
Post-Mortem Analysis of Neuropathological Changes in Human Tinnitus


 , ,
, , 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimens
2.2. Histology and Immunohistochemistry
2.2.1. Cresyl Violet (Nissl) Staining
2.2.2. Immunohistochemistry
2.3. Quantification
2.3.1. Defined Regions of Interest
2.3.2. Quantification of the Nissl-Stained Cells
2.3.3. Quantification of the Glial Cells
2.3.4. Quantification of the Serotonergic and Dopaminergic Cells
2.4. Statistics
3. Results
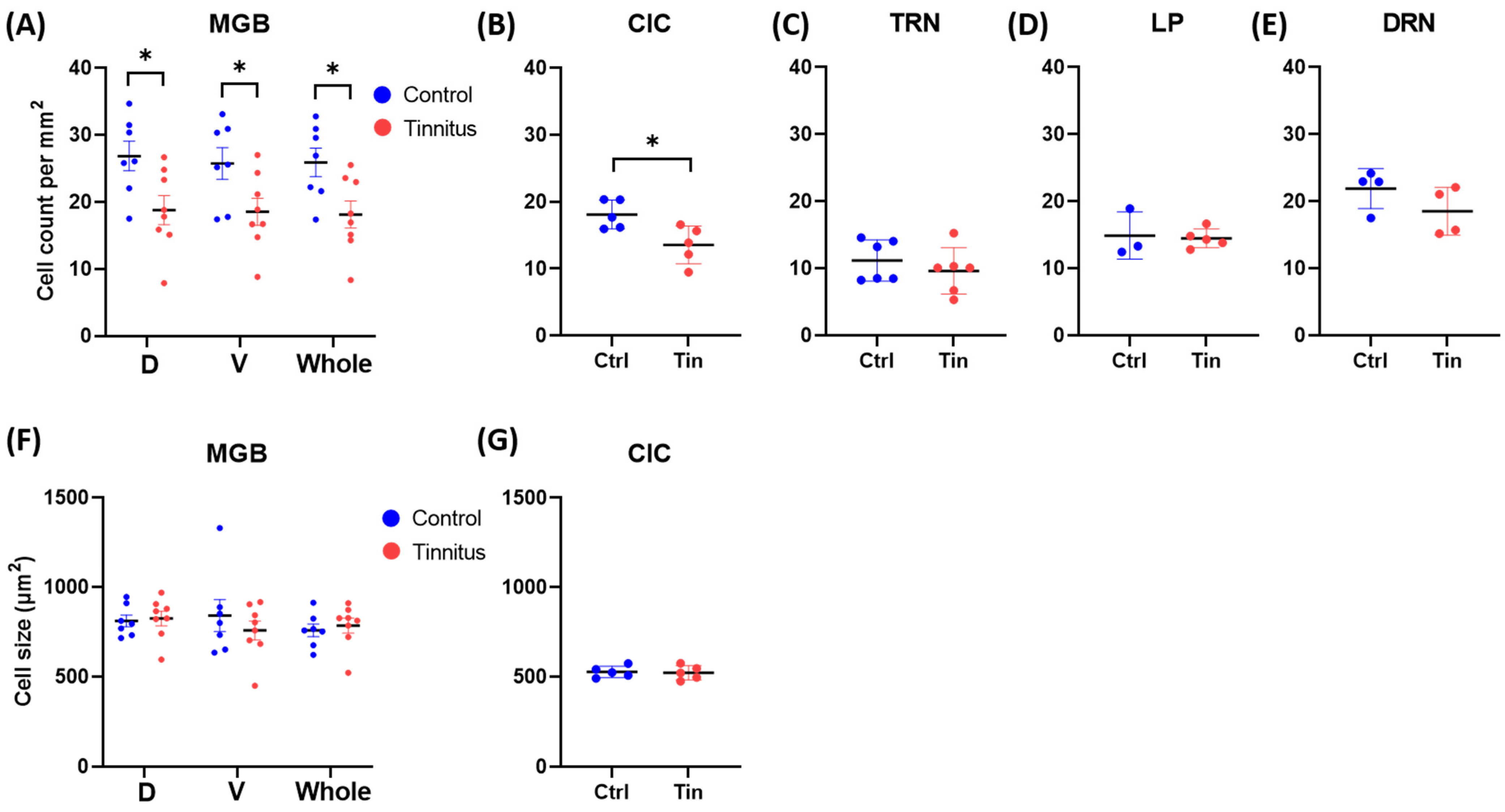
3.1. Cell Count and Size
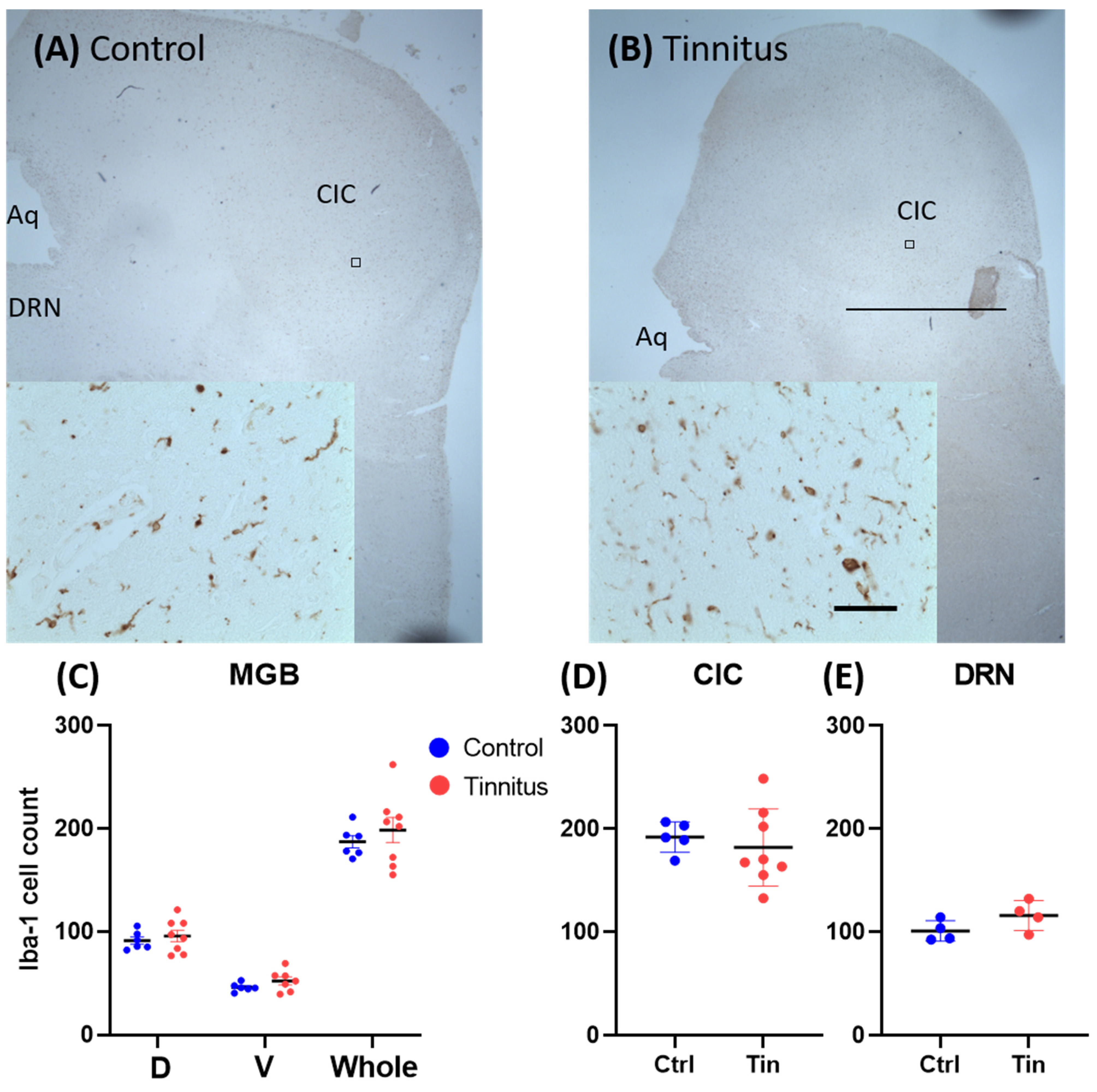
3.2. Glial Cells
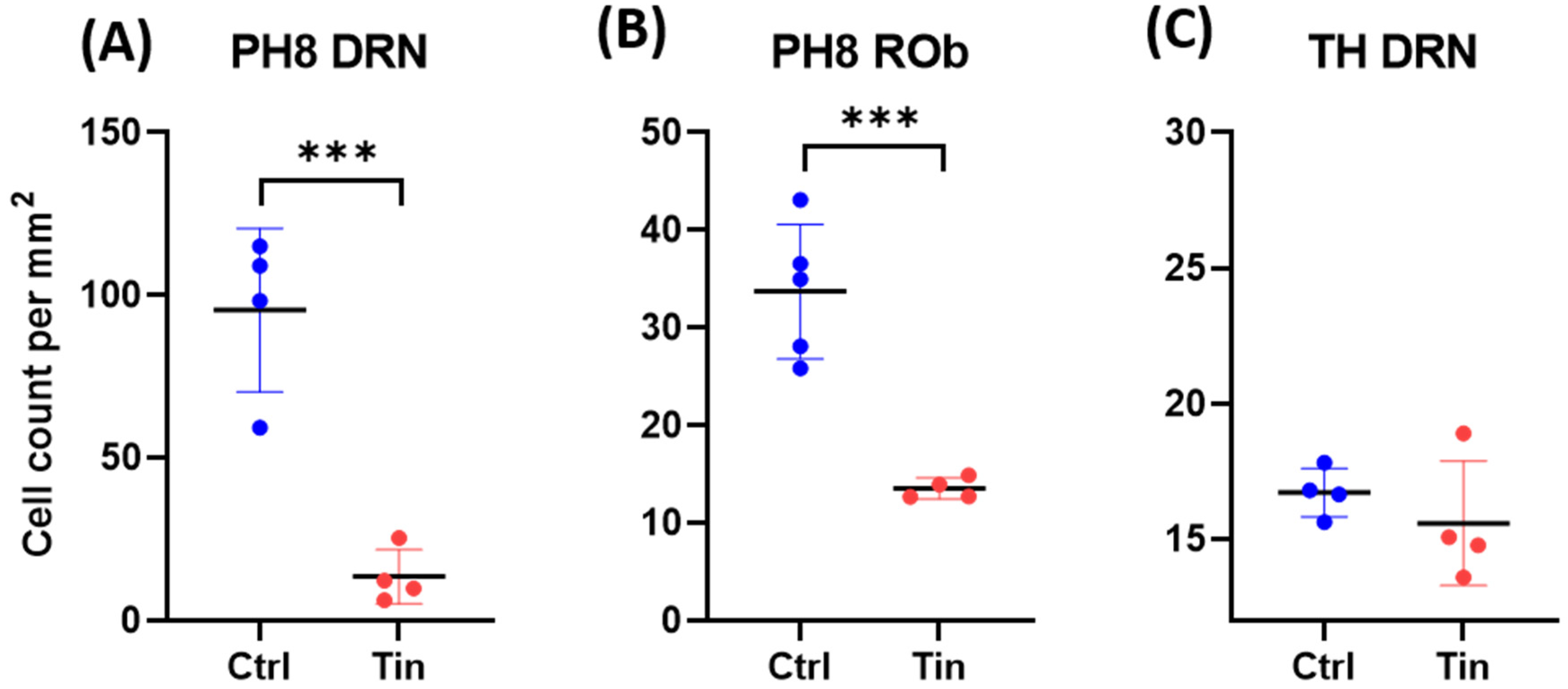
3.3. Serotonergic Cells
3.4. Dopaminergic Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Biswas, R.; Lugo, A.; Akeroyd, M.A.; Schlee, W.; Gallus, S.; Hall, D.A. Tinnitus prevalence in Europe: A multi-country cross-sectional population study. Lancet Reg. Health Eur. 2022, 12, 100250. [Google Scholar] [CrossRef] [PubMed]
- Durai, M.; Searchfield, G. Anxiety and depression, personality traits relevant to tinnitus: A scoping review. Int. J. Audiol. 2016, 55, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, Y.; Platz, E.A.; Niparko, J.K. Prevalence of hearing loss and differences by demographic characteristics among US adults: Data from the National Health and Nutrition Examination Survey, 1999–2004. Arch. Intern. Med. 2008, 168, 1522–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrawal, Y.; Platz, E.A.; Niparko, J.K. Risk factors for hearing loss in US adults: Data from the National Health and Nutrition Examination Survey, 1999 to 2002. Otol. Neurotol. 2009, 30, 139–145. [Google Scholar] [CrossRef]
- Eggermont, J.J.; Tass, P.A. Maladaptive neural synchrony in tinnitus: Origin and restoration. Front. Neurol. 2015, 6, 29. [Google Scholar] [CrossRef] [Green Version]
- Koops, E.A.; Eggermont, J.J. The thalamus and tinnitus: Bridging the gap between animal data and findings in humans. Hear. Res. 2021, 407, 108280. [Google Scholar] [CrossRef]
- Shore, S.E.; Wu, C. Mechanisms of Noise-Induced Tinnitus: Insights from Cellular Studies. Neuron 2019, 103, 8–20. [Google Scholar] [CrossRef]
- Zhang, J.S.; Kaltenbach, J.A.; Wang, J.; Kim, S.A. Fos-like immunoreactivity in auditory and nonauditory brain structures of hamsters previously exposed to intense sound. Exp. Brain Res. 2003, 153, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Luo, H.; Pace, E.; Li, L.; Liu, B. Psychophysical and neural correlates of noised-induced tinnitus in animals: Intra- and inter-auditory and non-auditory brain structure studies. Hear. Res. 2016, 334, 7–19. [Google Scholar] [CrossRef]
- Middleton, J.W.; Kiritani, T.; Pedersen, C.; Turner, J.G.; Shepherd, G.M.; Tzounopoulos, T. Mice with behavioral evidence of tinnitus exhibit dorsal cochlear nucleus hyperactivity because of decreased GABAergic inhibition. Proc. Natl. Acad. Sci. USA 2011, 108, 7601–7606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Weiner, B.D.; Zhang, L.S.; Cho, S.J.; Bao, S. Homeostatic plasticity drives tinnitus perception in an animal model. Proc. Natl. Acad. Sci. USA 2011, 108, 14974–14979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyakawa, A.; Wang, W.; Cho, S.-J.; Li, D.; Yang, S.; Bao, S. Tinnitus correlates with downregulation of cortical glutamate decarboxylase 65 expression but not auditory cortical map reorganization. J. Neurosci. 2019, 39, 9989–10001. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; Xia, C.; Wu, C.; Ji, Y.; Zhou, Y. Aberrant expression of Nav1.6 in the cochlear nucleus correlates with salicylate-induced tinnitus in rats. Biochem. Biophys. Res. Commun. 2020, 526, 786–792. [Google Scholar] [CrossRef] [PubMed]
- Heeringa, A.N.; Wu, C.; Chung, C.; West, M.; Martel, D.; Liberman, L.; Liberman, M.C.; Shore, S.E. Glutamatergic Projections to the Cochlear Nucleus are Redistributed in Tinnitus. Neuroscience 2018, 391, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Adjamian, P.; Hall, D.A.; Palmer, A.R.; Allan, T.W.; Langers, D.R. Neuroanatomical abnormalities in chronic tinnitus in the human brain. Neurosci. Biobehav. Rev. 2014, 45, 119–133. [Google Scholar] [CrossRef] [Green Version]
- Elgoyhen, A.B.; Langguth, B.; De Ridder, D.; Vanneste, S. Tinnitus: Perspectives from human neuroimaging. Nat. Rev. Neurosci. 2015, 16, 632–642. [Google Scholar] [CrossRef]
- Almasabi, F.; Janssen, M.L.F.; Devos, J.; Moerel, M.; Schwartze, M.; Kotz, S.A.; Jahanshahi, A.; Temel, Y.; Smit, J.V. The role of the medial geniculate body of the thalamus in the pathophysiology of tinnitus and implications for treatment. Brain Res. 2022, 1779, 147797. [Google Scholar] [CrossRef]
- Schneider, P.; Andermann, M.; Wengenroth, M.; Goebel, R.; Flor, H.; Rupp, A.; Diesch, E. Reduced volume of Heschl’s gyrus in tinnitus. Neuroimage 2009, 45, 927–939. [Google Scholar] [CrossRef]
- Aldhafeeri, F.M.; Mackenzie, I.; Kay, T.; Alghamdi, J.; Sluming, V. Neuroanatomical correlates of tinnitus revealed by cortical thickness analysis and diffusion tensor imaging. Neuroradiology 2012, 54, 883–892. [Google Scholar] [CrossRef]
- Muhlau, M.; Rauschecker, J.P.; Oestreicher, E.; Gaser, C.; Rottinger, M.; Wohlschlager, A.M.; Simon, F.; Etgen, T.; Conrad, B.; Sander, D. Structural brain changes in tinnitus. Cereb. Cortex 2006, 16, 1283–1288. [Google Scholar] [CrossRef]
- Landgrebe, M.; Langguth, B.; Rosengarth, K.; Braun, S.; Koch, A.; Kleinjung, T.; May, A.; de Ridder, D.; Hajak, G. Structural brain changes in tinnitus: Grey matter decrease in auditory and non-auditory brain areas. Neuroimage 2009, 46, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Tae, W.S.; Yakunina, N.; Lee, W.H.; Ryu, Y.J.; Ham, H.K.; Pyun, S.B.; Nam, E.C. Changes in the regional shape and volume of subcortical nuclei in patients with tinnitus comorbid with mild hearing loss. Neuroradiology 2018, 60, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Leaver, A.M.; Renier, L.; Chevillet, M.A.; Morgan, S.; Kim, H.J.; Rauschecker, J.P. Dysregulation of limbic and auditory networks in tinnitus. Neuron 2011, 69, 33–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyen, K.; Langers, D.R.; de Kleine, E.; van Dijk, P. Gray matter in the brain: Differences associated with tinnitus and hearing loss. Hear. Res. 2013, 295, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Zhang, J.; Li, X.W.; Xia, W.; Feng, X.; Gao, B.; Ju, S.H.; Wang, J.; Salvi, R.; Teng, G.J. Aberrant spontaneous brain activity in chronic tinnitus patients revealed by resting-state functional MRI. NeuroImage Clin. 2014, 6, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Chen, Y.C.; Feng, X.; Yang, M.; Liu, B.; Qian, C.; Wang, J.; Salvi, R.; Teng, G.J. Impairments of thalamic resting-state functional connectivity in patients with chronic tinnitus. Eur. J. Radiol. 2015, 84, 1277–1284. [Google Scholar] [CrossRef]
- Adams, M.E.; Huang, T.C.; Nagarajan, S.; Cheung, S.W. Tinnitus Neuroimaging. Otolaryngol. Clin. N. Am. 2020, 53, 583–603. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.J.; Davies, W.E. A review of evidence in support of a role for 5-HT in the perception of tinnitus. Hear. Res. 2000, 145, 1–7. [Google Scholar] [CrossRef]
- Leonard, B.E. Serotonin receptors and their function in sleep, anxiety disorders and depression. Psychother. Psychosom. 1996, 65, 66–75. [Google Scholar] [CrossRef]
- Bhatt, J.M.; Bhattacharyya, N.; Lin, H.W. Relationships between tinnitus and the prevalence of anxiety and depression. Laryngoscope 2017, 127, 466–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zöger, S.; Svedlund, J.; Holgers, K.M. The effects of sertraline on severe tinnitus suffering—A randomized, double-blind, placebo-controlled study. J. Clin. Psychopharmacol. 2006, 26, 32–39. [Google Scholar] [CrossRef]
- Andorn, A.C.; Vittorio, J.A.; Bellflower, J. 3H-spiroperidol binding in human temporal cortex (Brodmann areas 41–42) occurs at multiple high affinity states with serotonergic selectivity. Psychopharmacology 1989, 99, 520–525. [Google Scholar] [CrossRef] [PubMed]
- Klepper, A.; Herbert, H. Distribution and origin of noradrenergic and serotonergic fibers in the cochlear nucleus and inferior colliculus of the rat. Brain Res. 1991, 557, 190–201. [Google Scholar] [CrossRef]
- Lavoie, B.; Parent, A. Serotoninergic innervation of the thalamus in the primate: An immunohistochemical study. J. Comp. Neurol. 1991, 312, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Wallhausser-Franke, E. Salicylate evokes c-fos expression in the brain stem: Implications for tinnitus. Neuroreport 1997, 8, 725–728. [Google Scholar] [CrossRef] [PubMed]
- Caperton, K.K.; Thompson, A.M. Activation of serotonergic neurons during salicylate-induced tinnitus. Otol. Neurotol. 2011, 32, 301–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Li, X.; Wang, L.; Dong, Y.; Han, H.; Liu, G. Effects of salicylate on serotoninergic activities in rat inferior colliculus and auditory cortex. Hear. Res. 2003, 175, 45–53. [Google Scholar] [CrossRef]
- Rinne, J.O.; Mlic, J.R.; Paljärvi, L.; Rinne, U.K. Dementia in Parkinson’s disease is related to neuronal loss in the medial substantia nigra. Ann. Neurol. Off. J. Am. Neurol. Assoc. Child Neurol. Soc. 1989, 26, 47–50. [Google Scholar] [CrossRef]
- Berman, R.A.; Wurtz, R.H. Exploring the pulvinar path to visual cortex. Prog. Brain Res. 2008, 171, 467–473. [Google Scholar] [CrossRef] [Green Version]
- Ding, S.L.; Royall, J.J.; Sunkin, S.M.; Ng, L.; Facer, B.A.; Lesnar, P.; Guillozet-Bongaarts, A.; McMurray, B.; Szafer, A.; Dolbeare, T.A.; et al. Comprehensive cellular-resolution atlas of the adult human brain. J. Comp. Neurol. 2016, 524, 3127–3481. [Google Scholar] [CrossRef]
- Jensen, E.B.; Gundersen, H.J.G. Stereological Estimation of Surface Area of Arbitrary Particles. In Proceedings of the 7th International Congress for Stereology (7-Ics), Caen, France, 2–9 September 1987; pp. 25–30. [Google Scholar]
- Haan, E.A.; Jennings, I.G.; Cuello, A.C.; Nakata, H.; Fujisawa, H.; Chow, C.W.; Kushinsky, R.; Brittingham, J.; Cotton, R.G. Identification of serotonergic neurons in human brain by a monoclonal antibody binding to all three aromatic amino acid hydroxylases. Brain Res. 1987, 426, 19–27. [Google Scholar] [CrossRef]
- Törk, I.; Halliday, G.M.; Cotton, R.G. Application of antiphenylalanine hydroxylase antibodies to the study of the serotonergic system in the human brain. J. Chem. Neuroanat. 1992, 5, 311–313. [Google Scholar] [CrossRef]
- Jahanshahi, A.; Vlamings, R.; van Roon-Mom, W.M.C.; Faull, R.L.M.; Waldvogel, H.J.; Janssen, M.L.F.; Yakkioui, Y.; Zeef, D.H.; Kocabicak, E.; Steinbusch, H.W.M.; et al. Changes in brainstem serotonergic and dopaminergic cell populations in experimental and clinical Huntington’s disease. Neuroscience 2013, 238, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Basta, D.; Tzschentke, B.; Ernst, A. Noise-induced cell death in the mouse medial geniculate body and primary auditory cortex. Neurosci. Lett. 2005, 381, 199–204. [Google Scholar] [CrossRef]
- Groschel, M.; Gotze, R.; Ernst, A.; Basta, D. Differential impact of temporary and permanent noise-induced hearing loss on neuronal cell density in the mouse central auditory pathway. J. Neurotrauma 2010, 27, 1499–1507. [Google Scholar] [CrossRef]
- Chu, H.T.; Liang, C.S.; Yeh, T.C.; Hu, L.Y.; Yang, A.C.; Tsai, S.J.; Shen, C.C. Tinnitus and risk of Alzheimer’s and Parkinson’s disease: A retrospective nationwide population-based cohort study. Sci. Rep. 2020, 10, 12134. [Google Scholar] [CrossRef]
- Shulman, A.; Goldstein, B.; Strashun, A.M. Central nervous system neurodegeneration and tinnitus: A clinical experience. Part I: Diagnosis. Int. Tinnitus J. 2007, 13, 118–131. [Google Scholar]
- Sung, K.; Jimenez-Sanchez, M. Autophagy in Astrocytes and its Implications in Neurodegeneration. J. Mol. Biol. 2020, 432, 2605–2621. [Google Scholar] [CrossRef]
- Smith, D.H.; Chen, X.H.; Pierce, J.E.; Wolf, J.A.; Trojanowski, J.Q.; Graham, D.I.; McIntosh, T.K. Progressive atrophy and neuron death for one year following brain trauma in the rat. J. Neurotrauma 1997, 14, 715–727. [Google Scholar] [CrossRef]
- Rajkowska, G.; Stockmeier, C.A. Astrocyte pathology in major depressive disorder: Insights from human postmortem brain tissue. Curr. Drug Targets 2013, 14, 1225–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ongür, D.; Drevets, W.C.; Price, J.L. Glial reduction in the subgenual prefrontal cortex in mood disorders. Proc. Natl. Acad. Sci. USA 1998, 95, 13290–13295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajkowska, G.; Miguel-Hidalgo, J.J.; Wei, J.; Dilley, G.; Pittman, S.D.; Meltzer, H.Y.; Overholser, J.C.; Roth, B.L.; Stockmeier, C.A. Morphometric evidence for neuronal and glial prefrontal cell pathology in major depression. Biol. Psychiatry 1999, 45, 1085–1098. [Google Scholar] [CrossRef]
- Bowley, M.P.; Drevets, W.C.; Ongür, D.; Price, J.L. Low glial numbers in the amygdala in major depressive disorder. Biol. Psychiatry 2002, 52, 404–412. [Google Scholar] [CrossRef]
- Cotter, D.; Mackay, D.; Chana, G.; Beasley, C.; Landau, S.; Everall, I.P. Reduced neuronal size and glial cell density in area 9 of the dorsolateral prefrontal cortex in subjects with major depressive disorder. Cereb. Cortex 2002, 12, 386–394. [Google Scholar] [CrossRef]
- Xia, C.; Yin, M.; Wu, C.; Ji, Y.; Zhou, Y. Neuroglial activation in the auditory cortex and medial geniculate body of salicylate-induced tinnitus rats. Am. J. Transl. Res. 2020, 12, 6043–6059. [Google Scholar]
- Wang, W.; Zhang, L.S.; Zinsmaier, A.K.; Patterson, G.; Leptich, E.J.; Shoemaker, S.L.; Yatskievych, T.A.; Gibboni, R.; Pace, E.; Luo, H.; et al. Neuroinflammation mediates noise-induced synaptic imbalance and tinnitus in rodent models. PLoS Biol. 2019, 17, e3000307. [Google Scholar] [CrossRef]
- Mennink, L.M.; Aalbers, M.W.; van Dijk, P.; van Dijk, J.M.C. The Role of Inflammation in Tinnitus: A Systematic Review and Meta-Analysis. J. Clin. Med. 2022, 11, 1000. [Google Scholar] [CrossRef]
- Pattyn, T.; Van Den Eede, F.; Vanneste, S.; Cassiers, L.; Veltman, D.J.; Van De Heyning, P.; Sabbe, B.C.G. Tinnitus and anxiety disorders: A review. Hear. Res. 2016, 333, 255–265. [Google Scholar] [CrossRef]
- Salazar, J.W.; Meisel, K.; Smith, E.R.; Quiggle, A.; McCoy, D.B.; Amans, M.R. Depression in Patients with Tinnitus: A Systematic Review. Otolaryngol. Head Neck Surg. 2019, 161, 28–35. [Google Scholar] [CrossRef]
- Alster, J.; Shemesh, Z.; Ornan, M.; Attias, J. Sleep disturbance associated with chronic tinnitus. Biol. Psychiatry 1993, 34, 84–90. [Google Scholar] [CrossRef]
- Baldo, P.; Doree, C.; Molin, P.; McFerran, D.; Cecco, S. Antidepressants for patients with tinnitus. Cochrane Database Syst. Rev. 2012, 2012, CD003853. [Google Scholar] [CrossRef]
- Folmer, R.L.; Theodoroff, S.M.; Martin, W.H.; Shi, Y. Experimental, controversial, and futuristic treatments for chronic tinnitus. J. Am. Acad. Audiol. 2014, 25, 106–125. [Google Scholar] [CrossRef]
- Rauschecker, J.P.; Leaver, A.M.; Muhlau, M. Tuning out the noise: Limbic-auditory interactions in tinnitus. Neuron 2010, 66, 819–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stutzmann, G.E.; McEwen, B.S.; LeDoux, J.E. Serotonin modulation of sensory inputs to the lateral amygdala: Dependency on corticosterone. J. Neurosci. 1998, 18, 9529–9538. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.Q.; Trussell, L.O. Serotonergic Modulation of Sensory Representation in a Central Multisensory Circuit Is Pathway Specific. Cell Rep. 2017, 20, 1844–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervantes-Ramírez, V.; Canto-Bustos, M.; Aguilar-Magaña, D.; Pérez-Padilla, E.A.; Góngora-Alfaro, J.L.; Pineda, J.C.; Atzori, M.; Salgado, H. Citalopram reduces glutamatergic synaptic transmission in the auditory cortex via activation of 5-HT1A receptors. Neuroreport 2019, 30, 1316–1322. [Google Scholar] [CrossRef] [PubMed]
- Rademaker, M.M.; Stegeman, I.; Brabers, A.E.M.; de Jong, J.D.; Stokroos, R.J.; Smit, A.L. Differences in characteristics between people with tinnitus that seek help and that do not. Sci. Rep. 2021, 11, 22949. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Case # | Duration | Disorder Characteristic | Sex | Age | pmd | Weight | MGB | TRN | LP | CIC | DRN | Rob |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C1 | S98/233 | M | 72 | 4 | 1385 | √ | √ | √ | √ | ||||
| C2 | S11/091 | M | 76 | 7 | 1220 | √ | √ | ||||||
| C3 | S14/063 | F | 93 | 8 | 1025 | √ | √ | √ | √ | ||||
| C4 | S17/016 | M | 72 | 4 | 1385 | √ | √ | √ | √ | √ | √ | ||
| C5 | S17/093 | M | 82 | 6 | 1195 | √ | √ | √ | √ | √ | |||
| C6 | PDC019 | F | 74 | 29 | 1071 | √ | √ | ||||||
| C7 | PDC033 | F | 77 | 43 | 1450 | √ | √ | √ | |||||
| C8 | PDC086 | M | 89 | 34 | 1209 | √ | √ | √ | √ | ||||
| Total | 7 | 6 | 3 | 5 | 4 | 5 | |||||||
| T1 | PDC028 | - | B with HL | F | 84 | 11 | 602 Hf | √ | √ | ||||
| T2 | S00/157 | 5 | B with HL | M | 78 | 8 | 1222 | √ | √ | √ | |||
| T3 | S00/255 | 8 | B | F | 94 | 7 | 937 | √ | √ | √ | |||
| T4 | S01/119 | 7 | U | M | 75 | 6 | 1056 | √ | √ | √ | √ | ||
| T5 | S10/033 | 4 | U | M | 70 | 6 | 1502 | √ | √ | √ | √ | √ | √ |
| T6 | C033 | 8 | B with HL | M | 75 | 18 | 1519 | √ | √ | √ | √ | √ | |
| T7 | PDC015 | long standing | B with HL | F | 81 | 15 | 1352 | √ | √ | √ | √ | √ | |
| T8 | PDC029 | 15 | B with HL | M | 82 | 48 | 656 Hf | √ | √ | ||||
| T9 | PDC032 | 30 | B with HL | F | 91 | 19 | 1068 | √ | √ | ||||
| Total | 8 | 6 | 5 | 5 | 4 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almasabi, F.; Alosaimi, F.; Corrales-Terrón, M.; Wolters, A.; Strikwerda, D.; Smit, J.V.; Temel, Y.; Janssen, M.L.F.; Jahanshahi, A. Post-Mortem Analysis of Neuropathological Changes in Human Tinnitus. Brain Sci. 2022, 12, 1024. https://doi.org/10.3390/brainsci12081024
Almasabi F, Alosaimi F, Corrales-Terrón M, Wolters A, Strikwerda D, Smit JV, Temel Y, Janssen MLF, Jahanshahi A. Post-Mortem Analysis of Neuropathological Changes in Human Tinnitus. Brain Sciences. 2022; 12(8):1024. https://doi.org/10.3390/brainsci12081024
Chicago/Turabian StyleAlmasabi, Faris, Faisal Alosaimi, Minerva Corrales-Terrón, Anouk Wolters, Dario Strikwerda, Jasper V. Smit, Yasin Temel, Marcus L. F. Janssen, and Ali Jahanshahi. 2022. "Post-Mortem Analysis of Neuropathological Changes in Human Tinnitus" Brain Sciences 12, no. 8: 1024. https://doi.org/10.3390/brainsci12081024
APA StyleAlmasabi, F., Alosaimi, F., Corrales-Terrón, M., Wolters, A., Strikwerda, D., Smit, J. V., Temel, Y., Janssen, M. L. F., & Jahanshahi, A. (2022). Post-Mortem Analysis of Neuropathological Changes in Human Tinnitus. Brain Sciences, 12(8), 1024. https://doi.org/10.3390/brainsci12081024