε4-related differences in left thalamic microstructure in cognitively healthy adults. Sci. Rep. 2020, 10, 1–25. [Google Scholar] [CrossRef]
- Fleisher, A.; Grundman, M.; Clifford, R.J., Jr.; Petersen, R.C.; Taylor, C.; Kim, H.T.; Schiller, D.H.B.; Bagwell, V.; Sencakova, D.; Weiner, M.F.; et al. Sex, Apolipoprotein E e4 Status, and Hippocampal Volume in Mild Cognitive Impairment. Arch. Neurol. 2005, 62, 953–957. [Google Scholar] [CrossRef] [Green Version]
- Orihashi, R.; Mizoguchi, Y.; Imamura, Y.; Yamada, S.; Ueno, T.; Monji, A. Oxytocin and elderly MRI-based hippocampus and amygdala volume: A 7-year follow-up study. Brain Commun. 2020, 2, fcaa081. [Google Scholar] [CrossRef]
- Bergstra, J.; Bengio, Y. Random search for hyper-parameter optimization. J. Mach. Learn. Res. 2012, 13, 281–305. [Google Scholar]
- Franke, K.; Ziegler, G.; Klöppel, S.; Gaser, C.; Initiative, A.D.N. Estimating the age of healthy subjects from T1-weighted MRI scans using kernel methods: Exploring the influence of various parameters. NeuroImage 2010, 50, 883–892. [Google Scholar] [CrossRef]
- Wang, J.; Li, W.; Miao, W.; Dai, D.; Hua, J.; He, H. Age estimation using cortical surface pattern combining thickness with curvatures. Med Biol. Eng. Comput. 2014, 52, 331–341. [Google Scholar] [CrossRef]
- Mwangi, B.; Hasan, K.M.; Soares, J.C. Prediction of individual subject’s age across the human lifespan using diffusion tensor imaging: A machine learning approach. NeuroImage 2013, 75, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Dosenbach, N.U.; Nardos, B.; Cohen, A.L.; Fair, D.A.; Power, J.D.; Church, J.A.; Nelson, S.M.; Wig, G.S.; Vogel, A.C.; Lessov-Schlaggar, C.N.; et al. Prediction of individual brain maturity using fMRI. Science 2010, 329, 1358–1361. [Google Scholar] [CrossRef] [Green Version]
- Brown, T.T.; Kuperman, J.M.; Chung, Y.; Erhart, M.; McCabe, C.; Hagler, D.J., Jr.; Venkatraman, V.K.; Akshoomoff, N.; Amaral, D.G.; Bloss, C.S.; et al. Neuroanatomical assessment of biological maturity. Curr. Biol. 2012, 22, 1693–1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Ramirez, J.; Wu, J. Network-based biomarkers in Alzheimer’s disease: Review and future directions. Front. Aging Neurosci. 2014, 6, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckerström, C.; Klasson, N.; Olsson, E.; Selnes, P.; Rolstad, S.; Wallin, A. Similar pattern of atrophy in early-and late-onset Alzheimer’s disease. Alzheimer Dementia Diagn. Assess. Dis. Monit. 2018, 10, 253–259. [Google Scholar] [CrossRef]
- Kaye, J.A.; Swihart, T.; Howieson, D.; Dame, A.; Moore, M.; Karnos, T.; Camicioli, R.; Ball, M.; Oken, B.; Sexton, G. Volume loss of the hippocampus and temporal lobe in healthy elderly persons destined to develop dementia. Neurology 1997, 48, 1297–1304. [Google Scholar] [CrossRef] [PubMed]
- Bettio, L.E.; Rajendran, L.; Gil-Mohapel, J. The effects of aging in the hippocampus and cognitive decline. Neurosci. Biobehav. Rev. 2017, 79, 66–86. [Google Scholar] [CrossRef]
- Schultz, M.B.; Kane, A.E.; Mitchell, S.J.; MacArthur, M.R.; Warner, E.; Vogel, D.S.; Mitchell, J.R.; Howlett, S.E.; Bonkowski, M.S.; Sinclair, D.A. Age and life expectancy clocks based on machine learning analysis of mouse frailty. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Niu, X.; Zhang, F.; Kounios, J.; Liang, H. Improved prediction of brain age using multimodal neuroimaging data. Hum. Brain Mapp. 2020, 41, 1626–1643. [Google Scholar] [CrossRef]
- Smith, S.M.; Vidaurre, D.; Alfaro-Almagro, F.; Nichols, T.E.; Miller, K.L. Estimation of brain age delta from brain imaging. NeuroImage 2019, 200, 528–539. [Google Scholar] [CrossRef]
- Pardoe, H.R.; Kuzniecky, R. NAPR: A cloud-based framework for neuroanatomical age prediction. Neuroinformatics 2018, 16, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Schnack, H.G.; Van Haren, N.E.; Nieuwenhuis, M.; Hulshoff Pol, H.E.; Cahn, W.; Kahn, R.S. Accelerated brain aging in schizophrenia: A longitudinal pattern recognition study. Am. J. Psychiatry 2016, 173, 607–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, J.H.; Poudel, R.P.; Tsagkrasoulis, D.; Caan, M.W.; Steves, C.; Spector, T.D.; Montana, G. Predicting brain age with deep learning from raw imaging data results in a reliable and heritable biomarker. NeuroImage 2017, 163, 115–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habeck, C.; Gazes, Y.; Razlighi, Q.; Stern, Y. Cortical thickness and its associations with age, total cognition and education across the adult lifespan. PLoS ONE 2020, 15, e0230298. [Google Scholar] [CrossRef] [Green Version]
- Cabeza, R.; Nyberg, L.; Park, D.C. Cognitive Neuroscience of Aging: Linking Cognitive and Cerebral Aging; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Morcom, A.M.; Henson, R.N. Increased prefrontal activity with aging reflects nonspecific neural responses rather than compensation. J. Neurosci. 2018, 38, 7303–7313. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.E.; Biessels, G.J.; De Guio, F.; De Leeuw, F.E.; Duchesne, S.; Düring, M.; Frayne, R.; Ikram, M.A.; Jouvent, E.; MacIntosh, B.J.; et al. Harmonizing brain magnetic resonance imaging methods for vascular contributions to neurodegeneration. Alzheimer Dementia Diagn. Assess. Dis. Monit. 2019, 11, 191–204. [Google Scholar] [CrossRef]
- Valverde, J.M.; Imani, V.; Abdollahzadeh, A.; De Feo, R.; Prakash, M.; Ciszek, R.; Tohka, J. Transfer learning in magnetic resonance brain imaging: A systematic review. J. Imaging 2021, 7, 66. [Google Scholar] [CrossRef]
- Chen, C.L.; Hsu, Y.C.; Yang, L.Y.; Tung, Y.H.; Luo, W.B.; Liu, C.M.; Hwang, T.J.; Hwu, H.G.; Tseng, W.Y.I. Generalization of diffusion magnetic resonance imaging–based brain age prediction model through transfer learning. NeuroImage 2020, 217, 116831. [Google Scholar] [CrossRef]
- Shafto, M.A.; Tyler, L.K.; Dixon, M.; Taylor, J.R.; Rowe, J.B.; Cusack, R.; Calder, A.J.; Marslen-Wilson, W.D.; Duncan, J.; Dalgleish, T.; et al. The Cambridge Centre for Ageing and Neuroscience (Cam-CAN) study protocol: A cross-sectional, lifespan, multidisciplinary examination of healthy cognitive ageing. BMC Neurol. 2014, 14, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Beheshti, I.; Ganaie, M.; Paliwal, V.; Rastogi, A.; Razzak, I.; Tanveer, M. Predicting brain age using machine learning algorithms: A comprehensive evaluation. IEEE J. Biomed. Health Inform. 2021, 26, 1432–1440. [Google Scholar] [CrossRef]
- Cole, J.H.; Leech, R.; Sharp, D.J.; Initiative, A.D.N. Prediction of brain age suggests accelerated atrophy after traumatic brain injury. Ann. Neurol. 2015, 77, 571–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Knol, M.J.; Tiulpin, A.; Dubost, F.; de Bruijne, M.; Vernooij, M.W.; Adams, H.H.; Ikram, M.A.; Niessen, W.J.; Roshchupkin, G.V. Gray matter age prediction as a biomarker for risk of dementia. Proc. Natl. Acad. Sci. USA 2019, 116, 21213–21218. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SAMPLE SIZE (MRI Scans) = 3514 | |
|---|---|
| Age | |
| Sex | F 519, M 280 |
| APOE | 7 |
| Education | None 150, Primary 232, |
| Secondary 203, University 214 |
| Volume (mm) | Sex p-val | APOE PR (>F) | |
|---|---|---|---|
| Brain2ICV | ** | > | |
| LH Th | ** | > | |
| RH Th | ** | > | |
| LH Pu | ** | > | |
| RH Pu | ** | > | |
| LH Am | ** | ** | |
| RH Am | ** | * | |
| LH Pa | ** | > | |
| RH Pa | ** | > | |
| LH Ca | ** | > | |
| RH Ca | ** | > | |
| LH Hp | ** | > | |
| RH Hp | ** | ** | |
| LH Ac | ** | > | |
| RH Ac | ** | > | |
| Avg.Thickness (mm) | Sex (p-val) | APOE Pr (>F) | |
| RH STempSuperior | ** | > | |
| LH STempSuperior | ** | > | |
| RH STempSuperior | ** | > | |
| LH STempInferior | ** | > | |
| RH STempInferior | ** | > | |
| LH SOccTempMedandLingual | ** | > | |
| RH SOccTempMedandLingual | ** | > | |
| LH SOccTempLat | ** | > | |
| RH SOccTempLat | ** | > | |
| RH GTempMid | > | > | |
| LH GTempInf | > | > | |
| RH GTempInf | > | > | |
| LH GTempSup | ** | > | |
| RH GTempSup | ** | > | |
| LH GTempSupPlanPolar | * | > | |
| RH GTempSupPlanPolar | ** | > | |
| LH GTempSupLateral | ** | > | |
| RH GTempSupLateral | > | > | |
| LH GTempSupTransv | ** | > | |
| RH GTempSupTransv | ** | > | |
| RH SIn | ** | > | |
| LH SFrontSup | ** | > | |
| RH SFrontSup | ** | > | |
| LH SFrontMid | ** | > | |
| RH SFrontMid | > | > | |
| LH SFrontInf | > | > | |
| RH SFrontInf | > | ** | |
| LH SFrontSup | ** | > | |
| RH SFrontSup | ** | > | |
| LH GFrontSupp | ** | > | |
| RH GFrontSupp | ** | > | |
| LH GFrontMid | ** | > | |
| RH GFrontMid | ** | > | |
| LH GFrontInfTriangul | ** | ||
| RH GFrontInfTriangul | ** | ||
| LH GFrontInfOrbital | ** | > | |
| RH GFrontInfOrbital | ** | > | |
| LH GFrontInfOpercular | > | > | |
| RH GFrontInfOpercular | * | > | |
| LH GCingPostV | * | > | |
| RH GCingPostV | > | > | |
| LH SCingMarginalis | > | > | |
| RH SCingMarginalis | ** | > | |
| LH SSubParietal | ** | > | |
| RH SSubParietal | ** | > | |
| LH SSubOrbital | > | > | |
| RH SSubOrbital | * | ** | |
| LH SPreCentralSuperior | ** | > | |
| RH SPreCentralSuperior | ** | > | |
| LH SPreCentralInferior | ** | > | |
| RH SPreCentralInferior | > | > | |
| LH SPostCentral | ** | > | |
| RH SPostCentral | ** | > | |
| RH SPeriCallosal | ** | > | |
| LH SParietoOcc | ** | > | |
| RH SParietoOcc | ** | > | |
| LH SOrbMedOlfact | ** | > | |
| RH SOrbMedOlfact | ** | > | |
| LH SOrbitalLat | ** | > | |
| RH SOrbitalLat | ** | * | |
| LH SOrbitalHShaped | ** | > | |
| RH SOrbitalHShaped | ** | > | |
| LH SOccMideandLunatus | ** | > | |
| RH SOccMideandLunatus | ** | > | |
| LH SIntraParietandPariettrans | ** | ||
| RH SIntraParietandPariettrans | ** | ||
| RH GParietalSup | ** | > | |
| LH GParietInfSupramar | ** | > | |
| RH GParietInfSupramar | ** | > | |
| LH GParietInfAngular | ** | > | |
| RH GParietInfAngular | ** | ** | |
| LH SCollatTransvPost | ** | > | |
| RH SCollTatransvPost | ** | > | |
| LH SCollTransvAnt | ** | > | |
| RH SCollTransvAnt | ** | > | |
| LH PoleOcc | > | > | |
| RH PoleOcc | ** | > | |
| LH GOccSup | ** | > | |
| RH GOccSup | ** | > | |
| LH GOccMid | ** | > | |
| RH GOccMid | ** | > | |
| LH GOccTempMedParahip | ** | > | |
| RH GOccTempMedParahip | ** | > | |
| LH GOccTempMedLingual | > | > | |
| RH GOccTempMedLingual | > | > | |
| LH GOccTempLatFusi | > | > | |
| RH GOccTempLatFusi | ** | > | |
| LH SInsSup | > | > | |
| RH SInsSup | ** | ** | |
| LH SInsInf | ** | > | |
| RH SInsInf | > | > | |
| LH SCircInsAnt | > | > | |
| RH SCircInsAnt | ** | > | |
| LH GInsularShort | ** | > | |
| RH GInsularShort | ** | > | |
| LH GCentInsula | ** | > | |
| RH GCentInsula | ** | > | |
| LH SCentral | ** | > | |
| RH SCentral | ** | > | |
| LH GPreCentral | ** | > | |
| RH GPreCentral | ** | > | |
| LH GPostCentral | ** | > | |
| RH GPostCentral | ** | * | |
| LH SCalcarine | > | > | |
| RH SCalcarine | ** | > | |
| LH GRectus | > | > | |
| RH GRectus | > | > | |
| LH GOrbital | ** | > | |
| RH GOrbital | ** | > | |
| LH GCuneus | > | > | |
| RH GCuneus | ** | > | |
| LH LatFisPost | ** | > | |
| RH LatFisPost | ** | * | |
| LH LatFisAntHoriz | > | > | |
| RH LatFisAntHoriz | ** | > |
| Test Performance Measure | ||||
|---|---|---|---|---|
| Model | MAE | MXE | MAPE | MEDAE |
| PLS | 2.570177 | 10.2293404 | 0.03348523 | 2.15809710 |
| XGBoost | 2.0301 | 8.7138485 | 0.0265 | 1.74578 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Ramírez, J.; Fernández-Blázquez, M.A.; González-Rosa, J.J. Prediction of Chronological Age in Healthy Elderly Subjects with Machine Learning from MRI Brain Segmentation and Cortical Parcellation. Brain Sci. 2022, 12, 579. https://doi.org/10.3390/brainsci12050579
Gómez-Ramírez J, Fernández-Blázquez MA, González-Rosa JJ. Prediction of Chronological Age in Healthy Elderly Subjects with Machine Learning from MRI Brain Segmentation and Cortical Parcellation. Brain Sciences. 2022; 12(5):579. https://doi.org/10.3390/brainsci12050579
Chicago/Turabian StyleGómez-Ramírez, Jaime, Miguel A. Fernández-Blázquez, and Javier J. González-Rosa. 2022. "Prediction of Chronological Age in Healthy Elderly Subjects with Machine Learning from MRI Brain Segmentation and Cortical Parcellation" Brain Sciences 12, no. 5: 579. https://doi.org/10.3390/brainsci12050579
APA StyleGómez-Ramírez, J., Fernández-Blázquez, M. A., & González-Rosa, J. J. (2022). Prediction of Chronological Age in Healthy Elderly Subjects with Machine Learning from MRI Brain Segmentation and Cortical Parcellation. Brain Sciences, 12(5), 579. https://doi.org/10.3390/brainsci12050579