


Daytime-Restricted Feeding Ameliorates Oxidative Stress by Increasing NRF2 Transcriptional Factor in the Rat Hippocampus in the Pilocarpine-Induced Acute Seizure Model

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Daytime-Restricted Feeding and the Pilocarpine-Induced Seizure Model
2.3. Determination of Lipid Peroxidation
2.4. Determination of Reactive Oxygen Species in Hippocampi
2.5. Western Blotting
2.6. Immunofluorescence and Immunohistochemistry
2.7. Confocal Microscopy Analysis
2.8. Statistical Analysis
3. Results
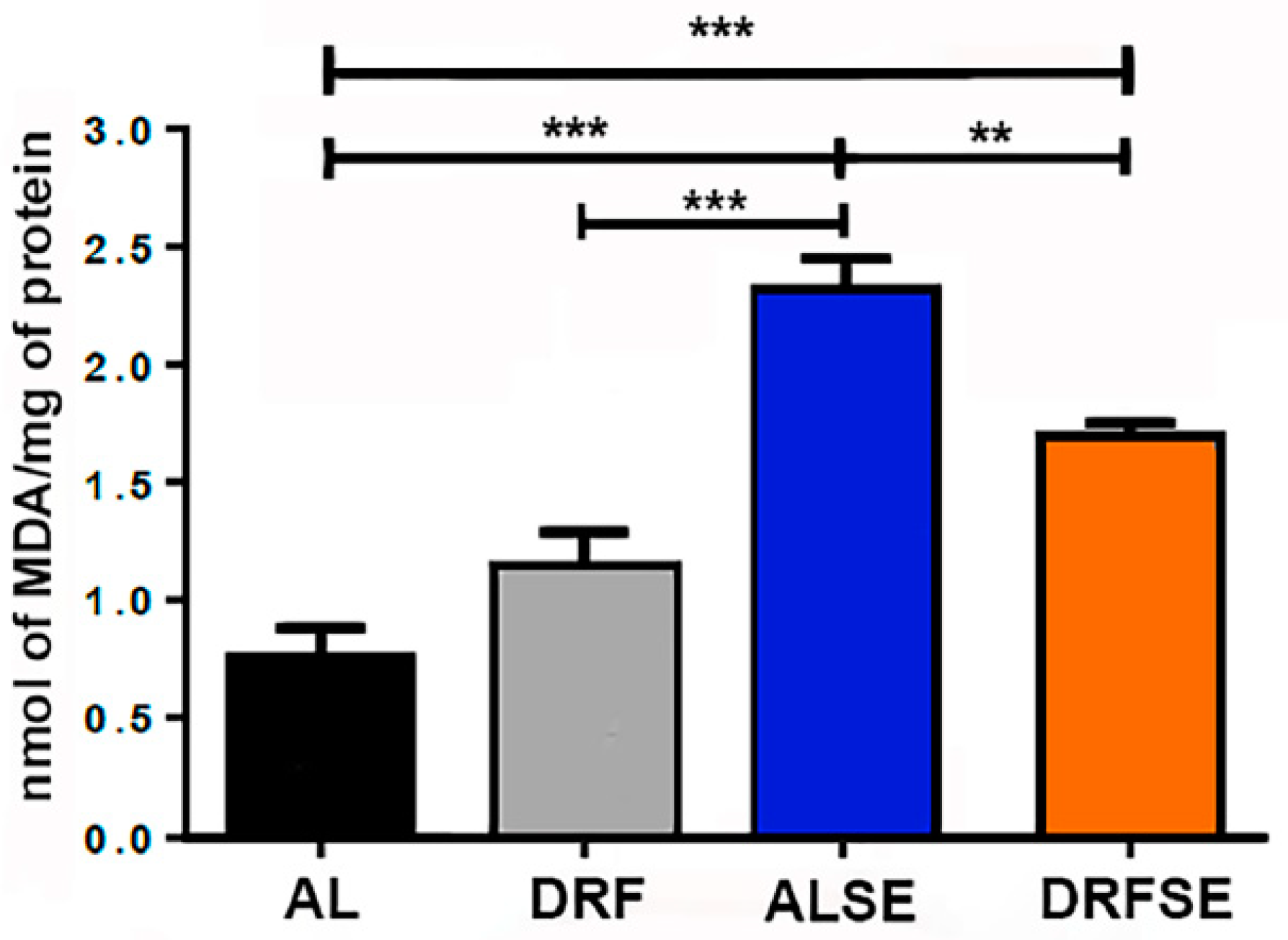
3.1. Dietary Restriction Reduces the Levels of Malondialdehyde in Seizure-Induced Animals
3.2. Daytime-Restricted Feeding Reduces the Levels of Reactive Oxygen Species in Seizure-Induced Animals in CA1 and CA3 Hippocampal Regions
3.3. Daytime-Restricted Feeding Modulates the Content of the Nrf2 Transcriptional Factor in Hippocampal Homogenates and Increases the Nrf2 Immunostaining in CA1 and CA3 Pyramidal Neurons in the SE Model
3.4. Daytime-Restricted Feeding Increases the Content of Superoxide Dismutase 2 in Hippocampal Homogenates and the Immunostaining in CA1 and CA3 Pyramidal Neurons after the Acute Seizure Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thijs, R.D.; Surges, R.; O’Brien, T.J.; Sander, J.W. Epilepsy in adults. Lancet 2019, 393, 689–710. [Google Scholar] [CrossRef]
- Manford, M. Recent advances in epilepsy. J. Neurol. 2017, 264, 1811–1824. [Google Scholar] [CrossRef] [PubMed]
- Trinka, E.; Höfler, J.; Leitinger, M.; Brigo, F. Pharmacotherapy for Status Epilepticus. Drugs 2015, 75, 1499–1521. [Google Scholar] [CrossRef]
- Shin, E.J.; Jeong, J.H.; Chung, Y.H.; Kim, W.K.; Ko, K.H.; Bach, J.H.; Hong, J.S.; Yoneda, Y.; Kim, H.C. Role of oxidative stress in epileptic seizures. Neurochem. Int. 2011, 59, 122–137. [Google Scholar] [CrossRef] [PubMed]
- Pearson-Smith, J.N.; Patel, M. Metabolic Dysfunction and Oxidative Stress in Epilepsy. Int. J. Mol. Sci. 2017, 18, 2365. [Google Scholar] [CrossRef] [PubMed]
- Shekh-Ahmad, T.; Kovac, S.; Abramov, A.Y.; Walker, M.C. Reactive oxygen species in status epilepticus. Epilepsy Behav. 2019, 101, 106410. [Google Scholar] [CrossRef]
- Huang, W.Y.; Lin, S.; Chen, H.Y.; Chen, Y.P.; Chen, T.Y.; Hsu, K.S.; Wu, H.M. NADPH oxidases as potential pharmacological targets against increased seizure susceptibility after systemic inflammation. J. Neuroinflamm. 2018, 15, 140. [Google Scholar] [CrossRef]
- Chuang, Y.C. Mitochondrial dysfunction and oxidative stress in seizure-induced neuronal cell death. Acta Neurol. Taiwan 2010, 19, 3–15. [Google Scholar]
- Yang, N.; Guan, Q.W.; Chen, F.H.; Xia, Q.X.; Yin, X.X.; Zhou, H.H.; Mao, X.Y. Antioxidants targeting mitochondrial oxidative stress: Promising neuroprotectants for epilepsy. Oxid. Med. Cell Longev. 2020, 2020, 6687185. [Google Scholar] [CrossRef]
- Kasai, S.; Shimizu, S.; Tatara, Y.; Mimura, J.; Itch, K. Regulation of Nrf2 by mitochondrial reactive oxygen species in physiology and pathology. Biomolecules 2020, 10, 320. [Google Scholar] [CrossRef]
- Zhao, J.; Kobori, N.; Aronowski, J.; Dash, P.K. Sulforaphane reduces infarct volume following focal cerebral ischemia in rodents. Neurosci. Lett. 2006, 393, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Hartman, A.L. Neuroprotection in metabolism-based therapy. Epilepsy Res. 2012, 100, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Landgrave-Gómez, J.; Mercado-Gómez, O.F.; Vázquez-García, M.; Rodríguez-Molina, V.; Córdova-Dávalos, L.; Arriaga-Ávila, V.; Miranda-Martínez, A.; Guevara-Guzmán, R. Anticonvulsant effect of time-restricted feeding in a pilocarpine-induced seizure model: Metabolic and epigenetic implications. Front. Cell Neurosci. 2016, 10, 7. [Google Scholar] [CrossRef] [PubMed]
- Santillán-Cigales, J.J.; Mercado-Gómez, O.F.; Arriaga-Ávila, V.; Landgrave-Gómez, J.; Guevara-Guzmán, R. Daytime-restricted feeding modulates the expression of inflammatory mediators and diminishes reactive astrogliosis and microgliosis following status epilepticus. Brain Res. 2020, 1734, 146724. [Google Scholar] [CrossRef]
- Racine, R.J.; Gartner, J.G.; Burnham, W.M. Epileptiform activity and neural plasticity in limbic structures. Brain Res. 1972, 47, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Scharfman, H.E. The dentate gyrus and temporal lobe epilepsy: An “exciting” era. Epilepsy Curr. 2019, 19, 249–255. [Google Scholar] [CrossRef]
- Kim, J.E.; Kang, T.C. CDDO-Me attenuates astroglial autophagy via Nrf2-, ERK1/2-SP1- and Src-CK2-PTEN-PI3K/AKT-mediated signaling pathways in the hippocampus of chronic epilepsy rats. Antioxidants 2021, 10, 655. [Google Scholar] [CrossRef]
- Halliwell, B. Reactive oxygen species and the central nervous system. J. Neurochem. 1992, 59, 1609–1623. [Google Scholar] [CrossRef]
- da Fonsêca, D.V.; da Silva Maia Bezerra Filho, C.; Lima, T.C.; de Almeida, R.N.; de Sousa, D.P. Anticonvulsant essential oils and their relationship with oxidative stress in epilepsy. Biomolecules 2019, 9, 835. [Google Scholar] [CrossRef]
- Ziegler, D.R.; Ribeiro, L.C.; Hagenn, M.; Siqueira, I.R.; Araújo, E.; Torres, I.L.; Gottfried, C.; Netto, C.A.; Gonçalves, C.A. Ketogenic diet increases glutathione peroxidase activity in rat hippocampus. Neurochem. Res. 2003, 28, 1793–1797. [Google Scholar] [CrossRef]
- Maalouf, M.; Sullivan, P.G.; Davis, L.; Kim, D.Y.; Rho, J.M. Ketones inhibit mitochondrial production of reactive oxygen species production following glutamate excitotoxicity by increasing NADH oxidation. Neuroscience 2007, 145, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Mazzuferi, M.; Kumar, G.; van Eyll, J.; Danis, B.; Foerch, P.; Kaminski, R.M. Nrf2 defense pathway: Experimental evidence for its protective role in epilepsy. Ann. Neurol. 2013, 74, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, W.P.; Zhang, G.L.; Wu, Y.F.; Xie, T.; Kan, M.C.; Fang, H.B.; Wang, H.C. Activation of Nrf2-ARE signal pathway in hippocampus of amygdala kindling rats. Neurosci. Lett. 2013, 543, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Alvi, A.M.; Al Kury, L.T.; Alattar, A.; Ullah, I.; Muhammad, A.J.; Alshaman, R.; Shah, F.A.; Khan, A.U.; Feng, J.; Li, S. Carveol Attenuates seizure severity and neuroinflammation in pentylenetetrazole-kindled epileptic rats by regulating the Nrf2 signaling pathway. Oxid. Med. Cell Longev. 2021, 2021, 9966663. [Google Scholar] [CrossRef]
- Wang, W.; Wu, Y.; Zhang, G.; Fang, H.; Wang, H.; Zang, H.; Xie, T.; Wang, W. Activation of Nrf2-ARE signal pathway protects the brain from damage induced by epileptic seizure. Brain Res. 2014, 1544, 54–61. [Google Scholar] [CrossRef]
- Liu, Z.; Yang, C.; Meng, X.; Li, Z.; Lv, C.; Cao, P. Neuroprotection of edaravone on the hippocampus of kainate-induced epilepsy rats through Nrf2/HO-1 pathway. Neurochem. Int. 2018, 112, 159–165. [Google Scholar] [CrossRef]
- Singh, N.; Vijayanti, S.; Saha, L.; Bhatia, A.; Banerjee, D.; Chakrabarti, A. Neuroprotective effect of Nrf2 activator dimethyl fumarate, on the hippocampal neurons in chemical kindling model in rat. Epilepsy Res. 2018, 143, 98–104. [Google Scholar] [CrossRef]
- Munguía-Martínez, M.F.; Nava-Ruíz, C.; Ruíz-Díaz, A.; Díaz-Ruíz, A.; Yescas-Gómez, P.; Méndez-Armenta, M. Immunohistochemical study of antioxidant enzymes regulated by Nrf2 in the models of epileptic seizures (KA and PTZ). Oxid. Med. Cell Longev. 2019, 2019, 1327986. [Google Scholar] [CrossRef]
- Coulter, D.A.; Steinhäuser, C. Role of astrocytes in epilepsy. Cold Spring Harb. Perspect. Med. 2015, 5, a022434. [Google Scholar] [CrossRef]
- Binder, D.K.; Steinhäuser, C. Astrocytes and Epilepsy. Neurochem. Res. 2021, 46, 2687–2695. [Google Scholar] [CrossRef]
- Kim, D.S.; Kim, J.E.; Kwak, S.E.; Choi, K.C.; Kim, D.W.; Kwon, O.S.; Choi, S.Y.; Kang, T.C. Spatiotemporal characteristics of astroglial death in the rat hippocampo-entorhinal complex following pilocarpine-induced status epilepticus. J. Comp. Neurol. 2008, 511, 581–598. [Google Scholar] [CrossRef]
- Rowley, S.; Patel, M. Mitochondrial involvement and oxidative stress in temporal lobe epilepsy. Free Radic. Biol. Med. 2013, 62, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Gluck, M.R.; Jayatilleke, E.; Shaw, S.; Rowan, A.J.; Haroutunian, V. CNS oxidative stress associated with the kainic acid rodent model of experimental epilepsy. Epilepsy Res. 2000, 39, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Frantseva, M.V.; Perez-Velazquez, J.L.; Tsoraklidis, G.; Mendonca, A.J.; Adamchik, Y.; Mills, L.R.; Carlen, P.L.; Burnham, M.W. Oxidative stress is involved in seizure-induced neurodegeneration in the kindling model of epilepsy. Neuroscience 2000, 97, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Palma, F.R.; He, C.; Danes, J.M.; Paviani, V.; Coelho, D.R.; Gantner, B.N.; Bonini, M.G. Mitochondrial superoxide dismutase: What the established, the intriguing, and the novel reveal about a key cellular redox switch. Antioxid. Redox. Signal 2020, 32, 701–714. [Google Scholar] [CrossRef]
- Liang, L.P.; Waldbaum, S.; Rowley, S.; Huang, T.T.; Day, B.J.; Patel, M. Mitochondrial oxidative stress and epilepsy in SOD2 deficient mice: Attenuation by a lipophilic metalloporphyrin. Neurobiol. Dis. 2012, 45, 1068–1076. [Google Scholar] [CrossRef]
- Fulton, R.E.; Pearson-Smith, J.N.; Huynh, C.Q.; Fabisiak, T.; Liang, L.P.; Aivazidis, S.; High, B.A.; Buscaglia, G.; Corrigan, T.; Valdez, R.; et al. Neuron-specific mitochondrial oxidative stress results in epilepsy, glucose dysregulation and a striking astrocyte response. Neurobiol. Dis. 2021, 158, 105470. [Google Scholar] [CrossRef]
- Shimazu, T.; Hirschey, M.D.; Newman, J.; He, W.; Shirakawa, K.; Le Moan, N.; Grueter, C.A.; Lim, H.; Saunders, L.R.; Stevens, R.D.; et al. Suppression of oxidative stress by β-hydroxybutyrate, an endogenous histone deacetylase inhibitor. Science 2013, 339, 211–214. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mercado-Gómez, O.F.; Arriaga-Ávila, V.S.; Vega-García, A.; Orozco-Suarez, S.; Pérez-Koldenkova, V.; Camarillo-Sánchez, J.J.; Álvarez-Herrera, M.; Guevara-Guzmán, R. Daytime-Restricted Feeding Ameliorates Oxidative Stress by Increasing NRF2 Transcriptional Factor in the Rat Hippocampus in the Pilocarpine-Induced Acute Seizure Model. Brain Sci. 2023, 13, 1442. https://doi.org/10.3390/brainsci13101442
Mercado-Gómez OF, Arriaga-Ávila VS, Vega-García A, Orozco-Suarez S, Pérez-Koldenkova V, Camarillo-Sánchez JJ, Álvarez-Herrera M, Guevara-Guzmán R. Daytime-Restricted Feeding Ameliorates Oxidative Stress by Increasing NRF2 Transcriptional Factor in the Rat Hippocampus in the Pilocarpine-Induced Acute Seizure Model. Brain Sciences. 2023; 13(10):1442. https://doi.org/10.3390/brainsci13101442
Chicago/Turabian StyleMercado-Gómez, Octavio Fabián, Virginia Selene Arriaga-Ávila, Angélica Vega-García, Sandra Orozco-Suarez, Vadim Pérez-Koldenkova, Juan José Camarillo-Sánchez, Marcelino Álvarez-Herrera, and Rosalinda Guevara-Guzmán. 2023. "Daytime-Restricted Feeding Ameliorates Oxidative Stress by Increasing NRF2 Transcriptional Factor in the Rat Hippocampus in the Pilocarpine-Induced Acute Seizure Model" Brain Sciences 13, no. 10: 1442. https://doi.org/10.3390/brainsci13101442
APA StyleMercado-Gómez, O. F., Arriaga-Ávila, V. S., Vega-García, A., Orozco-Suarez, S., Pérez-Koldenkova, V., Camarillo-Sánchez, J. J., Álvarez-Herrera, M., & Guevara-Guzmán, R. (2023). Daytime-Restricted Feeding Ameliorates Oxidative Stress by Increasing NRF2 Transcriptional Factor in the Rat Hippocampus in the Pilocarpine-Induced Acute Seizure Model. Brain Sciences, 13(10), 1442. https://doi.org/10.3390/brainsci13101442