
Cognitive Sequelae and Hippocampal Dysfunction in Chronic Kidney Disease following 5/6 Nephrectomy
 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Serum Biochemical Assays
2.3. Behavioral Tasks
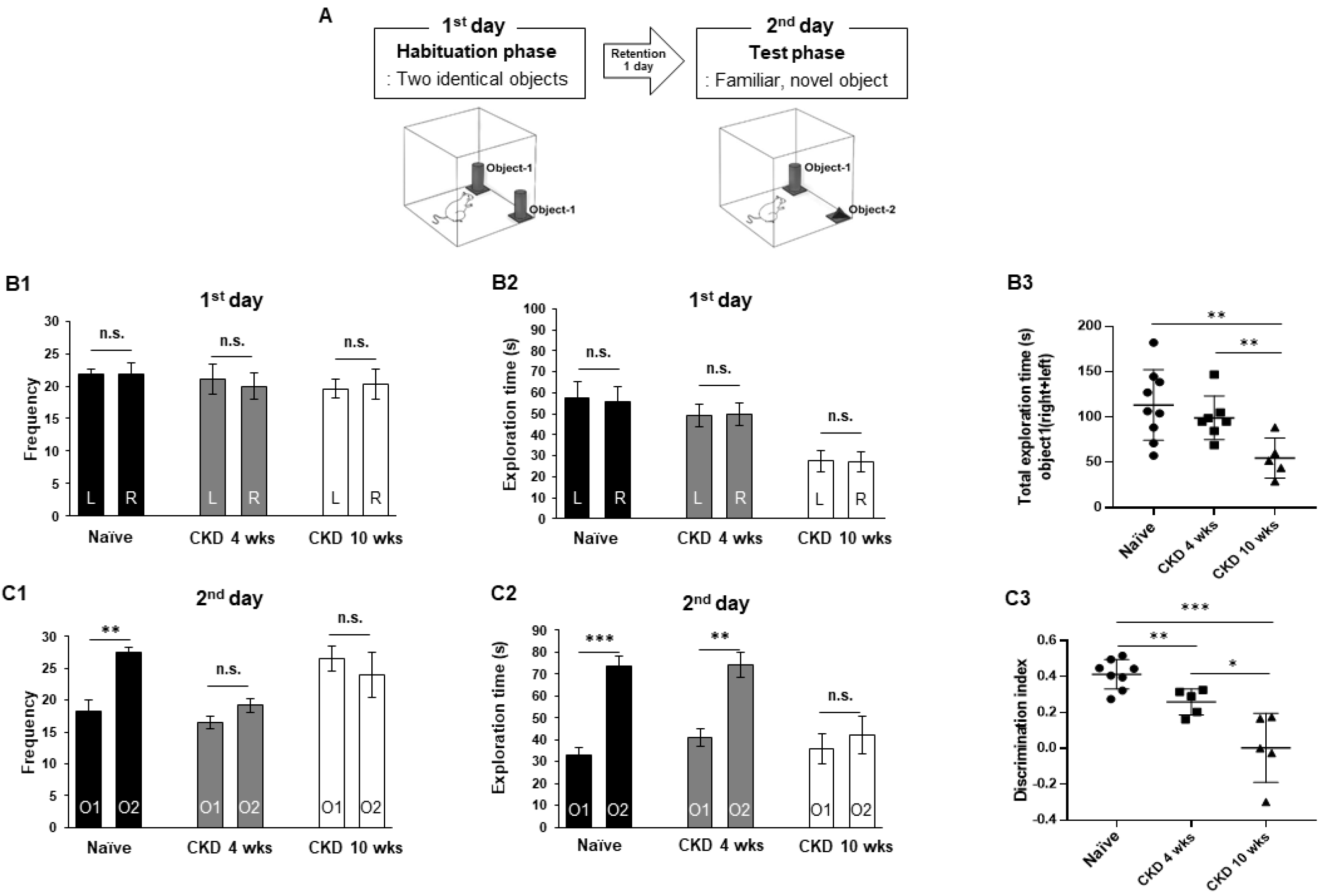
2.3.1. Novel Objective Recognition Test
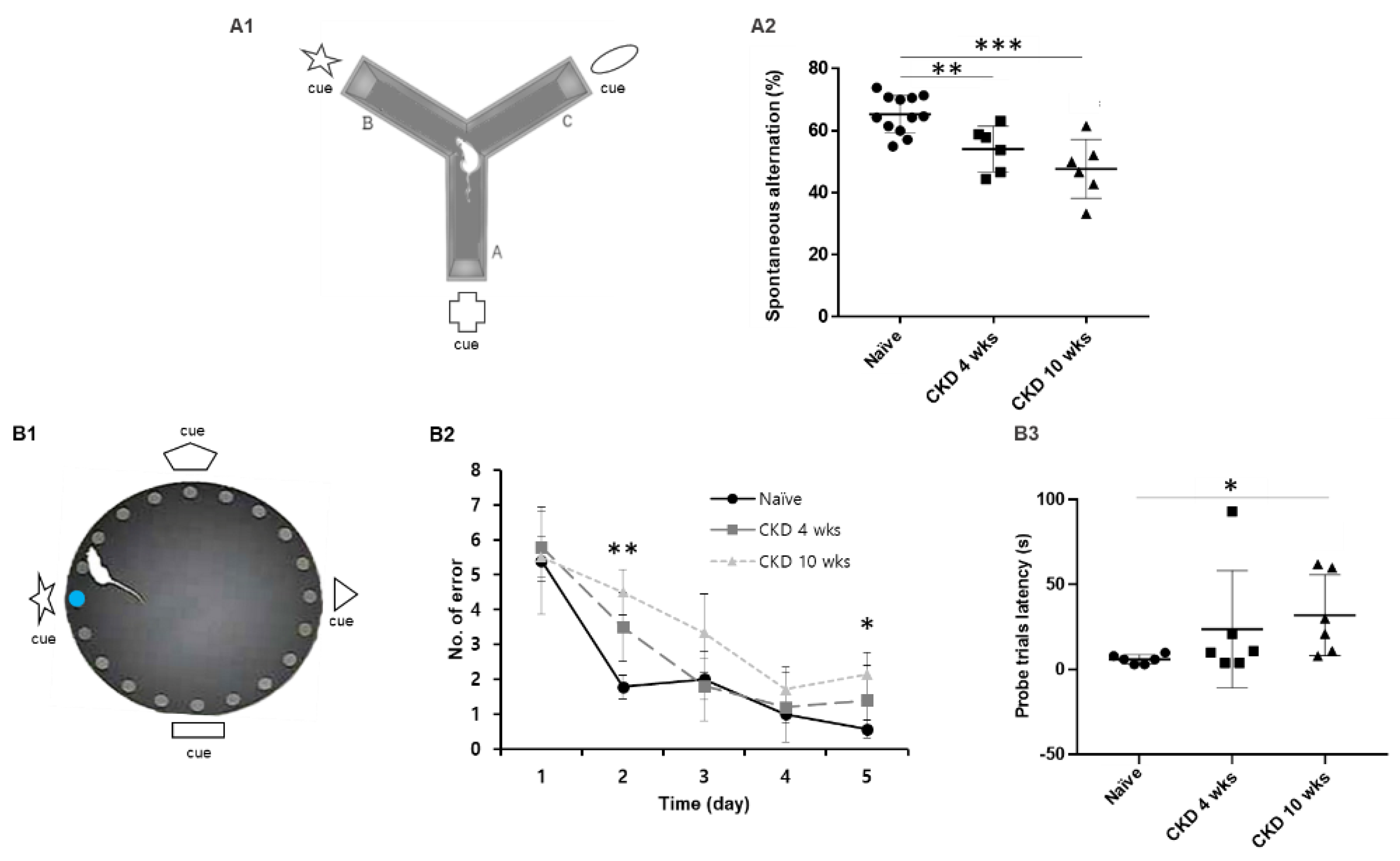
2.3.2. Y-Maze with Special Cue
2.3.3. Barnes Maze
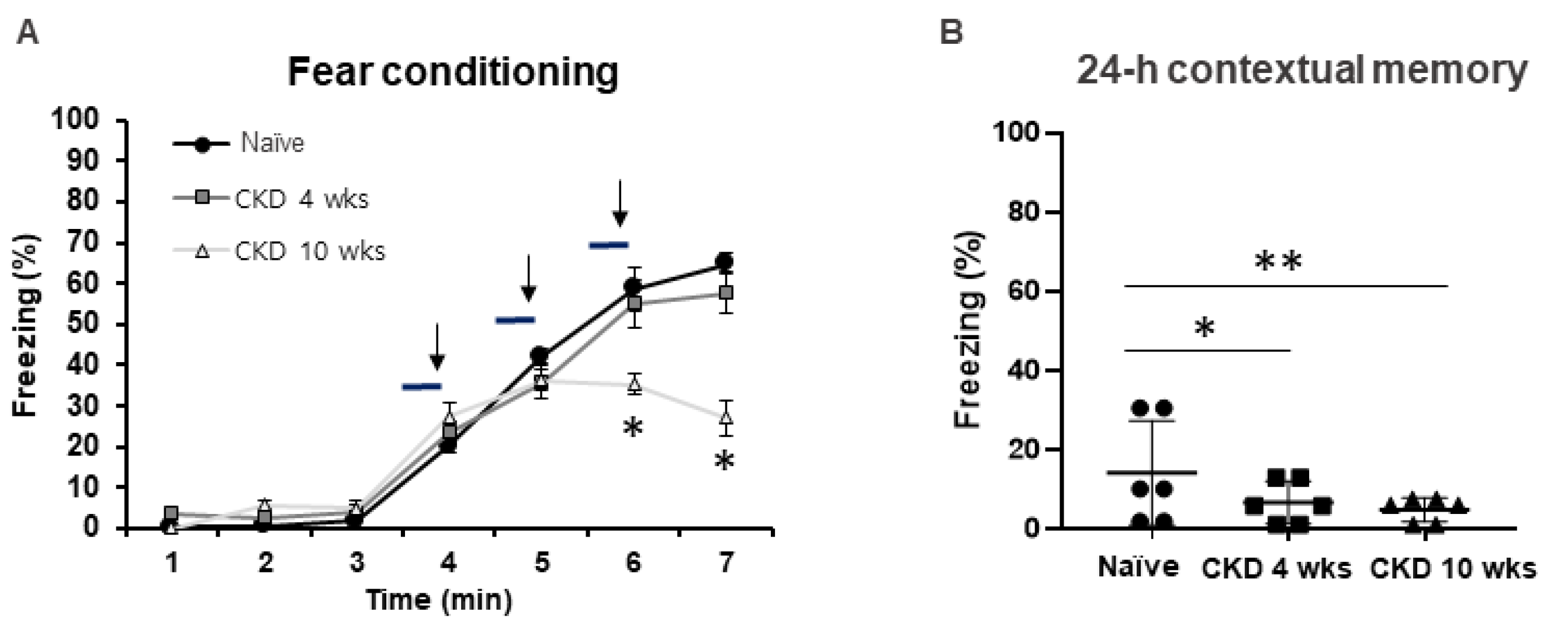
2.3.4. Classical Fear Conditioning Test
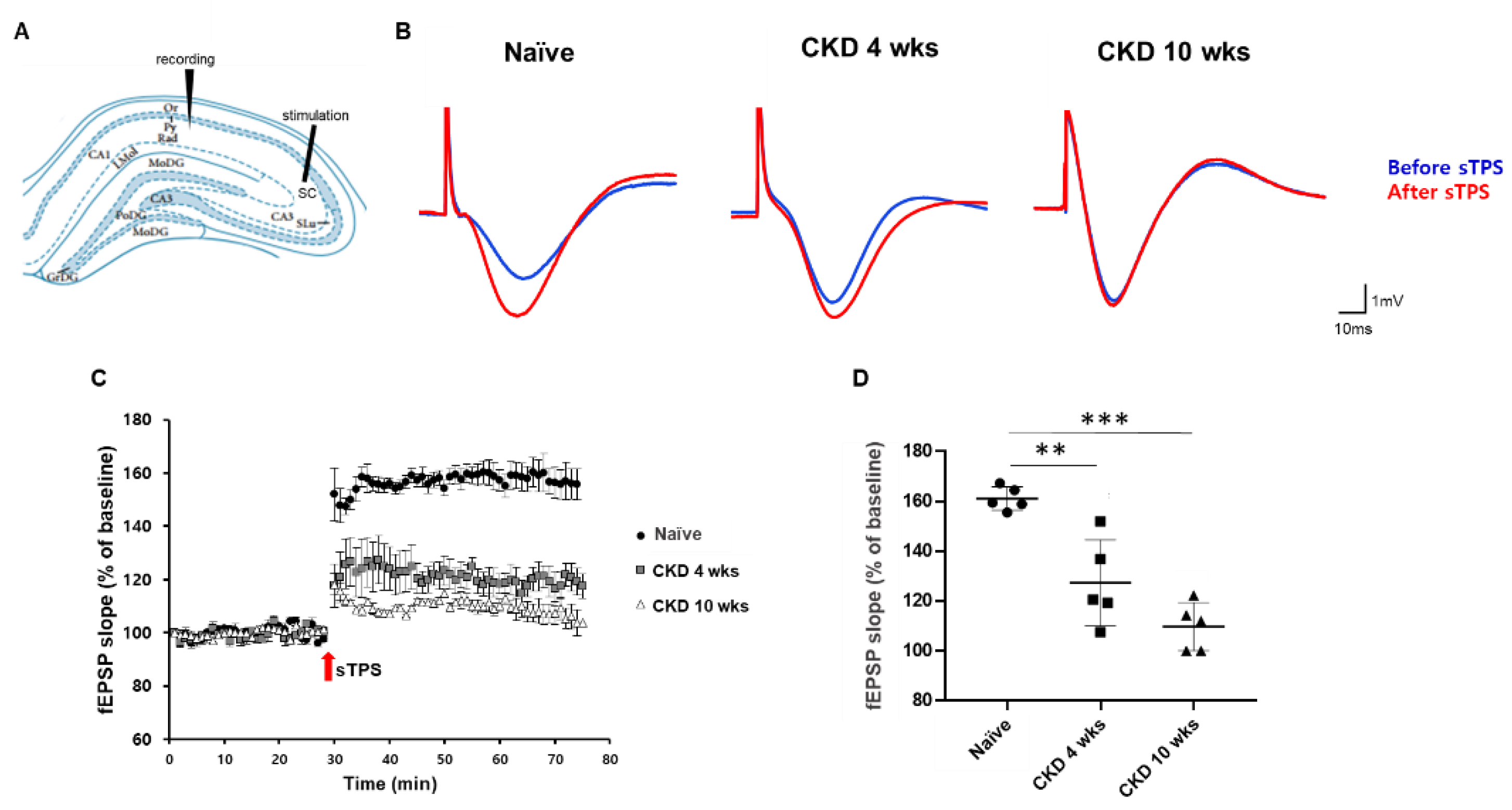
2.4. In Vivo fEPSP Recording
2.5. Tissue Processing and Immunohistochemistry (IHC)
2.6. Double Immunofluorescent Staining
2.7. Hippocampal Volume in Patients with Cognitive Impairment
2.8. Quantification of Data and Statistical Analysis
3. Results
3.1. Cognitive Behaviors Deficits in CKD Rat Model
3.2. Decreased fEPSP in the Hippocampal Neurons of CKD Rat Model
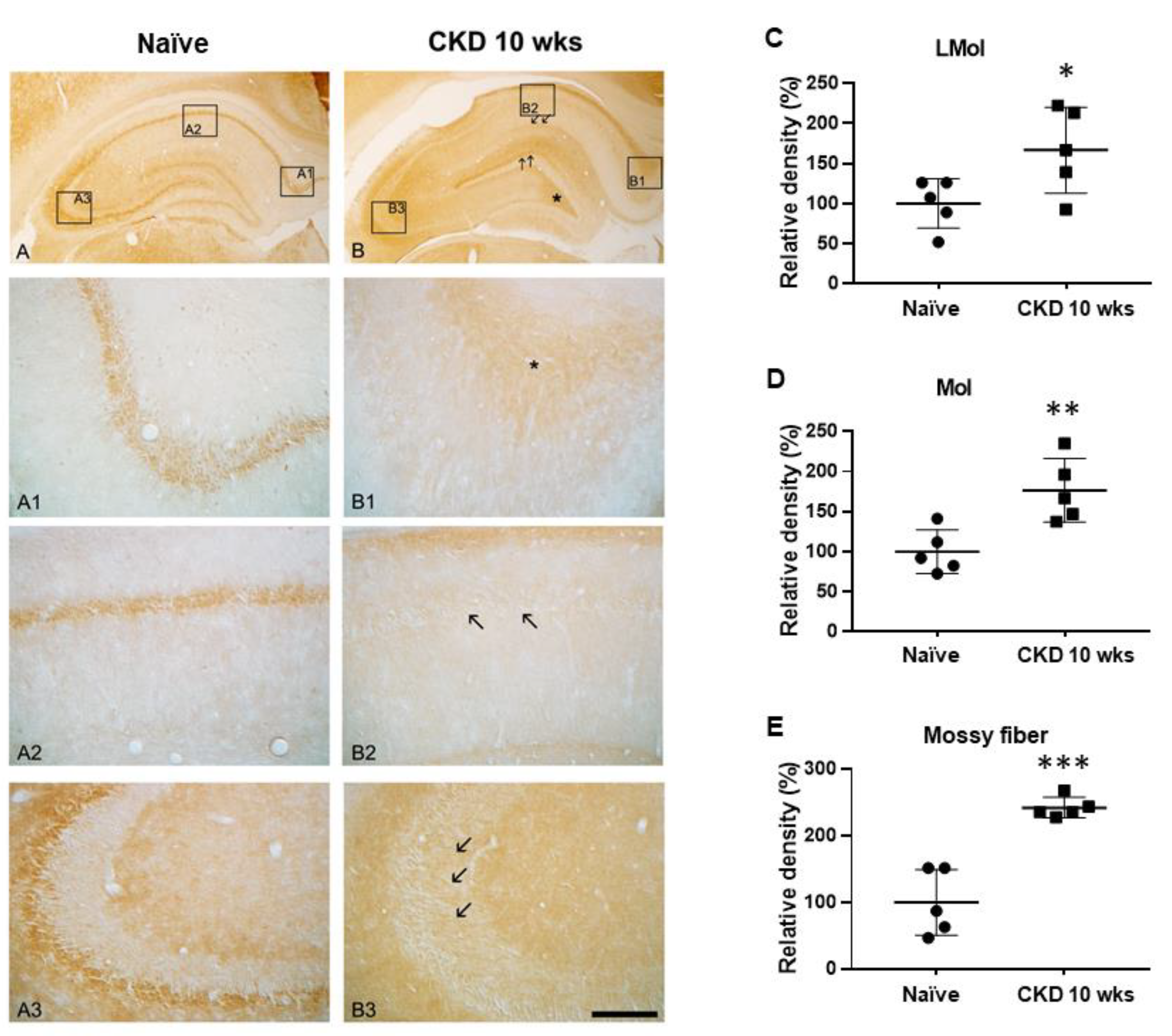
3.3. Immunoreactivities of Na+/H+ Exchanger-1 (NHE-1) in CKD Rat Model
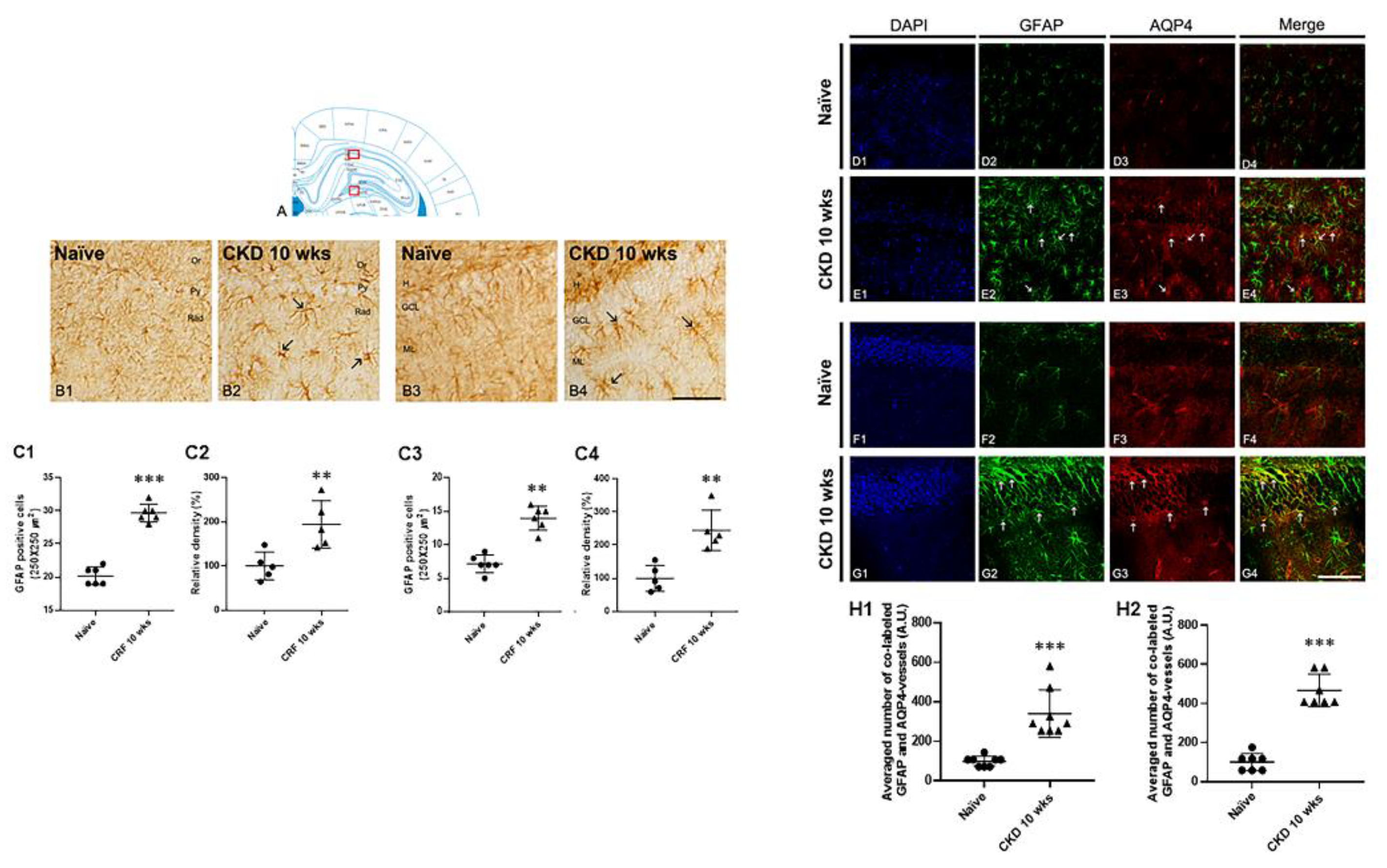
3.4. Astrogliosis in the Hippocampus of CKD Rat Model
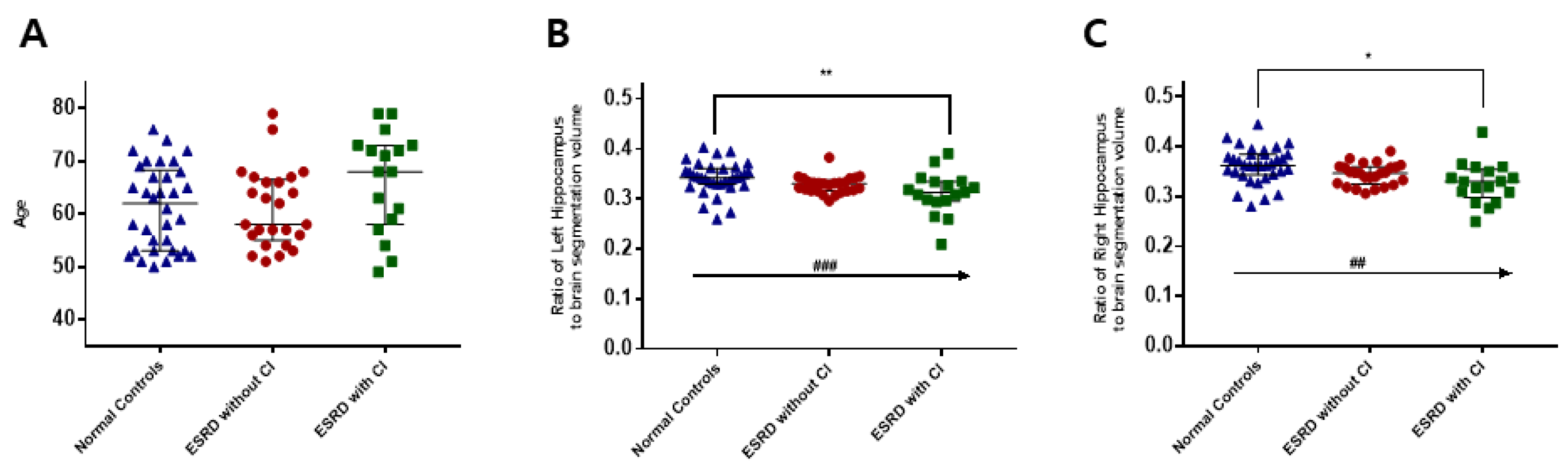
3.5. Hippocampal Volume in CKD Patients with Cognitive Impairment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lu, R.; Kiernan, M.C.; Murray, A.; Rosner, M.H.; Ronco, C. Kidney-brain crosstalk in the acute and chronic setting. Nat. Rev. Nephrol. 2015, 11, 707–719. [Google Scholar] [CrossRef] [PubMed]
- Madan, P.; Kalra, O.P.; Agarwal, S.; Tandon, O.P. Cognitive impairment in chronic kidney disease. Nephrol. Dial. Transplant. 2007, 22, 440–444. [Google Scholar] [CrossRef] [Green Version]
- Weiner, D.E.; Gaussoin, S.A.; Nord, J.; Auchus, A.P.; Chelune, G.J.; Chonchol, M.; Coker, L.; Haley, W.E.; Killeen, A.A.; Kimmel, P.L.; et al. Cognitive Function and Kidney Disease: Baseline Data From the Systolic Blood Pressure Intervention Trial (SPRINT). Am. J. Kidney Dis. 2017, 70, 357–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bugnicourt, J.M.; Godefroy, O.; Chillon, J.M.; Choukroun, G.; Massy, Z.A. Cognitive disorders and dementia in CKD: The neglected kidney-brain axis. J. Am. Soc. Nephrol. 2013, 24, 353–363. [Google Scholar] [CrossRef]
- Chaiben, V.B.O.; Silveira, T.B.D.; Guedes, M.H.; Fernandes, J.P.A.; Ferreira, J.H.F.; Beltrão, J.; Leal, G.F.; Erbano, L.H.O.; Bosch, N.L.; Pecoits Filho, R.; et al. Cognition and renal function: Findings from a Brazilian population. Braz. J. Nephrol. 2019, 41, 200–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, S.; Inoue, T.; Inagi, R. Neuroimmune interactions and kidney disease. Kidney Res. Clin. Pract. 2019, 38, 282–294. [Google Scholar] [CrossRef] [Green Version]
- Kurella Tamura, M.; Yaffe, K. Dementia and cognitive impairment in ESRD: Diagnostic and therapeutic strategies. Kidney Int. 2011, 79, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Cho, N.J.; Park, S.; Lee, E.Y.; Oh, S.W.; Oh, H.G.; Gil, H.W. Association of Intracranial Artery Calcification with Cognitive Impairment in Hemodialysis Patients. Med. Sci. Monit. 2019, 25, 5036–5043. [Google Scholar] [CrossRef] [PubMed]
- Mogi, M.; Horiuchi, M. Clinical Interaction between Brain and Kidney in Small Vessel Disease. Cardiol. Res. Pract. 2011, 2011, 306189. [Google Scholar] [CrossRef] [Green Version]
- Alster, P.; Dunalska, A.; Migda, B.; Madetko, N.; Królicki, L. The Rate of Decrease in Brain Perfusion in Progressive Supranuclear Palsy and Corticobasal Syndrome May Be Impacted by Glycemic Variability-A Pilot Study. Front. Neurol. 2021, 12, 767480. [Google Scholar] [CrossRef]
- Sexton, C.E.; Mackay, C.E.; Lonie, J.A.; Bastin, M.E.; Terrière, E.; O’Carroll, R.E.; Ebmeier, K.P. MRI correlates of episodic memory in Alzheimer’s disease, mild cognitive impairment, and healthy aging. Psychiatry Res. 2010, 184, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Josephs, K.A.; Murray, M.E.; Tosakulwong, N.; Whitwell, J.L.; Knopman, D.S.; Machulda, M.M.; Weigand, S.D.; Boeve, B.F.; Kantarci, K.; Petrucelli, L.; et al. Tau aggregation influences cognition and hippocampal atrophy in the absence of beta-amyloid: A clinico-imaging-pathological study of primary age-related tauopathy (PART). Acta Neuropathol. 2017, 133, 705–715. [Google Scholar] [CrossRef]
- Dawe, R.J.; Yu, L.; Arfanakis, K.; Schneider, J.A.; Bennett, D.A.; Boyle, P.A. Late-life cognitive decline is associated with hippocampal volume, above and beyond its associations with traditional neuropathologic indices. Alzheimers Dement 2020, 16, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Devanand, D.P.; Pradhaban, G.; Liu, X.; Khandji, A.; De Santi, S.; Segal, S.; Rusinek, H.; Pelton, G.H.; Honig, L.S.; Mayeux, R.; et al. Hippocampal and entorhinal atrophy in mild cognitive impairment: Prediction of Alzheimer disease. Neurology 2007, 68, 828–836. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, A.; Topiwala, M.A.; Gordon, J.A. Synchronized activity between the ventral hippocampus and the medial prefrontal cortex during anxiety. Neuron 2010, 65, 257–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.W.; Ha, G.Y.; Jung, Y.W. Chronic renal failure induces cell death in rat hippocampal CA1 via upregulation of alphaCaMKII/NR2A synaptic complex and phosphorylated GluR1-containing AMPA receptor cascades. Kidney Res. Clin. Pract. 2014, 33, 132–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malek, M. Brain consequences of acute kidney injury: Focusing on the hippocampus. Kidney Res. Clin. Pract. 2018, 37, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.H.; Kim, S.W.; Park, D.K.; Song, H.Y.; Kim, D.S.; Gil, H.W. Altered Emotional Phenotypes in Chronic Kidney Disease Following 5/6 Nephrectomy. Brain Sci. 2021, 11, 882. [Google Scholar] [CrossRef]
- Leger, M.; Quiedeville, A.; Bouet, V.; Haelewyn, B.; Boulouard, M.; Schumann-Bard, P.; Freret, T. Object recognition test in mice. Nat. Protoc. 2013, 8, 2531–2537. [Google Scholar] [CrossRef]
- Sivakumaran, M.H.; Mackenzie, A.K.; Callan, I.R.; Ainge, J.A.; O’Connor, A.R. The Discrimination Ratio derived from Novel Object Recognition tasks as a Measure of Recognition Memory Sensitivity, not Bias. Sci. Rep. 2018, 8, 11579. [Google Scholar] [CrossRef]
- Jiang, B.; Song, L.; Huang, C.; Zhang, W. P7C3 Attenuates the Scopolamine-Induced Memory Impairments in C57BL/6J Mice. Neurochem. Res. 2016, 41, 1010–1019. [Google Scholar] [CrossRef]
- Attar, A.; Liu, T.; Chan, W.T.; Hayes, J.; Nejad, M.; Lei, K.; Bitan, G. A shortened Barnes maze protocol reveals memory deficits at 4-months of age in the triple-transgenic mouse model of Alzheimer’s disease. PLoS ONE 2013, 8, e80355. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Guo, M.; Garza, J.; Rendon, S.; Sun, X.L.; Zhang, W.; Lu, X.Y. Cognitive and neural correlates of depression-like behaviour in socially defeated mice: An animal model of depression with cognitive dysfunction. Int. J. Neuropsychopharmacol. 2011, 14, 303–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, B.A.; Cho, T.; Lee, D.Z.; Lee, J.J.; Lee, B.; Kim, S.W.; Shin, H.S.; Kang, M.G. LARGE, an intellectual disability-associated protein, regulates AMPA-type glutamate receptor trafficking and memory. Proc. Natl. Acad. Sci. USA 2018, 115, 7111–7116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minamiura, Y.; Hirayama, K.; Murata, R.; Matsuura, S. Effect of hyperthermia on hippocampal synaptic transmission and CA3 kindling in developing rats. Brain Res. 1996, 732, 209–214. [Google Scholar] [CrossRef]
- Wong, T.P.; Howland, J.G.; Robillard, J.M.; Ge, Y.; Yu, W.; Titterness, A.K.; Brebner, K.; Liu, L.; Weinberg, J.; Christie, B.R.; et al. Hippocampal long-term depression mediates acute stress-induced spatial memory retrieval impairment. Proc. Natl. Acad. Sci. USA 2007, 104, 11471–11476. [Google Scholar] [CrossRef] [Green Version]
- Ge, Y.; Dong, Z.; Bagot, R.C.; Howland, J.G.; Phillips, A.G.; Wong, T.P.; Wang, Y.T. Hippocampal long-term depression is required for the consolidation of spatial memory. Proc. Natl. Acad. Sci. USA 2010, 107, 16697–16702. [Google Scholar] [CrossRef] [Green Version]
- Cho, T.; Ryu, J.K.; Taghibiglou, C.; Ge, Y.; Chan, A.W.; Liu, L.; Lu, J.; McLarnon, J.G.; Wang, Y.T. Long-term potentiation promotes proliferation/survival and neuronal differentiation of neural stem/progenitor cells. PLoS ONE 2013, 8, e76860. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.H.; Lee, K.; Sin, D.S.; Park, K.H.; Park, D.K.; Kim, D.S. Altered functional efficacy of hippocampal interneuron during epileptogenesis following febrile seizures. Brain Res. Bull. 2017, 131, 25–38. [Google Scholar] [CrossRef]
- Cho, Y.; Seong, J.K.; Jeong, Y.; Shin, S.Y. Individual subject classification for Alzheimer’s disease based on incremental learning using a spatial frequency representation of cortical thickness data. Neuroimage 2012, 59, 2217–2230. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.S.; Kim, C.; Shin, J.H.; Cho, H.; Shin, D.S.; Kim, N. Machine Learning-based Individual Assessment of Cortical Atrophy Pattern in Alzheimer’s Disease Spectrum: Development of the Classifier and Longitudinal Evaluation. Sci. Rep. 2018, 8, 4161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischl, B. FreeSurfer. Neuroimage 2012, 62, 774–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, C.; Ferreira, D.; Fink, K.; Westman, E.; Granberg, T. Repeatability and reproducibility of FreeSurfer, FSL-SIENAX and SPM brain volumetric measurements and the effect of lesion filling in multiple sclerosis. Eur Radiol. 2019, 29, 1355–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicoll, R.A. A Brief History of Long-Term Potentiation. Neuron 2017, 93, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Hölscher, C. Synaptic plasticity and learning and memory: LTP and beyond. J. Neurosci. Res. 1999, 58, 62–75. [Google Scholar] [CrossRef]
- Fujisaki, K.; Tsuruya, K.; Yamato, M.; Toyonaga, J.; Noguchi, H.; Nakano, T.; Taniguchi, M.; Tokumoto, M.; Hirakata, H.; Kitazono, T. Cerebral oxidative stress induces spatial working memory dysfunction in uremic mice: Neuroprotective effect of tempol. Nephrol. Dial. Transplant. 2014, 29, 529–538. [Google Scholar] [CrossRef] [Green Version]
- Degaspari, S.; Tzanno-Martins, C.B.; Fujihara, C.K.; Zatz, R.; Branco-Martins, J.P.; Viel, T.A.; Buck Hde, S.; Orellana, A.M.; Böhmer, A.E.; Lima Lde, S.; et al. Altered KLOTHO and NF-κB-TNF-α Signaling Are Correlated with Nephrectomy-Induced Cognitive Impairment in Rats. PLoS ONE 2015, 10, e0125271. [Google Scholar] [CrossRef]
- Tothova, L.; Babickova, J.; Borbelyova, V.; Filova, B.; Sebekova, K.; Hodosy, J. Chronic renal insufficiency does not induce behavioral and cognitive alteration in rats. Physiol. Behav. 2015, 138, 133–140. [Google Scholar] [CrossRef]
- Renczés, E.; Marônek, M.; Gaál Kovalčíková, A.; Vavrincová-Yaghi, D.; Tóthová, L.; Hodosy, J. Behavioral Changes During Development of Chronic Kidney Disease in Rats. Front. Med. 2019, 6, 311. [Google Scholar] [CrossRef] [Green Version]
- Antunes, M.; Biala, G. The novel object recognition memory: Neurobiology, test procedure, and its modifications. Cogn. Process. 2012, 13, 93–110. [Google Scholar] [CrossRef] [Green Version]
- Abu Ahmad, N.; Armaly, Z.; Berman, S.; Jabour, A.; Aga-Mizrachi, S.; Mosenego-Ornan, E.; Avital, A. l-Carnitine improves cognitive and renal functions in a rat model of chronic kidney disease. Physiol. Behav. 2016, 164, 182–188. [Google Scholar] [CrossRef]
- Teng, E.; Squire, L.R. Memory for places learned long ago is intact after hippocampal damage. Nature 1999, 400, 675–677. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.J.; Wiltgen, B.J.; Fanselow, M.S. The place of the hippocampus in fear conditioning. Eur. J. Pharmacol. 2003, 463, 217–223. [Google Scholar] [CrossRef]
- Franco, Á.O.; Starosta, R.T.; Roriz-Cruz, M. The specific impact of uremic toxins upon cognitive domains: A review. Braz. J. Nephrol. 2019, 41, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Palin, K.; Bluthé, R.M.; Verrier, D.; Tridon, V.; Dantzer, R.; Lestage, J. Interleukin-1beta mediates the memory impairment associated with a delayed type hypersensitivity response to bacillus Calmette-Guérin in the rat hippocampus. Brain Behav. Immun. 2004, 18, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Campbell, K.L.; Johnson, D.W.; Stanton, T.; Vesey, D.A.; Coombes, J.S.; Weston, K.S.; Hawley, C.M.; McWhinney, B.C.; Ungerer, J.P.; et al. Protein-bound uremic toxins, inflammation and oxidative stress: A cross-sectional study in stage 3–4 chronic kidney disease. Arch. Med. Res. 2014, 45, 309–317. [Google Scholar] [CrossRef]
- Russo, A.S.; Parsons, R.G. Behavioral Expression of Contextual Fear in Male and Female Rats. Front. Behav. Neurosci. 2021, 15, 671017. [Google Scholar] [CrossRef]
- Lynch, M.A. Long-term potentiation and memory. Physiol. Rev. 2004, 84, 87–136. [Google Scholar] [CrossRef]
- Hussain, R.J.; Carpenter, D.O. Development of synaptic responses and plasticity at the SC-CA1 and MF-CA3 synapses in rat hippocampus. Cell. Mol. Neurobiol. 2001, 21, 357–368. [Google Scholar] [CrossRef]
- Manabe, T.; Noda, Y.; Mamiya, T.; Katagiri, H.; Houtani, T.; Nishi, M.; Noda, T.; Takahashi, T.; Sugimoto, T.; Nabeshima, T.; et al. Facilitation of long-term potentiation and memory in mice lacking nociceptin receptors. Nature 1998, 394, 577–581. [Google Scholar] [CrossRef]
- Tang, Y.P.; Shimizu, E.; Dube, G.R.; Rampon, C.; Kerchner, G.A.; Zhuo, M.; Liu, G.; Tsien, J.Z. Genetic enhancement of learning and memory in mice. Nature 1999, 401, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Yamada, N.; Tsuzuki, K.; Horikawa, H.P.; Tanaka, K.; Ozawa, S. Long-term potentiation in the hippocampal CA1 area and dentate gyrus plays different roles in spatial learning. Eur. J. Neurosci. 2003, 17, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Dere, E.; De Souza-Silva, M.A.; Topic, B.; Spieler, R.E.; Haas, H.L.; Huston, J.P. Histidine-decarboxylase knockout mice show deficient nonreinforced episodic object memory, improved negatively reinforced water-maze performance, and increased neo- and ventro-striatal dopamine turnover. Learn. Mem. 2003, 10, 510–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, I.Y.; Lyons, W.E.; Mamounas, L.A.; Thompson, R.F. Brain-derived neurotrophic factor plays a critical role in contextual fear conditioning. J. Neurosci. 2004, 24, 7958–7963. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, S.F.; Ohtsu, H.; Benice, T.S.; Rizk-Jackson, A.; Raber, J. Age-dependent measures of anxiety and cognition in male histidine decarboxylase knockout (Hdc−/−) mice. Brain Res. 2006, 1071, 113–123. [Google Scholar] [CrossRef]
- Ma, E.; Haddad, G.G. Expression and localization of Na+/H+ exchangers in rat central nervous system. Neuroscience 1997, 79, 591–603. [Google Scholar] [CrossRef]
- Kang, T.C.; An, S.J.; Park, S.K.; Hwang, I.K.; Suh, J.G.; Oh, Y.S.; Bae, J.C.; Won, M.H. Alterations in Na+/H+ exchanger and Na+/HCO3− cotransporter immunoreactivities within the gerbil hippocampus following seizure. Mol. Brain Res. 2002, 109, 226–232. [Google Scholar] [CrossRef]
- Kang, T.C.; An, S.J.; Park, S.K.; Hwang, I.K.; Bae, J.C.; Won, M.H. The evidence for GABAB receptor-mediated regulation of acid-base balance: Involvement of Na+/H+ exchanger and Na+/HCO3− cotransporter. Mol. Brain Res. 2003, 114, 86–90. [Google Scholar] [CrossRef]
- Luo, J.; Sun, D. Physiology and pathophysiology of Na(+)/H(+) exchange isoform 1 in the central nervous system. Curr. Neurovasc. Res. 2007, 4, 205–215. [Google Scholar] [CrossRef]
- Song, H.; Yuan, S.; Zhang, Z.; Zhang, J.; Zhang, P.; Cao, J.; Li, H.; Li, X.; Shen, H.; Wang, Z.; et al. Sodium/Hydrogen Exchanger 1 Participates in Early Brain Injury after Subarachnoid Hemorrhage both in vivo and in vitro via Promoting Neuronal Apoptosis. Cell Transplant. 2019, 28, 985–1001. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Carney, K.E.; Falgoust, L.; Pan, J.W.; Sun, D.; Zhang, Z. Emerging roles of Na+/H+ exchangers in epilepsy and developmental brain disorders. Prog. Neurobiol. 2016, 138–140, 19–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haworth, R.S.; McCann, C.; Snabaitis, A.K.; Roberts, N.A.; Avkiran, M. Stimulation of the plasma membrane Na+/H+ exchanger NHE1 by sustained intracellular acidosis. Evidence for a novel mechanism mediated by the ERK pathway. J. Biol. Chem. 2003, 278, 31676–31684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coccaro, E.; Karki, P.; Cojocaru, C.; Fliegel, L. Phenylephrine and sustained acidosis activate the neonatal rat cardiomyocyte Na+/H+ exchanger through phosphorylation of amino acids Ser770 and Ser771. Am. J. Physiol. Heart Circ. 2009, 297, H846–H858. [Google Scholar] [CrossRef] [PubMed]
- Odunewu, A.; Fliegel, L. Acidosis-mediated regulation of the NHE1 isoform of the Na+/H+ exchanger in renal cells. Am. J. Physiol. Ren. Physiol. 2013, 305, F370–F381. [Google Scholar] [CrossRef]
- Luo, J.; Chen, H.; Kintner, D.B.; Shull, G.E.; Sun, D. Decreased neuronal death in Na+/H+ exchanger isoform 1-null mice after in vitro and in vivo ischemia. J. Neurosci. 2005, 25, 11256–11268. [Google Scholar] [CrossRef] [Green Version]
- Cengiz, P.; Kleman, N.; Uluc, K.; Kendigelen, P.; Hagemann, T.; Akture, E.; Messing, A.; Ferrazzano, P.; Sun, D. Inhibition of Na+/H+ exchanger isoform 1 is neuroprotective in neonatal hypoxic ischemic brain injury. Antioxid. Redox Signal. 2011, 14, 1803–1813. [Google Scholar] [CrossRef] [Green Version]
- Verma, V.; Bali, A.; Singh, N.; Jaggi, A.S. Implications of sodium hydrogen exchangers in various brain diseases. J. Basic Clin. Physiol. Pharmacol. 2015, 26, 417–426. [Google Scholar] [CrossRef]
- Rönicke, R.; Schröder, U.H.; Böhm, K.; Reymann, K.G. The Na+/H+ exchanger modulates long-term potentiation in rat hippocampal slices. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2009, 379, 233–239. [Google Scholar] [CrossRef]
- Kim, S.; Lee, J.; Heo, N.J.; Lee, J.W.; Han, J.S. Alkali therapy attenuates the progression of kidney injury via Na/H exchanger inhibition in 5/6 nephrectomized rats. J. Korean Med. Sci. 2014, 29, 691–698. [Google Scholar] [CrossRef] [Green Version]
- Vella, J.; Zammit, C.; Di Giovanni, G.; Muscat, R.; Valentino, M. The central role of aquaporins in the pathophysiology of ischemic stroke. Front. Cell. Neurosci. 2015, 9, 108. [Google Scholar] [CrossRef] [Green Version]
- Nagelhus, E.A.; Ottersen, O.P. Physiological roles of aquaporin-4 in brain. Physiol. Rev. 2013, 93, 1543–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manley, G.T.; Fujimura, M.; Ma, T.; Noshita, N.; Filiz, F.; Bollen, A.W.; Chan, P.; Verkman, A.S. Aquaporin-4 deletion in mice reduces brain edema after acute water intoxication and ischemic stroke. Nat. Med. 2000, 6, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Stavale, L.M.; Soares, E.S.; Mendonça, M.C.; Irazusta, S.P.; da Cruz Höfling, M.A. Temporal relationship between aquaporin-4 and glial fibrillary acidic protein in cerebellum of neonate and adult rats administered a BBB disrupting spider venom. Toxicon 2013, 66, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montagne, A.; Barnes, S.R.; Sweeney, M.D.; Halliday, M.R.; Sagare, A.P.; Zhao, Z.; Toga, A.W.; Jacobs, R.E.; Liu, C.Y.; Amezcua, L.; et al. Blood-brain barrier breakdown in the aging human hippocampus. Neuron 2015, 85, 296–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montagne, A.; Huuskonen, M.T.; Rajagopal, G.; Sweeney, M.D.; Nation, D.A.; Sepehrband, F.; D’Orazio, L.M.; Harrington, M.G.; Chui, H.C.; Law, M.; et al. Undetectable gadolinium brain retention in individuals with an age-dependent blood-brain barrier breakdown in the hippocampus and mild cognitive impairment. Alzheimer’s Dement 2019, 15, 1568–1575. [Google Scholar] [CrossRef]
- Nation, D.A.; Sweeney, M.D.; Montagne, A.; Sagare, A.P.; D’Orazio, L.M.; Pachicano, M.; Sepehrband, F.; Nelson, A.R.; Buennagel, D.P.; Harrington, M.G.; et al. Blood-brain barrier breakdown is an early biomarker of human cognitive dysfunction. Nat. Med. 2019, 25, 270–276. [Google Scholar] [CrossRef]
- Bobot, M.; Thomas, L.; Moyon, A.; Fernandez, S.; McKay, N.; Balasse, L.; Garrigue, P.; Brige, P.; Chopinet, S.; Poitevin, S.; et al. Uremic Toxic Blood-Brain Barrier Disruption Mediated by AhR Activation Leads to Cognitive Impairment during Experimental Renal Dysfunction. J. Am. Soc. Nephrol. 2020, 31, 1509–1521. [Google Scholar] [CrossRef]
- Shao, X.; Lu, W.; Gao, F.; Li, D.; Hu, J.; Li, Y.; Zuo, Z.; Jie, H.; Zhao, Y.; Cen, X. Uric Acid Induces Cognitive Dysfunction through Hippocampal Inflammation in Rodents and Humans. J. Neurosci. 2016, 36, 10990–11005. [Google Scholar] [CrossRef] [Green Version]
- Mazumder, M.K.; Paul, R.; Bhattacharya, P.; Borah, A. Neurological sequel of chronic kidney disease: From diminished Acetylcholinesterase activity to mitochondrial dysfunctions, oxidative stress and inflammation in mice brain. Sci. Rep. 2019, 9, 3097. [Google Scholar] [CrossRef] [Green Version]
- Oh, K.H.; Kang, M.; Kang, E.; Ryu, H.; Han, S.H.; Yoo, T.H.; Kim, S.W.; Chae, D.W.; Lee, K.B.; Park, S.K.; et al. The KNOW-CKD Study: What we have learned about chronic kidney diseases. Kidney Res. Clin. Pract. 2020, 39, 121–135. [Google Scholar] [CrossRef]
- Toda, T.; Parylak, S.L.; Linker, S.B.; Gage, F.H. The role of adult hippocampal neurogenesis in brain health and disease. Mol. Psychiatry 2019, 24, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Allebone, J.; Kanaan, R.; Maller, J.; O’Brien, T.; Mullen, S.A.; Cook, M.; Adams, S.J.; Vogrin, S.; Vaughan, D.N.; Connelly, A.; et al. Bilateral volume reduction in posterior hippocampus in psychosis of epilepsy. J. Neurol. Neurosurg. Psychiatry 2019, 90, 688–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Flores, R.; La Joie, R.; Chételat, G. Structural imaging of hippocampal subfields in healthy aging and Alzheimer’s disease. Neuroscience 2015, 309, 29–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras, A.; Polín, E.; Miguéns, M.; Pérez-García, C.; Pérez, V.; Ruiz-Gayo, M.; Morales, L.; Del Olmo, N. Intermittent-Excessive and Chronic-Moderate Ethanol Intake during Adolescence Impair Spatial Learning, Memory and Cognitive Flexibility in the Adulthood. Neuroscience 2019, 418, 205–217. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Y.H.; Kim, S.-W.; Im, H.; Oh, S.W.; Cho, N.-J.; Park, S.; Park, D.-K.; Kim, D.-S.; Gil, H.-W. Cognitive Sequelae and Hippocampal Dysfunction in Chronic Kidney Disease following 5/6 Nephrectomy. Brain Sci. 2022, 12, 905. https://doi.org/10.3390/brainsci12070905
Yu YH, Kim S-W, Im H, Oh SW, Cho N-J, Park S, Park D-K, Kim D-S, Gil H-W. Cognitive Sequelae and Hippocampal Dysfunction in Chronic Kidney Disease following 5/6 Nephrectomy. Brain Sciences. 2022; 12(7):905. https://doi.org/10.3390/brainsci12070905
Chicago/Turabian StyleYu, Yeon Hee, Seong-Wook Kim, Hyuna Im, Se Won Oh, Nam-Jun Cho, Samel Park, Dae-Kyoon Park, Duk-Soo Kim, and Hyo-Wook Gil. 2022. "Cognitive Sequelae and Hippocampal Dysfunction in Chronic Kidney Disease following 5/6 Nephrectomy" Brain Sciences 12, no. 7: 905. https://doi.org/10.3390/brainsci12070905
APA StyleYu, Y. H., Kim, S. -W., Im, H., Oh, S. W., Cho, N. -J., Park, S., Park, D. -K., Kim, D. -S., & Gil, H. -W. (2022). Cognitive Sequelae and Hippocampal Dysfunction in Chronic Kidney Disease following 5/6 Nephrectomy. Brain Sciences, 12(7), 905. https://doi.org/10.3390/brainsci12070905