
Chronic Low Dose Morphine Does Not Alter Two In Vitro BBB Models

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. hCMEC/D3 Media, hCMEC/D3 Cell Line Culture, and In Vitro Blood-Brain Barrier Generation
2.2. Primary Cell Culture
2.3. Human BBB Model
2.4. Morphine and Other Reagents
2.5. Morphine Exposure of Cells
2.6. Fluorescein Isothiocyanate—Dextran (FITC-D) Permeability Assay for hCMEC/D3
2.7. BBB Permeability for Primary Co-Culture
2.8. CCL2 (MCP-1) ELISA
2.9. Transmigration/Flow Cytometry
2.10. Statistics
3. Results
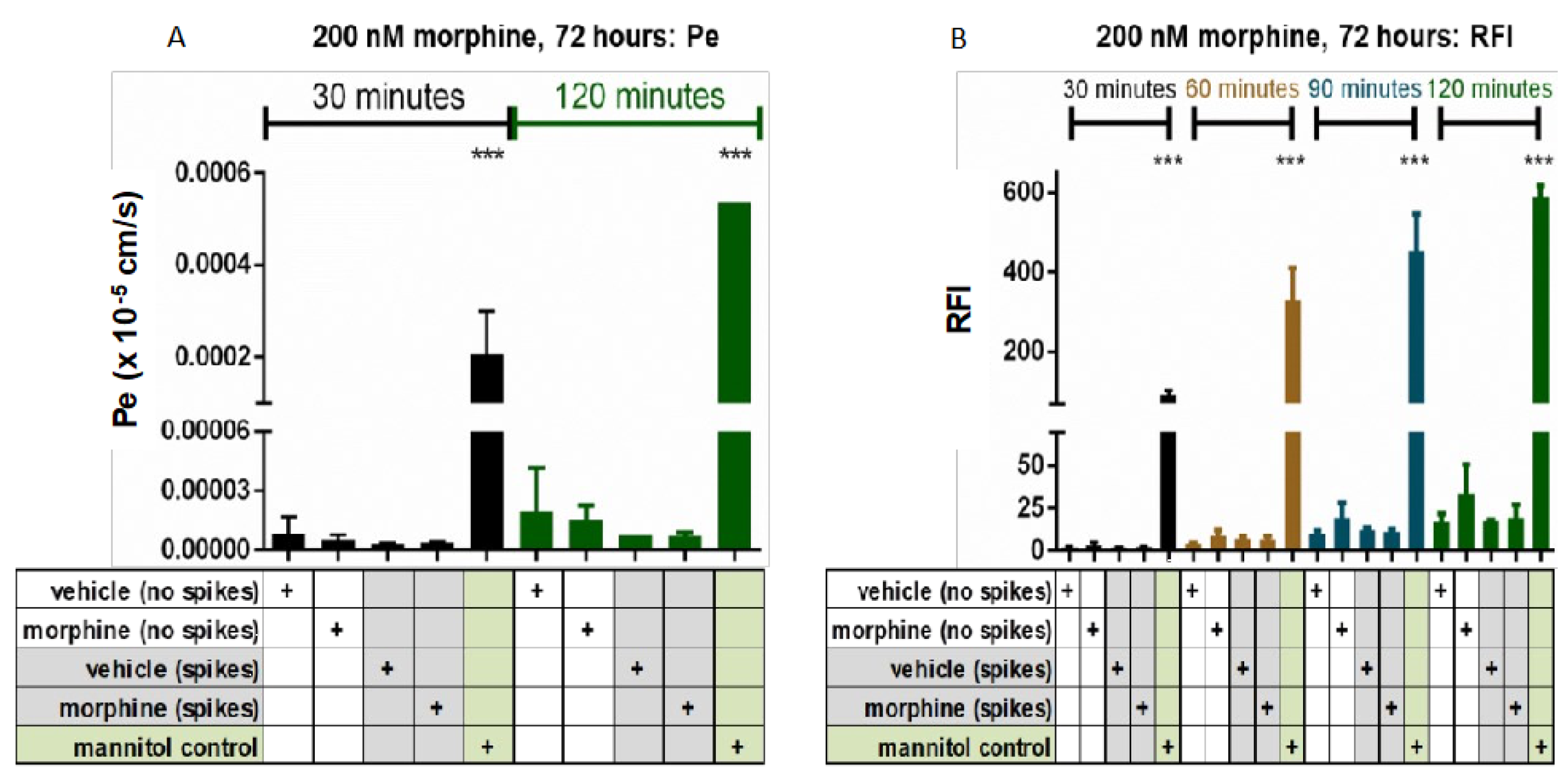
3.1. Morphine Exposure Does Not Alter the Basal Rate of Tracer Molecule Passage across hCMEC/D3 BBB Cells, Regardless of Single or Repeated Administration of Morphine
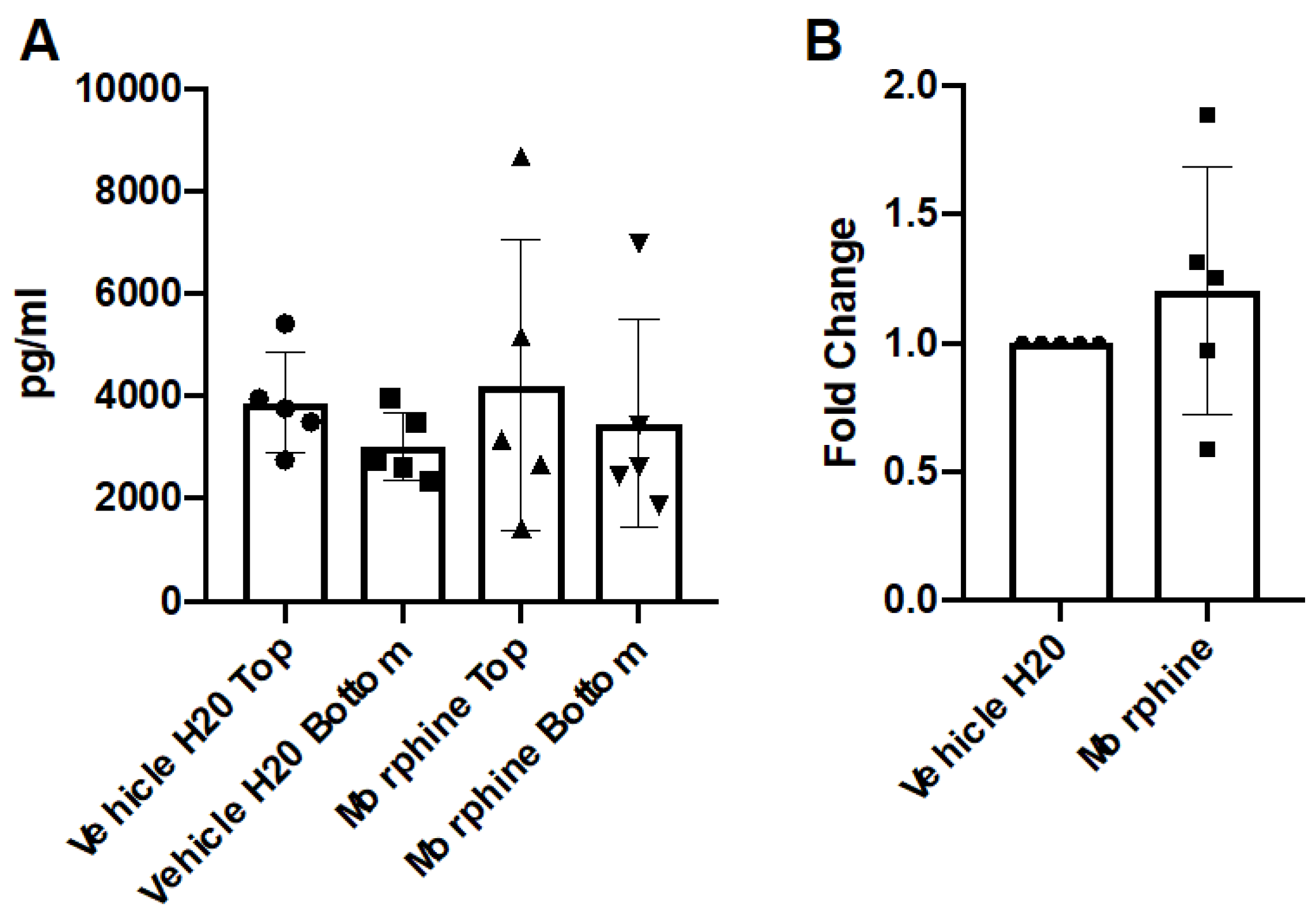
3.2. Repeated, Prolonged Morphine Exposure Does Not Induce Cytokine Gradients across a hCMEC/D3 BBB Model
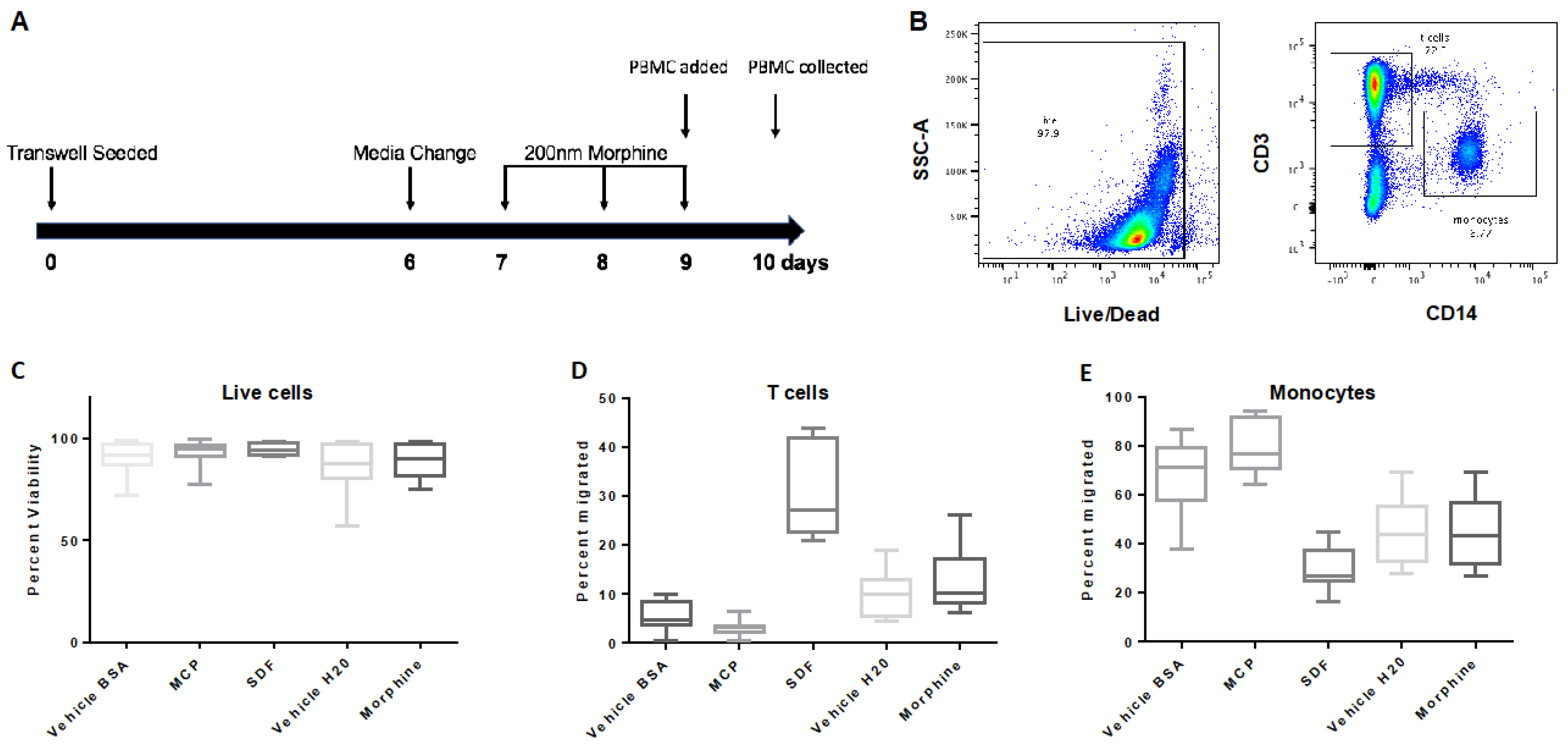
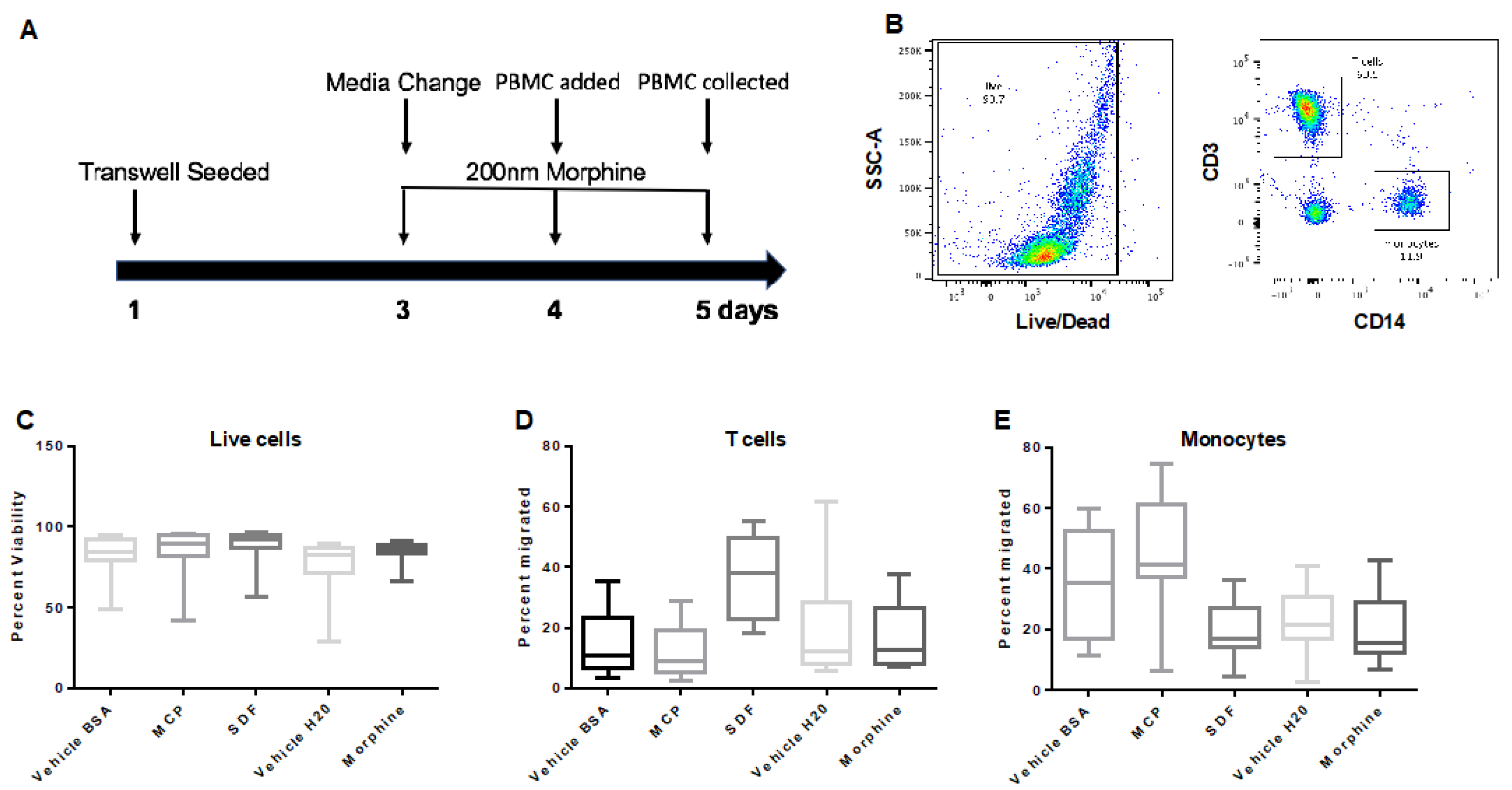
3.3. Repeated, Prolonged Morphine Exposure Does Not Alter PBMC Transmigration across a hCMEC/D3 BBB Model
3.4. Morphine Exposure Does Not Alter Evans Blue Permeability across a Primary Co-Culture BBB Model
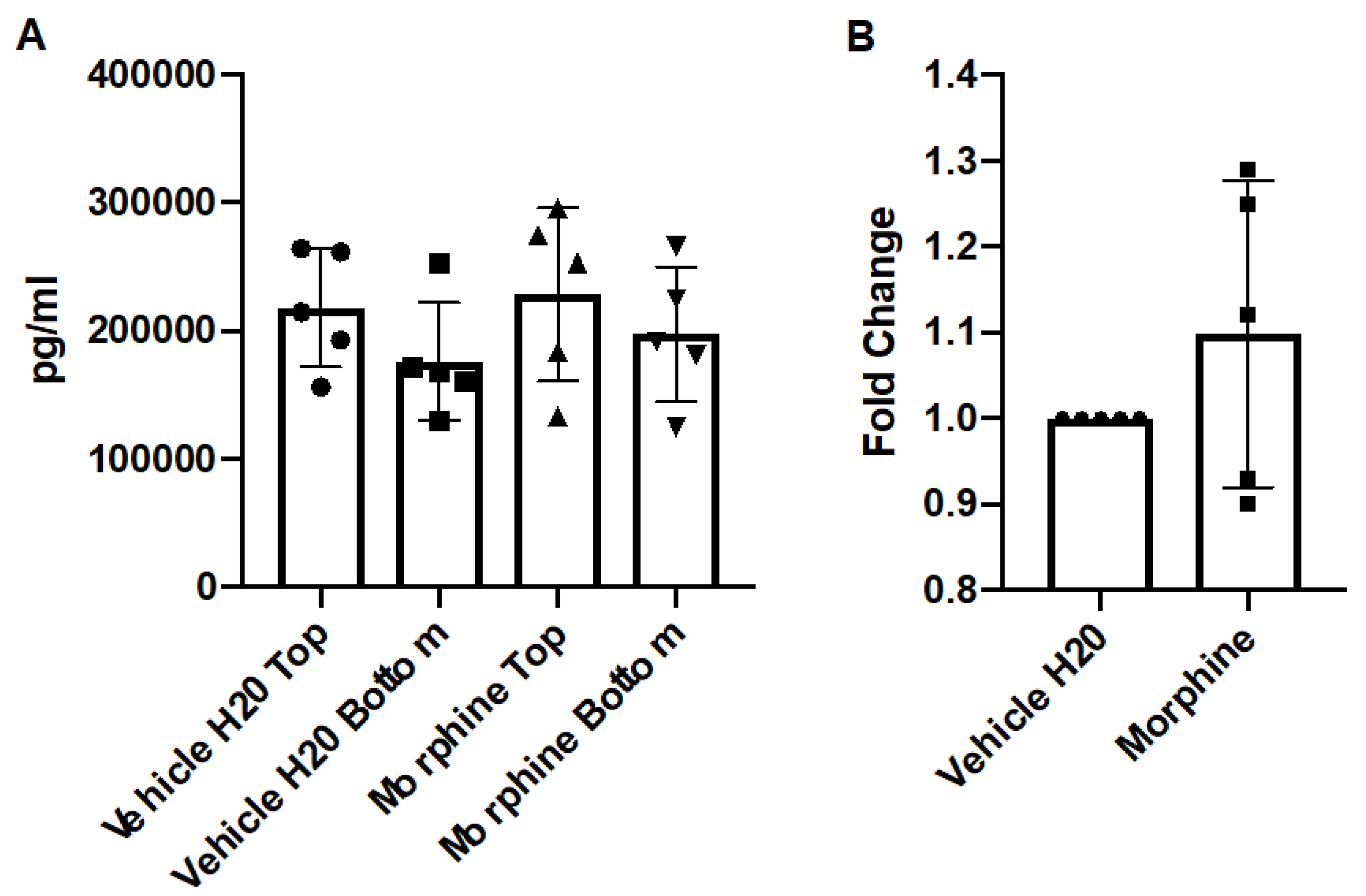
3.5. Repeated and Prolonged Morphine Exposure Does Not Induce Cytokine Gradients across a Primary Co-Culture BBB Model
3.6. Repeated, Prolonged Morphine Exposure Does Not Alter PBMC Transmigration across a Primary Human Co-Culture BBB Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, D.W.; Eugenin, E.A.; Calderon, T.M.; Berman, J.W. Monocyte maturation, HIV susceptibility, and transmigration across the blood brain barrier are critical in HIV neuropathogenesis. J. Leukoc. Biol. 2012, 91, 401–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachani, M.; Sacktor, N.; McArthur, J.C.; Nath, A.; Rumbaugh, J. Detection of anti-tat antibodies in CSF of individuals with HIV-associated neurocognitive disorders. J. Neurovirol. 2013, 19, 82–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, R.; Krishnan, A.; Meng, J.; Das, S.; Ma, J.; Banerjee, S.; Wang, J.; Charboneau, R.; Prakash, O.; Barke, R.A.; et al. Morphine modulation of toll-like receptors in microglial cells potentiates neuropathogenesis in a HIV-1 model of coinfection with pneumococcal pneumoniae. J. Neurosci. 2012, 32, 9917–9930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderon, T.M.; Williams, D.W.; Lopez, L.; Eugenin, E.A.; Cheney, L.; Gaskill, P.J.; Veenstra, M.; Anastos, K.; Morgello, S.; Berman, J.W. Dopamine Increases CD14(+)CD16(+) Monocyte Transmigration across the Blood Brain Barrier: Implications for Substance Abuse and HIV Neuropathogenesis. J. Neuroimmune Pharmacol. 2017, 12, 353–370. [Google Scholar] [CrossRef]
- Weiss, N.; Miller, F.; Cazaubon, S.; Couraud, P.O. The blood-brain barrier in brain homeostasis and neurological diseases. Biochim. Biophys. Acta 2009, 1788, 842–857. [Google Scholar] [CrossRef] [Green Version]
- De Silva, T.M.; Faraci, F.M. Microvascular Dysfunction and Cognitive Impairment. Cell Mol. Neurobiol. 2016, 36, 241–258. [Google Scholar] [CrossRef] [Green Version]
- Cotto, B.; Natarajaseenivasan, K.; Langford, D. Astrocyte activation and altered metabolism in normal aging, age-related CNS diseases, and HAND. J. Neurovirol. 2019, 25, 722–733. [Google Scholar] [CrossRef]
- Atluri, V.S.; Hidalgo, M.; Samikkannu, T.; Kurapati, K.R.; Jayant, R.D.; Sagar, V.; Nair, M.P. Effect of human immunodeficiency virus on blood-brain barrier integrity and function: An update. Front. Cell Neurosci. 2015, 9, 212. [Google Scholar] [CrossRef]
- Almutairi, M.M.; Gong, C.; Xu, Y.G.; Chang, Y.; Shi, H. Factors controlling permeability of the blood-brain barrier. Cell Mol. Life Sci. 2016, 73, 57–77. [Google Scholar] [CrossRef]
- Eugenin, E.A.; Clements, J.E.; Zink, M.C.; Berman, J.W. Human immunodeficiency virus infection of human astrocytes disrupts blood-brain barrier integrity by a gap junction-dependent mechanism. J. Neurosci. 2011, 31, 9456–9465. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.H.; Ismaiel, O.A.; Mylott, W.R., Jr.; Yuan, M.; McClay, J.L.; Paris, J.J.; Hauser, K.F.; McRae, M. Cell-type specific differences in antiretroviral penetration and the effects of HIV-1 Tat and morphine among primary human brain endothelial cells, astrocytes, pericytes, and microglia. Neurosci. Lett. 2019, 712, 134475. [Google Scholar] [CrossRef] [PubMed]
- Cassol, E.; Alfano, M.; Biswas, P.; Poli, G. Monocyte-derived macrophages and myeloid cell lines as targets of HIV-1 replication and persistence. J. Leukoc. Biol. 2006, 80, 1018–1030. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.D.; Aalinkeel, R.; Sykes, D.E.; Reynolds, J.L.; Bindukumar, B.; Fernandez, S.F.; Chawda, R.; Shanahan, T.C.; Schwartz, S.A. Tight junction regulation by morphine and HIV-1 tat modulates blood-brain barrier permeability. J. Clin. Immunol. 2008, 28, 528–541. [Google Scholar] [CrossRef] [PubMed]
- Merlin, J.S.; Long, D.; Becker, W.C.; Cachay, E.R.; Christopoulos, K.A.; Claborn, K.; Crane, H.M.; Edelman, E.J.; Harding, R.; Kertesz, S.G.; et al. Brief Report: The Association of Chronic Pain and Long-Term Opioid Therapy With HIV Treatment Outcomes. J. Acquir. Immune. Defic. Syndr. 2018, 79, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.J.; Fu, Y.Y.; Wei, Q.Q.; Zhang, Z.J. Neuroinflammation in HIV-Related Neuropathic Pain. Front. Pharmacol. 2021, 12, 653852. [Google Scholar] [CrossRef]
- Mikus, G.; Klimas, R. Contribution of oxycodone and its metabolites to the analgesic effect. Br. J. Anaesth. 2014, 112, 944–945. [Google Scholar] [CrossRef] [Green Version]
- Bicker, J.; Alves, G.; Fortuna, A.; Falcao, A. Blood-brain barrier models and their relevance for a successful development of CNS drug delivery systems: A review. Eur. J. Pharm. Biopharm. 2014, 87, 409–432. [Google Scholar] [CrossRef]
- Helms, H.C.; Abbott, N.J.; Burek, M.; Cecchelli, R.; Couraud, P.O.; Deli, M.A.; Forster, C.; Galla, H.J.; Romero, I.A.; Shusta, E.V.; et al. In vitro models of the blood-brain barrier: An overview of commonly used brain endothelial cell culture models and guidelines for their use. J. Cereb. Blood Flow. Metab. 2016, 36, 862–890. [Google Scholar] [CrossRef]
- Weksler, B.; Romero, I.A.; Couraud, P.O. The hCMEC/D3 cell line as a model of the human blood brain barrier. Fluids Barriers CNS 2013, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Weksler, B.B.; Subileau, E.A.; Perriere, N.; Charneau, P.; Holloway, K.; Leveque, M.; Tricoire-Leignel, H.; Nicotra, A.; Bourdoulous, S.; Turowski, P.; et al. Blood-brain barrier-specific properties of a human adult brain endothelial cell line. FASEB J. 2005, 19, 1872–1874. [Google Scholar] [CrossRef]
- Eigenmann, D.E.; Durig, C.; Jahne, E.A.; Smiesko, M.; Culot, M.; Gosselet, F.; Cecchelli, R.; Helms, H.C.C.; Brodin, B.; Wimmer, L.; et al. In vitro blood-brain barrier permeability predictions for GABAA receptor modulating piperine analogs. Eur. J. Pharm. Biopharm. 2016, 103, 118–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strazza, M.; Pirrone, V.; Wigdahl, B.; Dampier, W.; Lin, W.; Feng, R.; Maubert, M.E.; Weksler, B.; Romero, I.A.; Couraud, P.O.; et al. Prolonged Morphine Exposure Induces Increased Firm Adhesion in an in Vitro Model of the Blood-Brain Barrier. Int. J. Mol. Sci. 2016, 17, 916. [Google Scholar] [CrossRef] [PubMed]
- Berman, J.W.; Carvallo, L.; Buckner, C.M.; Luers, A.; Prevedel, L.; Bennett, M.V.; Eugenin, E.A. HIV-tat alters Connexin43 expression and trafficking in human astrocytes: Role in NeuroAIDS. J. Neuroinflammation 2016, 13, 54. [Google Scholar] [CrossRef] [Green Version]
- Eugenin, E.A.; Berman, J.W. Chemokine-dependent mechanisms of leukocyte trafficking across a model of the blood-brain barrier. Methods 2003, 29, 351–361. [Google Scholar] [CrossRef]
- Strazza, M.; Maubert, M.E.; Pirrone, V.; Wigdahl, B.; Nonnemacher, M.R. Co-culture model consisting of human brain microvascular endothelial and peripheral blood mononuclear cells. J. Neurosci. Methods 2016, 269, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Eugenin, E.A.; Osiecki, K.; Lopez, L.; Goldstein, H.; Calderon, T.M.; Berman, J.W. CCL2/monocyte chemoattractant protein-1 mediates enhanced transmigration of human immunodeficiency virus (HIV)-infected leukocytes across the blood-brain barrier: A potential mechanism of HIV-CNS invasion and NeuroAIDS. J. Neurosci. 2006, 26, 1098–1106. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Hu, X.T.; Napier, T.C.; Al-Harthi, L. Methamphetamine and HIV-1 Tat down regulate beta-catenin signaling: Implications for methampetamine abuse and HIV-1 co-morbidity. J. Neuroimmune Pharmacol. 2011, 6, 597–607. [Google Scholar] [CrossRef] [Green Version]
- Eugenin, E.A.; D’Aversa, T.G.; Lopez, L.; Calderon, T.M.; Berman, J.W. MCP-1 (CCL2) protects human neurons and astrocytes from NMDA or HIV-tat-induced apoptosis. J. Neurochem. 2003, 85, 1299–1311. [Google Scholar] [CrossRef]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
- Weiss Jonathan, D.S.; Berman, J. Astrocyte-Derived Monocyte-Chemoattractant protein-1 directs the transmigration of leukocytes across a model of the human blood-brain barrier. J. Immunol. 1998, 161, 6896–6903. [Google Scholar]
- Dhillon, N.K.; Williams, R.; Callen, S.; Zien, C.; Narayan, O.; Buch, S. Roles of MCP-1 in development of HIV-dementia. Front. Biosci. 2008, 13, 3913–3918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, S.; Sawaya, B.E.; Safak, M.; Batuman, O.; Khalili, K.; Amini, S. Regulation of MCP-1 gene transcription by Smads and HIV-1 Tat in human glial cells. Virology 2003, 309, 196–202. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Tsirka, S.E. Monocyte chemoattractant protein-1 and the blood-brain barrier. Cell Mol. Life Sci. 2014, 71, 683–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maubert, M.E.; Pirrone, V.; Rivera, N.T.; Wigdahl, B.; Nonnemacher, M.R. Interaction between Tat and Drugs of Abuse during HIV-1 Infection and Central Nervous System Disease. Front. Microbiol. 2015, 6, 1512. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marino, J.; Maubert, M.E.; Lawrence, J.M.; Wigdahl, B.; Nonnemacher, M.R. Chronic Low Dose Morphine Does Not Alter Two In Vitro BBB Models. Brain Sci. 2022, 12, 888. https://doi.org/10.3390/brainsci12070888
Marino J, Maubert ME, Lawrence JM, Wigdahl B, Nonnemacher MR. Chronic Low Dose Morphine Does Not Alter Two In Vitro BBB Models. Brain Sciences. 2022; 12(7):888. https://doi.org/10.3390/brainsci12070888
Chicago/Turabian StyleMarino, Jamie, Monique E. Maubert, Jill M. Lawrence, Brian Wigdahl, and Michael R. Nonnemacher. 2022. "Chronic Low Dose Morphine Does Not Alter Two In Vitro BBB Models" Brain Sciences 12, no. 7: 888. https://doi.org/10.3390/brainsci12070888
APA StyleMarino, J., Maubert, M. E., Lawrence, J. M., Wigdahl, B., & Nonnemacher, M. R. (2022). Chronic Low Dose Morphine Does Not Alter Two In Vitro BBB Models. Brain Sciences, 12(7), 888. https://doi.org/10.3390/brainsci12070888