
The Effect of Noise Trauma and Deep Brain Stimulation of the Medial Geniculate Body on Tissue Activity in the Auditory Pathway

 ,
,  ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Animals
2.2. Noise Trauma
2.3. Gap Prepulse Inhibition of Acoustic Startles
2.4. Surgical Procedures
2.5. Deep Brain Stimulation
2.6. Tissue Collection
2.7. Histochemistry and Immunohistochemistry
2.7.1. Cytochrome C Oxidase Histochemistry
2.7.2. C-Fos Immunohistochemistry
2.8. Statistical Analysis
3. Results
3.1. Cytochrome C Oxidase Density
3.2. C-Fos Cell Count
4. Discussion
4.1. Effect of Noise Trauma on Activity Markers
4.2. Effect of the High-Frequency Stimulation in the Medial Geniculate Body on Neuronal Activity Markers
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biswas, R.; Lugo, A.; Akeroyd, M.A.; Schlee, W.; Gallus, S.; Hall, D.A. Tinnitus prevalence in Europe: A multi-country cross-sectional population study. Lancet Reg. Health Eur. 2022, 12, 100250. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, J.M.; Bhattacharyya, N.; Lin, H.W. Relationships between tinnitus and the prevalence of anxiety and depression. Laryngoscope 2017, 127, 466–469. [Google Scholar] [CrossRef] [PubMed]
- Vanneste, S.; De Ridder, D. Noninvasive and invasive neuromodulation for the treatment of tinnitus: An overview. Neuromodulation J. Int. Neuromodulation Soc. 2012, 15, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Smit, J.V.; Janssen, M.L.; Schulze, H.; Jahanshahi, A.; Van Overbeeke, J.J.; Temel, Y.; Stokroos, R.J. Deep brain stimulation in tinnitus: Current and future perspectives. Brain Res. 2015, 1608, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Lozano, A.M.; Lipsman, N.; Bergman, H.; Brown, P.; Chabardes, S.; Chang, J.W.; Matthews, K.; McIntyre, C.C.; Schlaepfer, T.E.; Schulder, M.; et al. Deep brain stimulation: Current challenges and future directions. Nat. Rev. Neurol. 2019, 15, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Zhang, X.; Nation, J.; Pace, E.; Lepczyk, L.; Zhang, J. Tinnitus suppression by electrical stimulation of the rat dorsal cochlear nucleus. Neurosci. Lett. 2012, 522, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Smit, J.V.; Janssen, M.L.; van Zwieten, G.; Jahanshahi, A.; Temel, Y.; Stokroos, R.J. Deep brain stimulation of the inferior colliculus in the rodent suppresses tinnitus. Brain Res. 2016, 1650, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Van Zwieten, G.; Jahanshahi, A.; van Erp, M.L.; Temel, Y.; Stokroos, R.J.; Janssen, M.L.F.; Smit, J.V. Alleviation of Tinnitus With High-Frequency Stimulation of the Dorsal Cochlear Nucleus: A Rodent Study. Trends Hear. 2019, 23, 2331216519835080. [Google Scholar] [CrossRef] [PubMed]
- Van Zwieten, G.; Janssen, M.L.F.; Smit, J.V.; Janssen, A.M.L.; Roet, M.; Jahanshahi, A.; Stokroos, R.J.; Temel, Y. Inhibition of Experimental Tinnitus With High Frequency Stimulation of the Rat Medial Geniculate Body. Neuromodulation J. Int. Neuromodulation Soc. 2019, 22, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, P.; Kotz, S.A.; Smit, J.V.; Janssen, M.L.F.; Schwartze, M. Auditory thalamus dysfunction and pathophysiology in tinnitus: A predictive network hypothesis. Brain Struct. Funct. 2021, 226, 1659–1676. [Google Scholar] [CrossRef] [PubMed]
- Koops, E.A.; Eggermont, J.J. The thalamus and tinnitus: Bridging the gap between animal data and findings in humans. Hear. Res. 2021, 407, 108280. [Google Scholar] [CrossRef] [PubMed]
- Almasabi, F.; Janssen, M.L.F.; Devos, J.; Moerel, M.; Schwartze, M.; Kotz, S.A.; Jahanshahi, A.; Temel, Y.; Smit, J.V. The role of the medial geniculate body of the thalamus in the pathophysiology of tinnitus and implications for treatment. Brain Res. 2022, 1779, 147797. [Google Scholar] [CrossRef] [PubMed]
- Van Zwieten, G.; Smit, J.V.; Jahanshahi, A.; Temel, Y.; Stokroos, R.J. Tinnitus: Is there a place for brain stimulation? Surg. Neurol. Int. 2016, 7, S125–S129. [Google Scholar] [CrossRef] [PubMed]
- Budzikowski, A.S.; Vahid-Ansari, F.; Leenen, F.H. Chronic activation of brain areas by high-sodium diet in Dahl salt-sensitive rats. Am. J. Physiol. 1998, 274, H2046–H2052. [Google Scholar] [CrossRef] [PubMed]
- Jahanshahi, A.; Schonfeld, L.; Janssen, M.L.; Hescham, S.; Kocabicak, E.; Steinbusch, H.W.; van Overbeeke, J.J.; Temel, Y. Electrical stimulation of the motor cortex enhances progenitor cell migration in the adult rat brain. Exp. Brain Res. 2013, 231, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Van Zwieten, G.; Roberts, M.J.; Schaper, F.; Smit, J.V.; Temel, Y.; Janssen, M.L.F. Noise-induced neurophysiological alterations in the rat medial geniculate body and thalamocortical desynchronization by deep brain stimulation. J. Neurophysiol. 2021, 125, 661–671. [Google Scholar] [CrossRef]
- Turner, J.G.; Brozoski, T.J.; Bauer, C.A.; Parrish, J.L.; Myers, K.; Hughes, L.F.; Caspary, D.M. Gap detection deficits in rats with tinnitus: A potential novel screening tool. Behav. Neurosci. 2006, 120, 188. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates: Hard Cover Edition; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Tan, S.; Vlamings, R.; Lim, L.; Sesia, T.; Janssen, M.L.; Steinbusch, H.W.; Visser-Vandewalle, V.; Temel, Y. Experimental deep brain stimulation in animal models. Neurosurgery 2010, 67, 1073–1079, discussion Neurosurgery 2010, 67, 1080. [Google Scholar] [CrossRef] [PubMed]
- Eggermont, J.J. Can Animal Models Contribute to Understanding Tinnitus Heterogeneity in Humans? Front. Aging Neurosci. 2016, 8, 265. [Google Scholar] [CrossRef]
- Von der Behrens, W. Animal models of subjective tinnitus. Neural Plast. 2014, 2014, 741452. [Google Scholar] [CrossRef]
- Kraus, K.S.; Canlon, B. Neuronal connectivity and interactions between the auditory and limbic systems. Effects of noise and tinnitus. Hear. Res. 2012, 288, 34–46. [Google Scholar] [CrossRef]
- Shore, S.E.; Roberts, L.E.; Langguth, B. Maladaptive plasticity in tinnitus--triggers, mechanisms and treatment. Nat. Rev. Neurol. 2016, 12, 150–160. [Google Scholar] [CrossRef]
- Shi, Y.; Burchiel, K.J.; Anderson, V.C.; Martin, W.H. Deep brain stimulation effects in patients with tinnitus. Otolaryngol. -Head Neck Surg. Off. J. Am. Acad. Otolaryngol.-Head Neck Surg. 2009, 141, 285–287. [Google Scholar] [CrossRef]
- Cheung, S.W.; Larson, P.S. Tinnitus modulation by deep brain stimulation in locus of caudate neurons (area LC). Neuroscience 2010, 169, 1768–1778. [Google Scholar] [CrossRef] [PubMed]
- Smit, J.V.; Janssen, M.L.; Engelhard, M.; de Bie, R.M.; Schuurman, P.R.; Contarino, M.F.; Mosch, A.; Temel, Y.; Stokroos, R.J. The impact of deep brain stimulation on tinnitus. Surg. Neurol. Int. 2016, 7, S848–S854. [Google Scholar] [CrossRef]
- Dijkstra, E.; Figee, M.; Schuurman, P.R.; Denys, D. Effective deep brain stimulation of intractable tinnitus: A case study. Brain Stimul. 2018, 11, 1205–1207. [Google Scholar] [CrossRef]
- Cheung, S.W.; Racine, C.A.; Henderson-Sabes, J.; Demopoulos, C.; Molinaro, A.M.; Heath, S.; Nagarajan, S.S.; Bourne, A.L.; Rietcheck, J.E.; Wang, S.S.; et al. Phase I trial of caudate deep brain stimulation for treatment-resistant tinnitus. J. Neurosurg. 2019, 133, 992–1001. [Google Scholar] [CrossRef] [PubMed]
- Muhlau, M.; Rauschecker, J.P.; Oestreicher, E.; Gaser, C.; Rottinger, M.; Wohlschlager, A.M.; Simon, F.; Etgen, T.; Conrad, B.; Sander, D. Structural brain changes in tinnitus. Cerebral Cortex 2006, 16, 1283–1288. [Google Scholar] [CrossRef] [PubMed]
- Landgrebe, M.; Langguth, B.; Rosengarth, K.; Braun, S.; Koch, A.; Kleinjung, T.; May, A.; de Ridder, D.; Hajak, G. Structural brain changes in tinnitus: Grey matter decrease in auditory and non-auditory brain areas. Neuroimage 2009, 46, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Boyen, K.; Langers, D.R.; de Kleine, E.; van Dijk, P. Gray matter in the brain: Differences associated with tinnitus and hearing loss. Hear. Res. 2013, 295, 67–78. [Google Scholar] [CrossRef]
- Elgoyhen, A.B.; Langguth, B.; De Ridder, D.; Vanneste, S. Tinnitus: Perspectives from human neuroimaging. Nat. Rev. Neurosci. 2015, 16, 632–642. [Google Scholar] [CrossRef] [PubMed]
- Seki, S.; Eggermont, J.J. Changes in spontaneous firing rate and neural synchrony in cat primary auditory cortex after localized tone-induced hearing loss. Hear. Res. 2003, 180, 28–38. [Google Scholar] [CrossRef]
- Weisz, N.; Dohrmann, K.; Elbert, T. The relevance of spontaneous activity for the coding of the tinnitus sensation. In Progress in Brain Research; Langguth, B., Hajak, G., Kleinjung, T., Cacace, A., Møller, A.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; Volume 166, pp. 61–70. [Google Scholar]
- Weisz, N.; Muller, S.; Schlee, W.; Dohrmann, K.; Hartmann, T.; Elbert, T. The neural code of auditory phantom perception. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 1479–1484. [Google Scholar] [CrossRef] [PubMed]
- Ortmann, M.; Müller, N.; Schlee, W.; Weisz, N. Rapid increases of gamma power in the auditory cortex following noise trauma in humans. Eur. J. Neurosci. 2011, 33, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Rauschecker, J.P.; Leaver, A.M.; Muhlau, M. Tuning out the noise: Limbic-auditory interactions in tinnitus. Neuron 2010, 66, 819–826. [Google Scholar] [CrossRef]
- De Ridder, D.; Vanneste, S.; Langguth, B.; Llinas, R. Thalamocortical Dysrhythmia: A Theoretical Update in Tinnitus. Front. Neurol. 2015, 6, 124. [Google Scholar] [CrossRef]
- Kalappa, B.I.; Brozoski, T.J.; Turner, J.G.; Caspary, D.M. Single unit hyperactivity and bursting in the auditory thalamus of awake rats directly correlates with behavioural evidence of tinnitus. J. Physiol. 2014, 592, 5065–5078. [Google Scholar] [CrossRef]
- Cook, J.A.; Barry, K.M.; Zimdahl, J.W.; Leggett, K.; Mulders, W. Spontaneous firing patterns in the medial geniculate nucleus in a guinea pig model of tinnitus. Hear. Res. 2021, 403, 108190. [Google Scholar] [CrossRef]
- Barry, K.M.; Robertson, D.; Mulders, W. Changes in auditory thalamus neural firing patterns after acoustic trauma in rats. Hear. Res. 2019, 379, 89–97. [Google Scholar] [CrossRef]
- Ueyama, T.; Donishi, T.; Ukai, S.; Ikeda, Y.; Hotomi, M.; Yamanaka, N.; Shinosaki, K.; Terada, M.; Kaneoke, Y. Brain regions responsible for tinnitus distress and loudness: A resting-state FMRI study. PLoS ONE 2013, 8, e67778. [Google Scholar] [CrossRef]
- Kapolowicz, M.R.; Thompson, L.T. Acute high-intensity noise induces rapid Arc protein expression but fails to rapidly change GAD expression in amygdala and hippocampus of rats: Effects of treatment with D-cycloserine. Hear. Res. 2016, 342, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wu, C.; Martel, D.T.; West, M.; Sutton, M.A.; Shore, S.E. Remodeling of cholinergic input to the hippocampus after noise exposure and tinnitus induction in Guinea pigs. Hippocampus 2019, 29, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wu, C.; Martel, D.T.; West, M.; Sutton, M.A.; Shore, S.E. Noise Exposure Alters Glutamatergic and GABAergic Synaptic Connectivity in the Hippocampus and Its Relevance to Tinnitus. Neural Plast. 2021, 2021, 8833087. [Google Scholar] [CrossRef] [PubMed]
- Lanting, C.P.; de Kleine, E.; van Dijk, P. Neural activity underlying tinnitus generation: Results from PET and fMRI. Hear. Res. 2009, 255, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.E.; Huang, T.C.; Nagarajan, S.; Cheung, S.W. Tinnitus Neuroimaging. Otolaryngol. Clin. N. Am. 2020, 53, 583–603. [Google Scholar] [CrossRef] [PubMed]
- Qu, T.; Qi, Y.; Yu, S.; Du, Z.; Wei, W.; Cai, A.; Wang, J.; Nie, B.; Liu, K.; Gong, S. Dynamic Changes of Functional Neuronal Activities between the Auditory Pathway and Limbic Systems Contribute to Noise-Induced Tinnitus with a Normal Audiogram. Neuroscience 2019, 408, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Hofmeier, B.; Wolpert, S.; Aldamer, E.S.; Walter, M.; Thiericke, J.; Braun, C.; Zelle, D.; Rüttiger, L.; Klose, U.; Knipper, M. Reduced sound-evoked and resting-state BOLD fMRI connectivity in tinnitus. NeuroImage Clin. 2018, 20, 637–649. [Google Scholar] [CrossRef]
- Chen, Y.C.; Zhang, J.; Li, X.W.; Xia, W.; Feng, X.; Gao, B.; Ju, S.H.; Wang, J.; Salvi, R.; Teng, G.J. Aberrant spontaneous brain activity in chronic tinnitus patients revealed by resting-state functional MRI. NeuroImage Clin. 2014, 6, 222–228. [Google Scholar] [CrossRef]
- Boyen, K.; de Kleine, E.; van Dijk, P.; Langers, D.R. Tinnitus-related dissociation between cortical and subcortical neural activity in humans with mild to moderate sensorineural hearing loss. Hear. Res. 2014, 312, 48–59. [Google Scholar] [CrossRef]
- Carpenter-Thompson, J.R.; Akrofi, K.; Schmidt, S.A.; Dolcos, F.; Husain, F.T. Alterations of the emotional processing system may underlie preserved rapid reaction time in tinnitus. Brain Res. 2014, 1567, 28–41. [Google Scholar] [CrossRef]
- Chen, Y.C.; Li, X.; Liu, L.; Wang, J.; Lu, C.Q.; Yang, M.; Jiao, Y.; Zang, F.C.; Radziwon, K.; Chen, G.D.; et al. Tinnitus and hyperacusis involve hyperactivity and enhanced connectivity in auditory-limbic-arousal-cerebellar network. eLife 2015, 4, e06576. [Google Scholar] [CrossRef] [PubMed]
- Ghazaleh, N.; Zwaag, W.V.; Clarke, S.; Ville, D.V.; Maire, R.; Saenz, M. High-Resolution fMRI of Auditory Cortical Map Changes in Unilateral Hearing Loss and Tinnitus. Brain Topogr. 2017, 30, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Berlot, E.; Arts, R.; Smit, J.; George, E.; Gulban, O.F.; Moerel, M.; Stokroos, R.; Formisano, E.; De Martino, F. A 7 Tesla fMRI investigation of human tinnitus percept in cortical and subcortical auditory areas. NeuroImage Clin. 2020, 25, 102166. [Google Scholar] [CrossRef] [PubMed]
- Wallhausser-Franke, E.; Mahlke, C.; Oliva, R.; Braun, S.; Wenz, G.; Langner, G. Expression of c-fos in auditory and non-auditory brain regions of the gerbil after manipulations that induce tinnitus. Exp. Brain Res. 2003, 153, 649–654. [Google Scholar] [CrossRef]
- Zhang, J.S.; Kaltenbach, J.A.; Wang, J.; Kim, S.A. Fos-like immunoreactivity in auditory and nonauditory brain structures of hamsters previously exposed to intense sound. Exp. Brain Res. 2003, 153, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.H.; Qin, Z.B. Expression of c-fos and NR2A in auditory cortex of rats experienced tinnitus. Chin. J. Otorhinolaryngol. Head Neck Surg. 2006, 41, 451–454. [Google Scholar]
- Ouda, L.; Jilek, M.; Syka, J. Expression of c-Fos in rat auditory and limbic systems following 22-kHz calls. Behav. Brain Res. 2016, 308, 196–204. [Google Scholar] [CrossRef]
- Zhang, J.; Luo, H.; Pace, E.; Li, L.; Liu, B. Psychophysical and neural correlates of noised-induced tinnitus in animals: Intra- and inter-auditory and non-auditory brain structure studies. Hear. Res. 2016, 334, 7–19. [Google Scholar] [CrossRef]
- Basura, G.J.; Koehler, S.D.; Shore, S.E. Bimodal stimulus timing-dependent plasticity in primary auditory cortex is altered after noise exposure with and without tinnitus. J. Neurophysiol. 2015, 114, 3064–3075. [Google Scholar] [CrossRef]
- Tan, S.K.; Janssen, M.L.; Jahanshahi, A.; Chouliaras, L.; Visser-Vandewalle, V.; Lim, L.W.; Steinbusch, H.W.; Sharp, T.; Temel, Y. High frequency stimulation of the subthalamic nucleus increases c-fos immunoreactivity in the dorsal raphe nucleus and afferent brain regions. J. Psychiatr. Res. 2011, 45, 1307–1315. [Google Scholar] [CrossRef]
- Takata, N. Thalamic reticular nucleus in the thalamocortical loop. Neurosci. Res. 2020, 156, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Aizenberg, M.; Rolon-Martinez, S.; Pham, T.; Rao, W.; Haas, J.S.; Geffen, M.N. Projection from the Amygdala to the Thalamic Reticular Nucleus Amplifies Cortical Sound Responses. Cell Rep. 2019, 28, 605–615.e604. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.S.; Lee, C.J.; Gibbs, J.W.; Coulter, D.A. Postnatal development of GABAA receptor function in somatosensory thalamus and cortex: Whole-cell voltage-clamp recordings in acutely isolated rat neurons. J. Neurosci. Off. J. Soc. Neurosci. 1995, 15, 1341–1351. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Region | Bregma | Optical Density | Statistical Outcome |
|---|---|---|---|
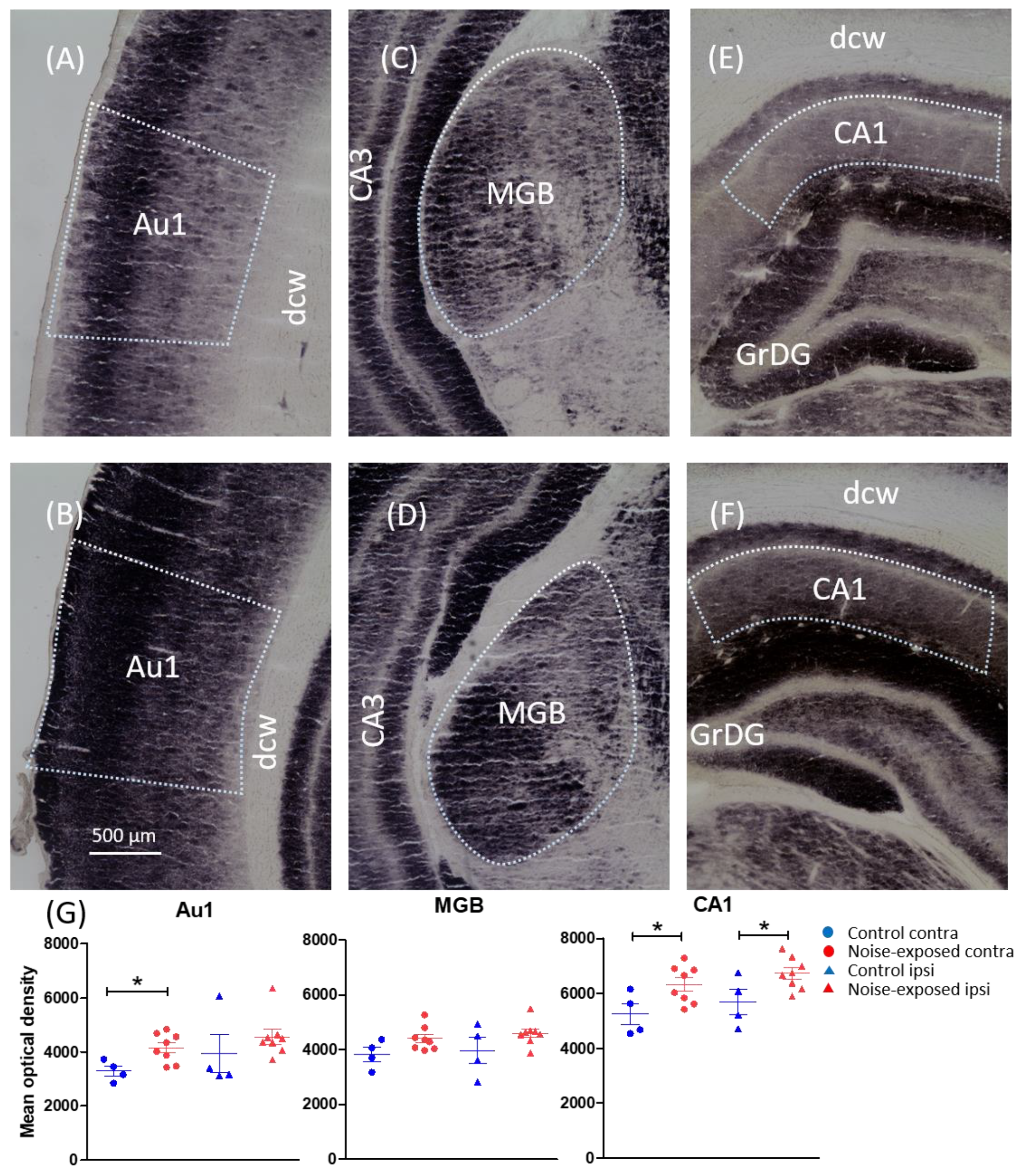
| Primary auditory cortex | −3.60, −4.08, −4.56 and −5.04 | C: 3619 (361) N: 4360 (171) | F(1, 20) = 4.570, p = 0.045 |
| Medial geniculate body | −5.04, −5.52, −6.00 and −6.48 | C: 3897 (250) N: 4503 (110) | F(1, 20) = 6.292, p = 0.021 |
| Inferior colliculus | −8.04, −8.52 and −9.00 | C: 3895 (356) N: 3756 (277) | F(1, 20) = 0.082, p = 0.778 |
| Dorsal cochlear nucleus | −10.68, −11.16 and −11.64 | C: 3328 (249) N: 3774 (132) | F(1, 20) = 2.946, p = 0.102 |
| CA1 of the hippocampus | −3.60, −4.56, −5.52 and −6.36 | C: 5471 (288) N: 6503 (172) | F(1, 20) = 10.73, p = 0.004 |
| CA3 of the hippocampus | −3.60, −4.56, −5.52 and −6.36 | C: 6640 (489) N: 7703 (276) | F(1, 20) = 3.952, p = 0.061 |
| Thalamic reticular nucleus | −3.60 and −4.08 | C: 5068 (163) N: 5375 (193) | F(1, 20) = 0.974, p = 0.336 |
| Basolateral amygdala | −3.60 and −4.08 | C: 1987 (368) N: 2196 (323) | F(1, 20) = 0.181, p = 0.675 |
| Paraflocculus of the cerebellum | −10.68, −11.16 and −11.64 | C: 2722 (199) N: 2905 (133) | F(1, 20) = 0.658, p = 0.427 |
| Subthalamic nucleus | −3.60 and −4.08 | C: 3114 (249) N: 3141 (125) | F(1, 20) = 0.012, p = 0.915 |
| Locus coeruleus | −9.60 and −10.08 | C: 4899 (279) N: 4716 (222) | F(1, 20) = 0.230, p = 0.637 |
| Lateral habenula | −3.60 and −4.08 | C: 3688 (239) N: 4328 (255) | F(1, 20) = 2.321, p = 0.143 |
| Substantia nigra | −5.52, −6.00 and −6.48 | C: 5439 (114) N: 5450 (152) | F(1, 20) = 0.002, p = 0.962 |
| Dorsal-CA3 | −5.04 and −6.00 | C: 4665 (320) N: 5233 (190) | F(1, 20) = 2.489, p = 0.130 |
| Ventral-CA3 | −5.04 and −6.00 | C: 4042 (188) N: 4519 (171) | F(1, 20) = 2.911, p = 0.103 |
| Dorsal-CA1 | −5.04 and −6.00 | C: 7578 (389) N: 8517 (234) | F(1, 20) = 5.629, p = 0.028 |
| Ventral-CA1 | −5.04 and −6.00 | C: 6521 (502) N: 7298 (284) | F(1, 20) = 2.872, p = 0.106 |
| Primary somatosensory cortex | −4.08 and −4.56 | C: 3436 (471) N: 3991 (158) | F(1, 20) = 1.790, p = 0.196 |
| Dorsal raphe nucleus | −7.20, −7.68 and −8.16 | C: 3793 (762) N: 2995 (351) | F(1, 20) = 1.204, p = 0.301 |
| Posterior commissure | −4.56 and −5.04 | C: 7808 (52) N: 7869 (110) | F(1, 20) = 0.142, p = 0.714 |
| Region | Bregma | Fos-Positive Cells | Statistical Outcome |
|---|---|---|---|
| Primary auditory cortex | −3.60, −4.08, −4.56 and −5.04 | C: 220 (30) N: 316 (25) | F(1, 16) = 8.897, p = 0.009 |
| Medial part of medial geniculate body | −5.04, −5.52, −6.00 and −6.48 | C: 455 (29) N: 520 (30) | F(1, 16) = 0.261, p = 0.61 |
| Central inferior colliculus | −8.04, −8.52 and −9.00 | C: 632 (49) N: 579 (47) | F(1, 16) = 0.088, p = 0.77 |
| Dorsal cochlear nucleus | −10.68, −11.16 and −11.64 | C: 344 (19) N: 362 (23) | F(1, 16) = 1.075, p = 0.31 |
| Region | Bregma | Fos-Positive Cells | Statistical Outcome |
|---|---|---|---|
| Primary auditory cortex | −3.72, −4.08 and −4.44 | ON: 189 (11) OFF: 187 (13) | F (1, 18) = 0.010, p = 0.92 |
| Central inferior colliculus | −8.04, −8.40 and −8.88 | ON: 141 (9) OFF: 121 (17) | F (1, 18) = 2.414, p = 0.138 |
| CA1 of hippocampus | −3.36, −3.72 and −4.08 | ON: 570 (58) OFF: 668 (124) | F (1, 18) = 0.521, p = 0.48 |
| Dentate gyrus | −3.36, −3.72 and −4.08 | ON: 206 (20) OFF: 230 (36) | F (1, 18) = 0.338, p = 0.568 |
| Basolateral amygdala | −2.64, −3.00 and −3.36 | ON: 133 (9) OFF: 116 (10) | F (1, 18) = 1.601, p = 0.222 |
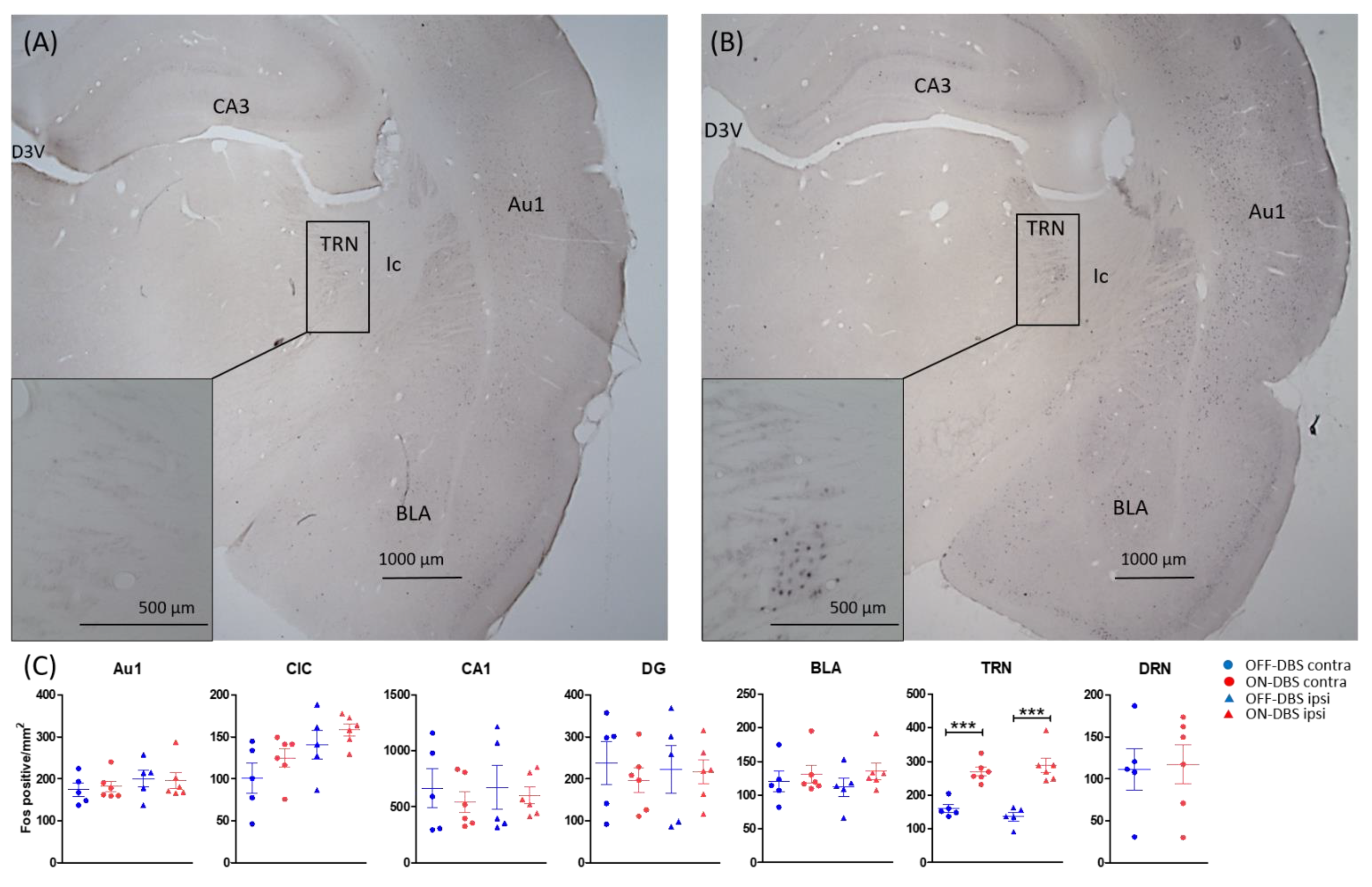
| Thalamic reticular nucleus | −3.00, −3.36 and −3.72 | ON: 279 (15) OFF: 148 (11) | F (1, 18) = 65.379, p < 0.001 |
| Dorsal raphe nucleus | −7.32, −7.68 and −8.04 | ON: 117 (28) OFF: 111 (32) | F (1, 9) = 0.029, p = 0.869 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almasabi, F.; van Zwieten, G.; Alosaimi, F.; Smit, J.V.; Temel, Y.; Janssen, M.L.F.; Jahanshahi, A. The Effect of Noise Trauma and Deep Brain Stimulation of the Medial Geniculate Body on Tissue Activity in the Auditory Pathway. Brain Sci. 2022, 12, 1099. https://doi.org/10.3390/brainsci12081099
Almasabi F, van Zwieten G, Alosaimi F, Smit JV, Temel Y, Janssen MLF, Jahanshahi A. The Effect of Noise Trauma and Deep Brain Stimulation of the Medial Geniculate Body on Tissue Activity in the Auditory Pathway. Brain Sciences. 2022; 12(8):1099. https://doi.org/10.3390/brainsci12081099
Chicago/Turabian StyleAlmasabi, Faris, Gusta van Zwieten, Faisal Alosaimi, Jasper V. Smit, Yasin Temel, Marcus L. F. Janssen, and Ali Jahanshahi. 2022. "The Effect of Noise Trauma and Deep Brain Stimulation of the Medial Geniculate Body on Tissue Activity in the Auditory Pathway" Brain Sciences 12, no. 8: 1099. https://doi.org/10.3390/brainsci12081099
APA StyleAlmasabi, F., van Zwieten, G., Alosaimi, F., Smit, J. V., Temel, Y., Janssen, M. L. F., & Jahanshahi, A. (2022). The Effect of Noise Trauma and Deep Brain Stimulation of the Medial Geniculate Body on Tissue Activity in the Auditory Pathway. Brain Sciences, 12(8), 1099. https://doi.org/10.3390/brainsci12081099