
Atypical Associations between Functional Connectivity during Pragmatic and Semantic Language Processing and Cognitive Abilities in Children with Autism
 ,
, 
Abstract
:1. Introduction
2. Methods
2.1. Participants
2.2. Behavioral Assessment
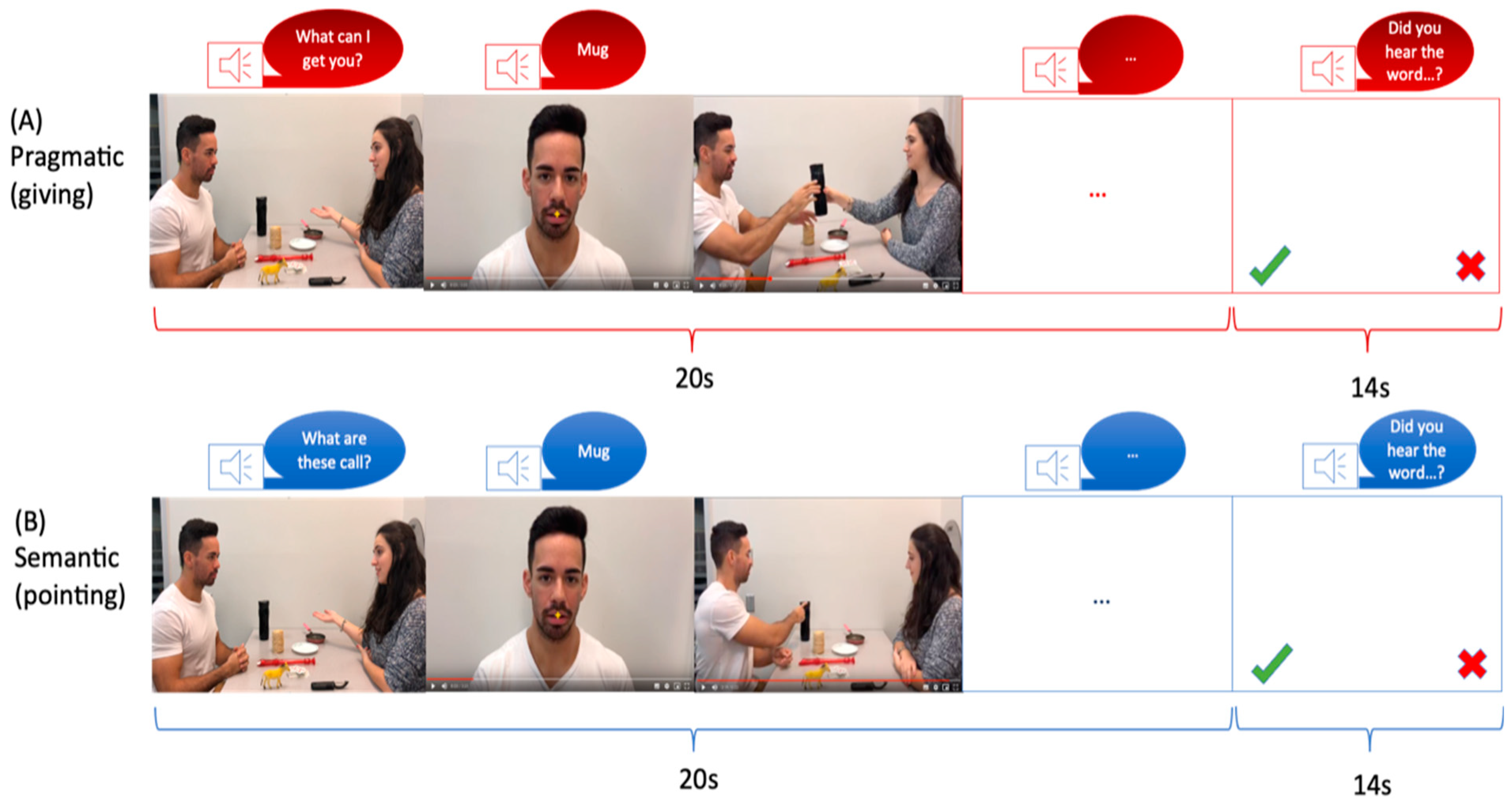
2.3. Task Protocol
2.4. MRI Data Acquisition
2.5. fMRI Pre-Processing
2.6. Estimation of Functional Connectivity during Pragmatic and Semantic Processing
2.7. Associations between Brain Connectivity and Psychometrics
2.8. Multivariate Framework for Analysis of Correlations
3. Results
3.1. Participants’ Psychometric Profiles
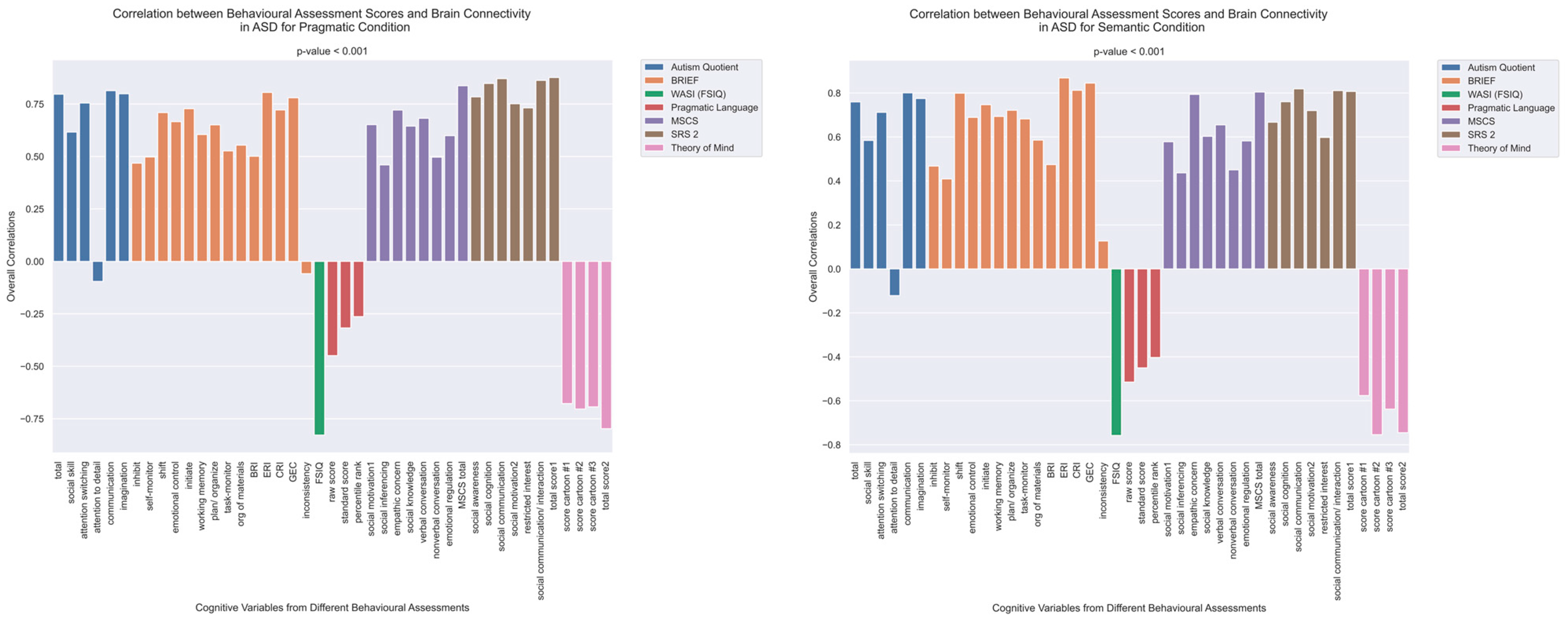
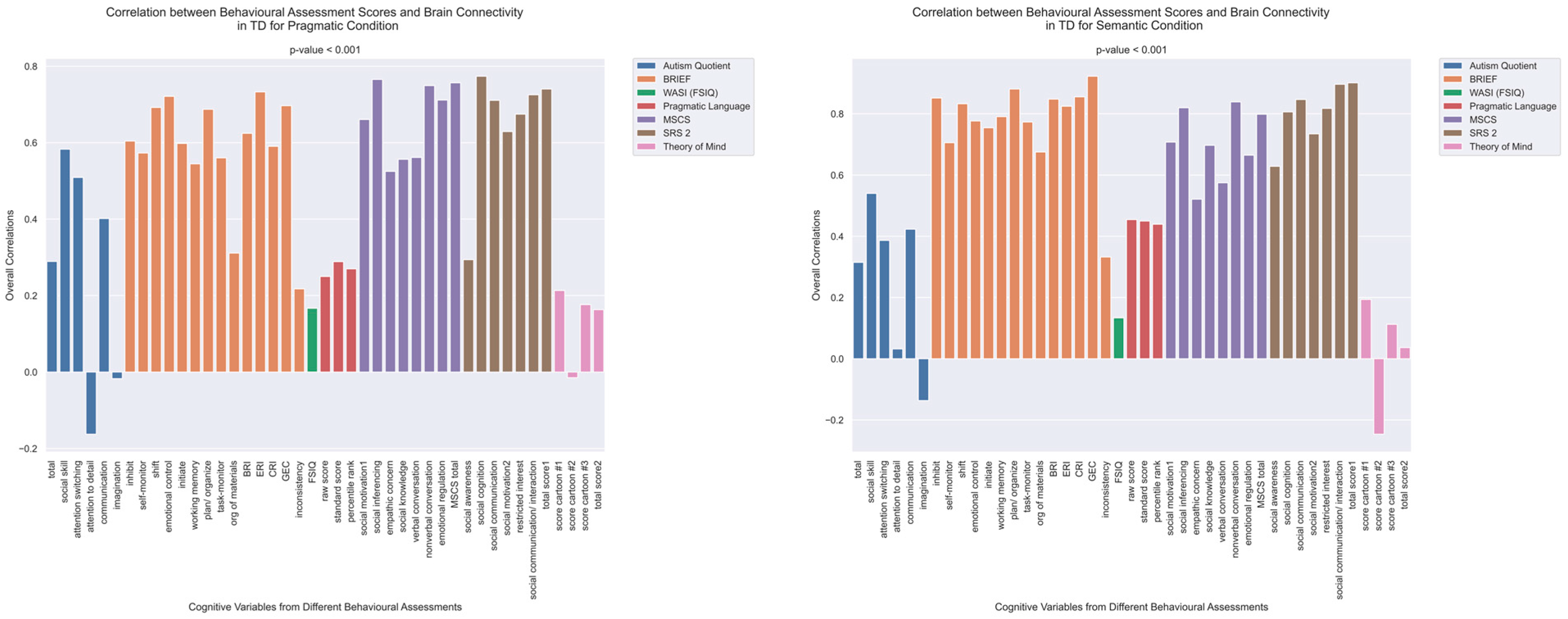
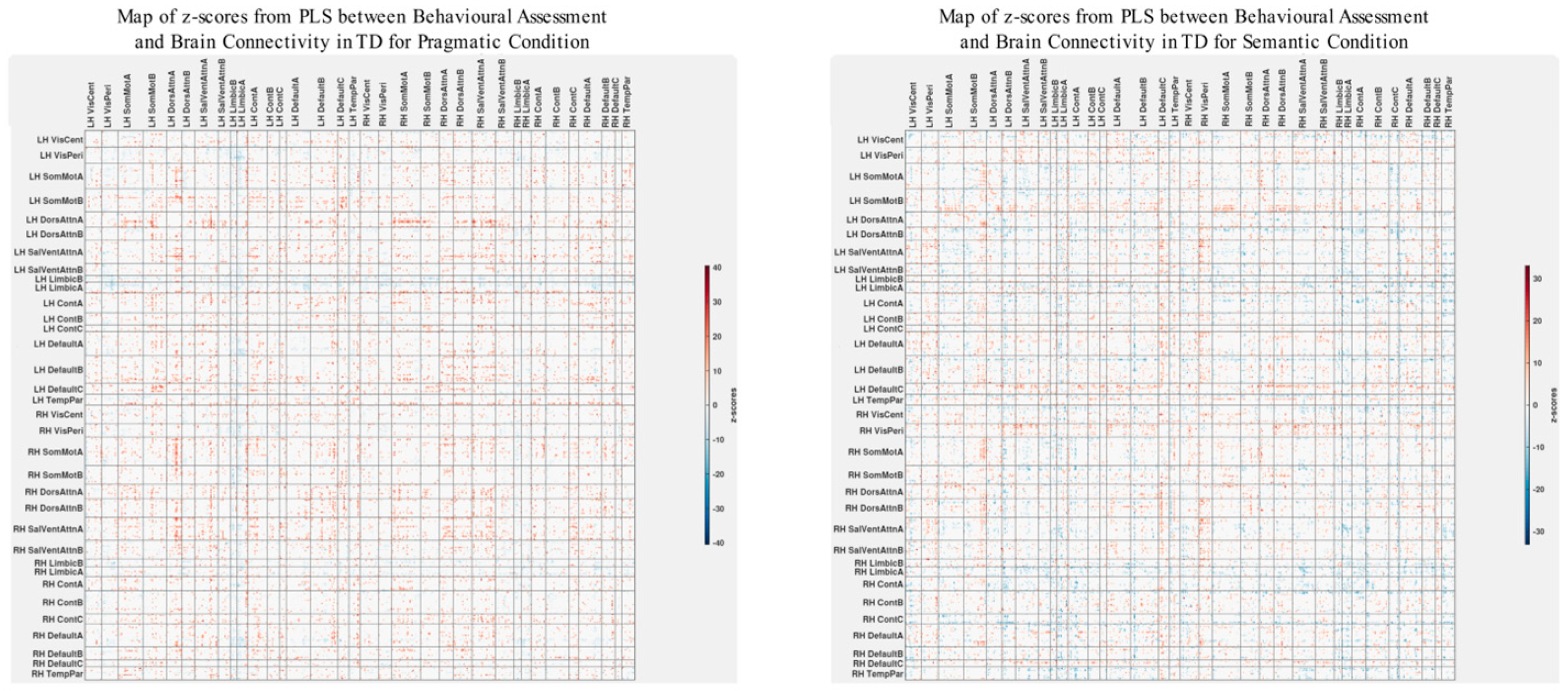
3.2. Cognitive Performance Associated with Functional Connectivity in TD and ASD Groups
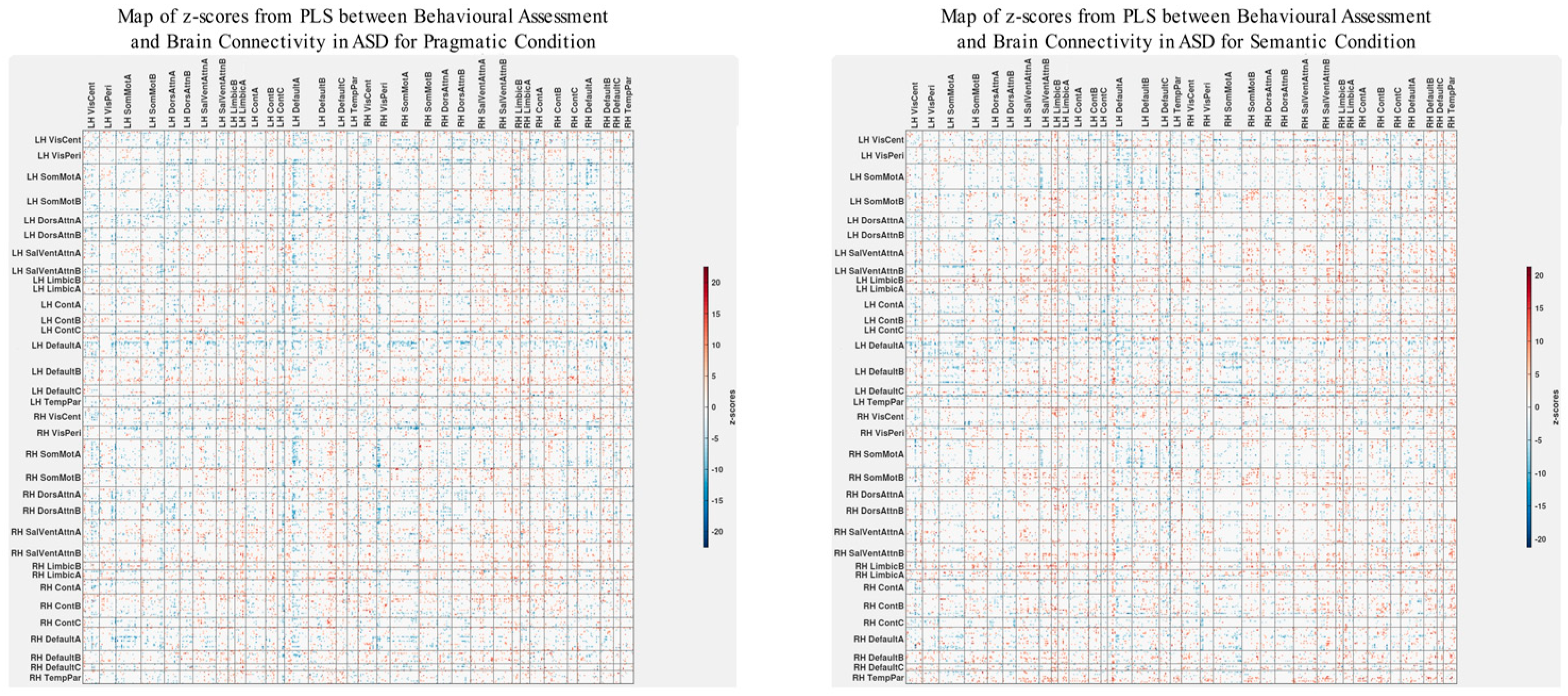
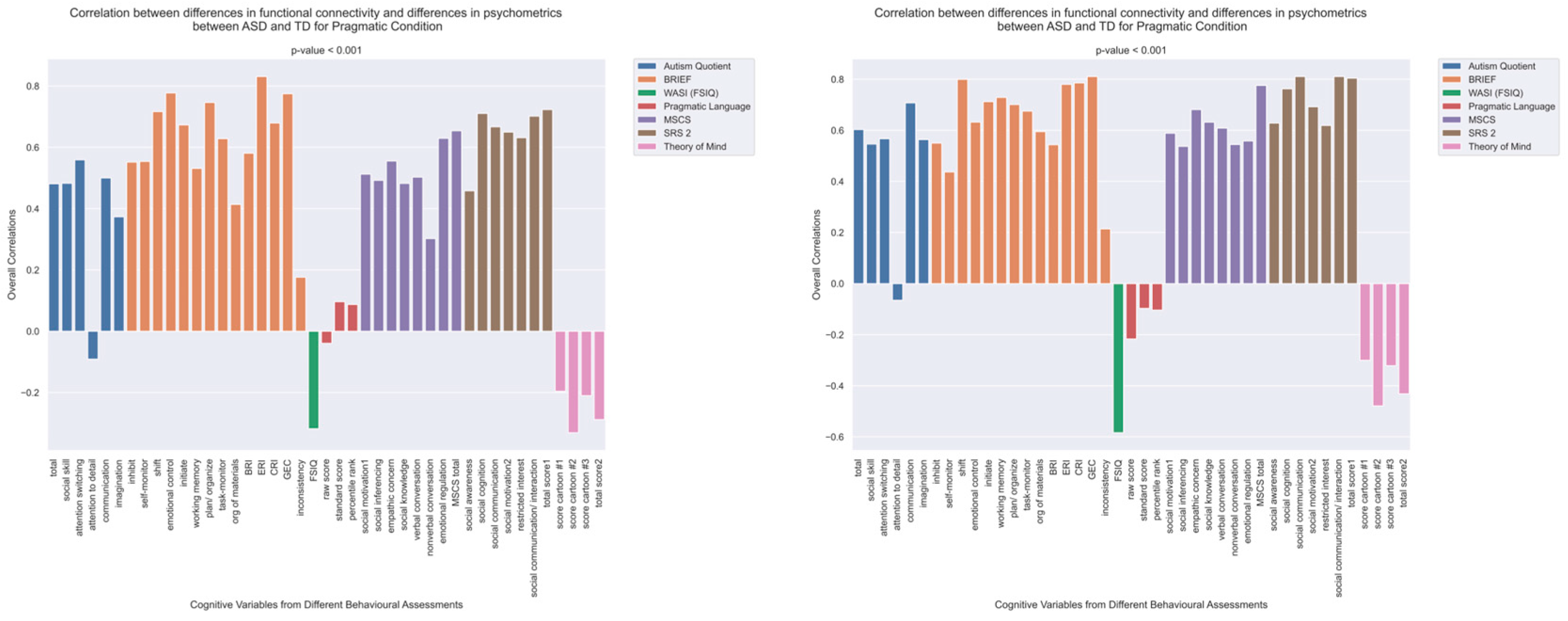
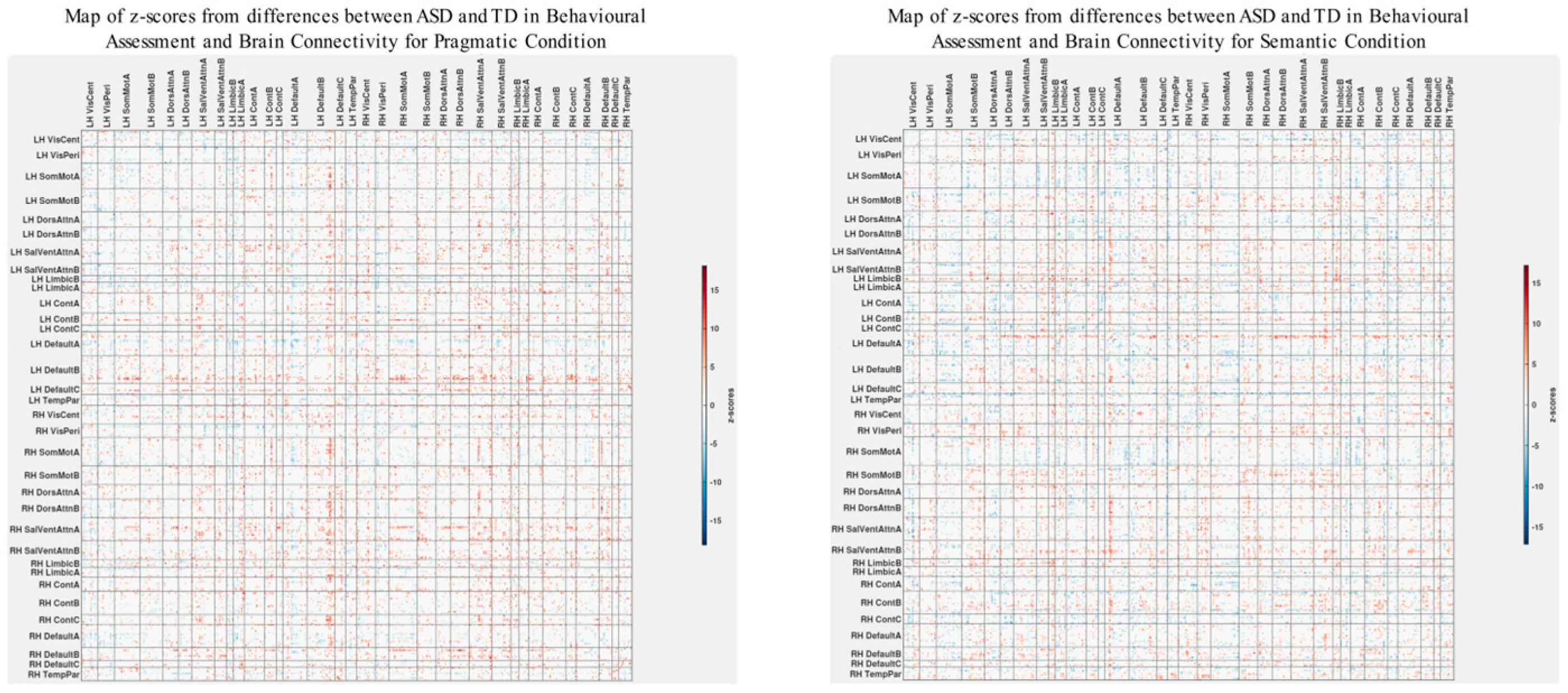
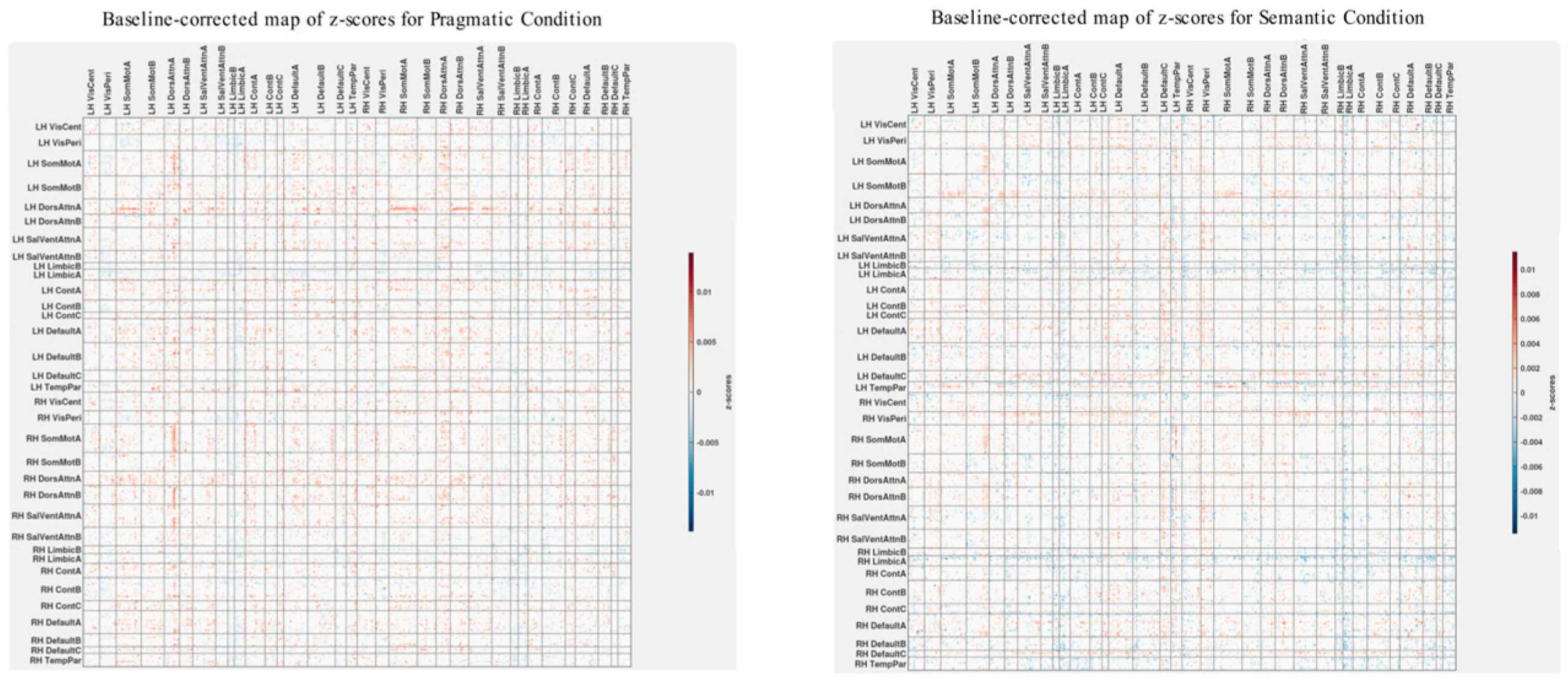
3.3. Differences in Correlations between Group Differences in Bran Connectivity and Group Differences in Psychometrics
4. Discussion
5. Limitations
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maenner, M.J.; Warren, Z.; Williams, A.R.; Amoakohene, E.; Bakian, A.V.; Bilder, D.A.; Durkin, M.S.; Fitzgerald, R.T.; Furnier, S.M.; Hughes, M.M.; et al. Prevalence and Characteristics of Autism Spectrum Disorder Among Children Aged 8 Years—Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2020. MMWR Surveill. Summ. 2023, 72, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kopanska, M.; Ochojska, D.; Banas-Zabczyk, A.; Sochocka, M.; Ochnik, M.; Torices, S.; Morgan, R.C.; Leszek, J.; Podgorska-Bednarz, J.; Szczygielski, J. Brain as the House of Autistic Soul: Current Trends in Analyzing the Risk Factors and Building up the Diagnosis in Mild Forms of Autism Spectrum Disorder. J. Physiol. Pharmacol. 2022, 72, 6. [Google Scholar] [CrossRef]
- Lord, C.; Luyster, R. Early Diagnosis of Children with Autism Spectrum Disorders. Clin. Neurosci. Res. 2006, 6, 189–194. [Google Scholar] [CrossRef]
- Herringshaw, A.J.; Ammons, C.J.; DeRamus, T.P.; Kana, R.K. Hemispheric Differences in Language Processing in Autism Spectrum Disorders: A Meta-Analysis of Neuroimaging Studies: Neural Correlates of Language in Autism. Autism Res. 2016, 9, 1046–1057. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 4th ed.; American Psychiatric Association: Washington, DC, USA, 2013; ISBN 978-0-89042-555-8. [Google Scholar]
- Vulchanova, M.; Saldaña, D.; Chahboun, S.; Vulchanov, V. Figurative Language Processing in Atypical Populations: The ASD Perspective. Front. Hum. Neurosci. 2015, 9, 24. [Google Scholar] [CrossRef]
- Corbett, B.A.; Newsom, C.; Key, A.P.; Qualls, L.R.; Edmiston, E.K. Examining the Relationship between Face Processing and Social Interaction Behavior in Children with and without Autism Spectrum Disorder. J. Neurodev. Disord. 2014, 6, 35. [Google Scholar] [CrossRef]
- Hull, J.V.; Dokovna, L.B.; Jacokes, Z.J.; Torgerson, C.M.; Irimia, A.; Van Horn, J.D. Resting-State Functional Connectivity in Autism Spectrum Disorders: A Review. Front. Psychiatry 2017, 7, 205. [Google Scholar] [CrossRef]
- You, X.; Norr, M.; Murphy, E.; Kuschner, E.S.; Bal, E.; Gaillard, W.D.; Kenworthy, L.; Vaidya, C.J. Atypical Modulation of Distant Functional Connectivity by Cognitive State in Children with Autism Spectrum Disorders. Front. Hum. Neurosci. 2013, 7, 482. [Google Scholar] [CrossRef]
- Marta, K.; Justyna, C.-Ł.; Marta, S.; Jerzy, L.; Justyna, P.-B.; Agnieszka, B.-Z.; Danuta, O. Selected Methods of Therapeutic Interactions With People With Mild Symptoms of Autism Spectrum Disorder. Front. Psychiatry 2022, 13, 942218. [Google Scholar] [CrossRef]
- Rane, P.; Cochran, D.; Hodge, S.M.; Haselgrove, C.; Kennedy, D.N.; Frazier, J.A. Connectivity in Autism: A Review of MRI Connectivity Studies. Harv. Rev. Psychiatry 2015, 23, 223–244. [Google Scholar] [CrossRef]
- Kana, R.K.; Uddin, L.Q.; Kenet, T.; Chugani, D.; Müller, R.-A. Brain Connectivity in Autism. Front. Hum. Neurosci. 2014, 8, 349. [Google Scholar] [CrossRef] [PubMed]
- Fetit, R.; Hillary, R.F.; Price, D.J.; Lawrie, S.M. The Neuropathology of Autism: A Systematic Review of Post-Mortem Studies of Autism and Related Disorders. Neurosci. Biobehav. Rev. 2021, 129, 35–62. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.M.Y.; Yau, S.-Y.; Chan, M.M.Y.; Wong, C.-K.; Chan, A.S. Altered Cytokine and BDNF Levels in Individuals with Autism Spectrum Disorders. Brain Sci. 2022, 12, 460. [Google Scholar] [CrossRef] [PubMed]
- Babaeeghazvini, P.; Rueda-Delgado, L.M.; Gooijers, J.; Swinnen, S.P.; Daffertshofer, A. Brain Structural and Functional Connectivity: A Review of Combined Works of Diffusion Magnetic Resonance Imaging and Electro-Encephalography. Front. Hum. Neurosci. 2021, 15, 721206. [Google Scholar] [CrossRef]
- Ellenbroek, B.; Youn, J. Gene-Environment Interactions in Psychiatry: Nature, Nurture, Neuroscience; ScienceDirect; Academic Press: London, UK, 2016; ISBN 978-0-12-801783-8. [Google Scholar]
- Müller, R.-A.; Shih, P.; Keehn, B.; Deyoe, J.R.; Leyden, K.M.; Shukla, D.K. Underconnected, but How? A Survey of Functional Connectivity MRI Studies in Autism Spectrum Disorders. Cerebral. Cortex. 2011, 21, 2233–2243. [Google Scholar] [CrossRef]
- Lindell, A.K.; Hudry, K. Atypicalities in Cortical Structure, Handedness, and Functional Lateralization for Language in Autism Spectrum Disorders. Neuropsychol. Rev. 2013, 23, 257–270. [Google Scholar] [CrossRef]
- Colich, N.L.; Wang, A.-T.; Rudie, J.D.; Hernandez, L.M.; Bookheimer, S.Y.; Dapretto, M. Atypical Neural Processing of Ironic and Sincere Remarks in Children and Adolescents with Autism Spectrum Disorders. Metaphor. Symb. 2012, 27, 70–92. [Google Scholar] [CrossRef]
- Khan, S.; Gramfort, A.; Shetty, N.R.; Kitzbichler, M.G.; Ganesan, S.; Moran, J.M.; Lee, S.M.; Gabrieli, J.D.E.; Tager-Flusberg, H.B.; Joseph, R.M.; et al. Local and Long-Range Functional Connectivity Is Reduced in Concert in Autism Spectrum Disorders. Proc. Natl. Acad. Sci. USA 2013, 110, 3107–3112. [Google Scholar] [CrossRef]
- Galvez-Contreras, A.Y.; Zarate-Lopez, D.; Torres-Chavez, A.L.; Gonzalez-Perez, O. Role of Oligodendrocytes and Myelin in the Pathophysiology of Autism Spectrum Disorder. Brain Sci. 2020, 10, 951. [Google Scholar] [CrossRef]
- Shen, H.-Y.; Huang, N.; Reemmer, J.; Xiao, L. Adenosine Actions on Oligodendroglia and Myelination in Autism Spectrum Disorder. Front. Cell Neurosci. 2018, 12, 482. [Google Scholar] [CrossRef]
- Harris, G.J.; Chabris, C.F.; Clark, J.; Urban, T.; Aharon, I.; Steele, S.; McGrath, L.; Condouris, K.; Tager-Flusberg, H. Brain Activation during Semantic Processing in Autism Spectrum Disorders via Functional Magnetic Resonance Imaging. Brain Cogn. 2006, 61, 54–68. [Google Scholar] [CrossRef] [PubMed]
- Just, M.A.; Cherkassky, V.L.; Buchweitz, A.; Keller, T.A.; Mitchell, T.M. Identifying Autism from Neural Representations of Social Interactions: Neurocognitive Markers of Autism. PLoS ONE 2014, 9, e113879. [Google Scholar] [CrossRef] [PubMed]
- Kana, R.K. Sentence Comprehension in Autism: Thinking in Pictures with Decreased Functional Connectivity. Brain 2006, 129, 2484–2493. [Google Scholar] [CrossRef] [PubMed]
- Knaus, T.A.; Silver, A.M.; Kennedy, M.; Lindgren, K.A.; Dominick, K.C.; Siegel, J.; Tager-Flusberg, H. Language Laterality in Autism Spectrum Disorder and Typical Controls: A Functional, Volumetric, and Diffusion Tensor MRI Study. Brain Lang. 2010, 112, 113–120. [Google Scholar] [CrossRef]
- Lawrence, K.E.; Hernandez, L.M.; Bookheimer, S.Y.; Dapretto, M. Atypical Longitudinal Development of Functional Connectivity in Adolescents with Autism Spectrum Disorder. Autism Res. 2019, 12, 53–65. [Google Scholar] [CrossRef]
- Rohr, C.S.; Kamal, S.; Bray, S. Building Functional Connectivity Neuromarkers of Behavioral Self-Regulation across Children with and without Autism Spectrum Disorder. Dev. Cogn. Neurosci. 2020, 41, 100747. [Google Scholar] [CrossRef]
- Courchesne, E. Evidence of Brain Overgrowth in the First Year of Life in Autism. JAMA 2003, 290, 337. [Google Scholar] [CrossRef]
- Courchesne, E.; Redcay, E.; Kennedy, D.P. The Autistic Brain: Birth through Adulthood. Curr. Opin. Neurol. 2004, 17, 489–496. [Google Scholar] [CrossRef]
- Uddin, L.Q.; Supekar, K.; Lynch, C.J.; Khouzam, A.; Phillips, J.; Feinstein, C.; Ryali, S.; Menon, V. Salience Network–Based Classification and Prediction of Symptom Severity in Children With Autism. JAMA Psychiatry 2013, 70, 869. [Google Scholar] [CrossRef]
- Liakakis, G.; Nickel, J.; Seitz, R.J. Diversity of the Inferior Frontal Gyrus—A Meta-Analysis of Neuroimaging Studies. Behav. Brain Res. 2011, 225, 341–347. [Google Scholar] [CrossRef]
- Uchiyama, H.; Seki, A.; Kageyama, H.; Saito, D.N.; Koeda, T.; Ohno, K.; Sadato, N. Neural Substrates of Sarcasm: A Functional Magnetic-Resonance Imaging Study. Brain Res. 2006, 1124, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Aron, A.R.; Fletcher, P.C.; Bullmore, E.T.; Sahakian, B.J.; Robbins, T.W. Erratum: Stop-Signal Inhibition Disrupted by Damage to Right Inferior Frontal Gyrus in Humans. Nat. Neurosci. 2003, 6, 1329. [Google Scholar] [CrossRef]
- Groen, W.; Zwiers, M.; Vandergaag, R.; Buitelaar, J. The Phenotype and Neural Correlates of Language in Autism: An Integrative Review. Neurosci. Biobehav. Rev. 2008, 32, 1416–1425. [Google Scholar] [CrossRef] [PubMed]
- Mash, L.E.; Reiter, M.A.; Linke, A.C.; Townsend, J.; Müller, R. Multimodal Approaches to Functional Connectivity in Autism Spectrum Disorders: An Integrative Perspective. Dev. Neurobiol. 2018, 78, 456–473. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, A.; Williams, L.J.; McIntosh, A.R.; Abdi, H. Partial Least Squares (PLS) Methods for Neuroimaging: A Tutorial and Review. NeuroImage 2011, 56, 455–475. [Google Scholar] [CrossRef]
- McIntosh, A.R.; Bookstein, F.L.; Haxby, J.V.; Grady, C.L. Spatial Pattern Analysis of Functional Brain Images Using Partial Least Squares. NeuroImage 1996, 3, 143–157. [Google Scholar] [CrossRef]
- Baron-Cohen, S.; Wheelwright, S.; Skinner, R.; Martin, J.; Clubley, E. The Autism-Spectrum Quotient (AQ): Evidence from Asperger Syndrome/High-Functioning Autism, Males and Females, Scientists and Mathematicians. J. Autism Dev. Disord. 2001, 31, 5–17. [Google Scholar] [CrossRef]
- Chandler, M.J. Egocentrism and Antisocial Behavior: The Assessment and Training of Social Perspective-Taking Skills. Dev. Psychol. 1973, 9, 326–332. [Google Scholar] [CrossRef]
- Yager, J.; Iarocci, G. The Development of the Multidimensional Social Competence Scale: A Standardized Measure of Social Competence in Autism Spectrum Disorders: Social Competence in Autism. Autism. Res. 2013, 6, 631–641. [Google Scholar] [CrossRef]
- Gioia, G.A.; Isquith, P.K.; Guys, S.C.; Kenworthy, L.L. Behavior Rating Inventory of Executive Function®, 2nd ed.; (BRIEF®2); PAR Inc.: Lutz, FL, USA, 2015. [Google Scholar]
- Constantino, J.N.; Gruber, C.P. Social Responsiveness Scale (SRS); Western Psychological Services: Los Angeles, CA, USA, 2011. [Google Scholar]
- Carrow-Woolfolk, E. Comprehensive Assessment of Spoken Language, 2nd ed.; (CASL-2) [Manual]; WPS: Torrance, CA, USA, 2017. [Google Scholar]
- Wechsler, D. Wechsler Abbreviated Scale of Intelligence: WASI-II, 2nd ed.; Manual; Pearson: Bloomington, MI, USA, 2011; ISBN 978-0-15-898158-1. [Google Scholar]
- Reynolds, C.R.; Kamphaus, R.W. Behavior Assessment System for Children; Ags Publishing: Circle Pines, MI, USA, 2004. [Google Scholar]
- Márquez-Garcia, A.V.; Ng, B.; Vakorin, V.A.; Kozhemiako, N.; Iarocci, G.; Moreno, S.; Doesburg, S.M. Functional brain connectivity alterations during sematic and pragmatic language processing in children with autism. Sci. Rep. 2023; submitted. [Google Scholar]
- Egorova, N.; Shtyrov, Y.; Pulvermüller, F. Early and Parallel Processing of Pragmatic and Semantic Information in Speech Acts: Neurophysiological Evidence. Front. Hum. Neurosci. 2013, 7, 86. [Google Scholar] [CrossRef]
- Egorova, N.; Shtyrov, Y.; Pulvermüller, F. Brain Basis of Communicative Actions in Language. NeuroImage 2016, 125, 857–867. [Google Scholar] [CrossRef] [PubMed]
- Dale, A.M.; Fischl, B.; Sereno, M.I. Cortical Surface-Based Analysis. NeuroImage 1999, 9, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Dale, A.M.; Sereno, M.I. Improved Localizadon of Cortical Activity by Combining EEG and MEG with MRI Cortical Surface Reconstruction: A Linear Approach. J. Cogn. Neurosci. 1993, 5, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Fischl, B.; Dale, A.M. Measuring the Thickness of the Human Cerebral Cortex from Magnetic Resonance Images. Proc. Natl. Acad. Sci. USA 2000, 97, 11050–11055. [Google Scholar] [CrossRef]
- Fischl, B.; Liu, A.; Dale, A.M. Automated Manifold Surgery: Constructing Geometrically Accurate and Topologically Correct Models of the Human Cerebral Cortex. IEEE Trans. Med. Imaging 2001, 20, 70–80. [Google Scholar] [CrossRef]
- Fischl, B.; Salat, D.H.; Busa, E.; Albert, M.; Dieterich, M.; Haselgrove, C.; Van Der Kouwe, A.; Killiany, R.; Kennedy, D.; Klaveness, S.; et al. Whole Brain Segmentation. Neuron 2002, 33, 341–355. [Google Scholar] [CrossRef]
- Fischl, B.; Salat, D.H.; Van Der Kouwe, A.J.W.; Makris, N.; Ségonne, F.; Quinn, B.T.; Dale, A.M. Sequence-Independent Segmentation of Magnetic Resonance Images. NeuroImage 2004, 23, S69–S84. [Google Scholar] [CrossRef]
- Fischl, B. Automatically Parcellating the Human Cerebral Cortex. Cereb. Cortex 2004, 14, 11–22. [Google Scholar] [CrossRef]
- Fischl, B.; Sereno, M.I.; Dale, A.M. Cortical Surface-Based Analysis. NeuroImage 1999, 9, 195–207. [Google Scholar] [CrossRef]
- Fischl, B.; Sereno, M.I.; Tootell, R.B.H.; Dale, A.M. High-Resolution Intersubject Averaging and a Coordinate System for the Cortical Surface. Hum. Brain Mapp. 1999, 8, 272–284. [Google Scholar] [CrossRef]
- Han, X.; Jovicich, J.; Salat, D.; Van Der Kouwe, A.; Quinn, B.; Czanner, S.; Busa, E.; Pacheco, J.; Albert, M.; Killiany, R.; et al. Reliability of MRI-Derived Measurements of Human Cerebral Cortical Thickness: The Effects of Field Strength, Scanner Upgrade and Manufacturer. NeuroImage 2006, 32, 180–194. [Google Scholar] [CrossRef] [PubMed]
- Jovicich, J.; Czanner, S.; Greve, D.; Haley, E.; Van Der Kouwe, A.; Gollub, R.; Kennedy, D.; Schmitt, F.; Brown, G.; MacFall, J.; et al. Reliability in Multi-Site Structural MRI Studies: Effects of Gradient Non-Linearity Correction on Phantom and Human Data. NeuroImage 2006, 30, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Reuter, M.; Rosas, H.D.; Fischl, B. Highly Accurate Inverse Consistent Registration: A Robust Approach. NeuroImage 2010, 53, 1181–1196. [Google Scholar] [CrossRef] [PubMed]
- Reuter, M.; Schmansky, N.J.; Rosas, H.D.; Fischl, B. Within-Subject Template Estimation for Unbiased Longitudinal Image Analysis. NeuroImage 2012, 61, 1402–1418. [Google Scholar] [CrossRef] [PubMed]
- Ségonne, F.; Dale, A.M.; Busa, E.; Glessner, M.; Salat, D.; Hahn, H.K.; Fischl, B. A Hybrid Approach to the Skull Stripping Problem in MRI. NeuroImage 2004, 22, 1060–1075. [Google Scholar] [CrossRef]
- Tustison, N.J.; Avants, B.B.; Cook, P.A.; Zheng, Y.; Egan, A.; Yushkevich, P.A.; Gee, J.C. N4ITK: Improved N3 Bias Correction. IEEE Trans. Med. Imaging 2010, 29, 1310–1320. [Google Scholar] [CrossRef]
- Avants, B.; Epstein, C.; Grossman, M.; Gee, J. Symmetric Diffeomorphic Image Registration with Cross-Correlation: Evaluating Automated Labeling of Elderly and Neurodegenerative Brain. Med. Image Anal. 2008, 12, 26–41. [Google Scholar] [CrossRef]
- Zhang, Y.; Brady, M.; Smith, S. Segmentation of Brain MR Images through a Hidden Markov Random Field Model and the Expectation-Maximization Algorithm. IEEE Trans. Med. Imaging 2001, 20, 45–57. [Google Scholar] [CrossRef]
- Fonov, V.; Evans, A.; McKinstry, R.; Almli, C.; Collins, D. Unbiased Nonlinear Average Age-Appropriate Brain Templates from Birth to Adulthood. NeuroImage 2009, 47, S102. [Google Scholar] [CrossRef]
- Jenkinson, M.; Smith, S. A Global Optimisation Method for Robust Affine Registration of Brain Images. Med. Image Anal. 2001, 5, 143–156. [Google Scholar] [CrossRef]
- Greve, D.N.; Fischl, B. Accurate and Robust Brain Image Alignment Using Boundary-Based Registration. NeuroImage 2009, 48, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Jenkinson, M.; Bannister, P.; Brady, M.; Smith, S. Improved Optimization for the Robust and Accurate Linear Registration and Motion Correction of Brain Images. NeuroImage 2002, 17, 825–841. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.W.; Hyde, J.S. Software Tools for Analysis and Visualization of fMRI Data. NMR Biomed. 1997, 10, 171–178. [Google Scholar] [CrossRef]
- Power, J.D.; Mitra, A.; Laumann, T.O.; Snyder, A.Z.; Schlaggar, B.L.; Petersen, S.E. Methods to Detect, Characterize, and Remove Motion Artifact in Resting State fMRI. NeuroImage 2014, 84, 320–341. [Google Scholar] [CrossRef]
- Behzadi, Y.; Restom, K.; Liau, J.; Liu, T.T. A Component Based Noise Correction Method (CompCor) for BOLD and Perfusion Based fMRI. NeuroImage 2007, 37, 90–101. [Google Scholar] [CrossRef]
- Satterthwaite, T.D.; Elliott, M.A.; Gerraty, R.T.; Ruparel, K.; Loughead, J.; Calkins, M.E.; Eickhoff, S.B.; Hakonarson, H.; Gur, R.C.; Gur, R.E.; et al. An Improved Framework for Confound Regression and Filtering for Control of Motion Artifact in the Preprocessing of Resting-State Functional Connectivity Data. NeuroImage 2013, 64, 240–256. [Google Scholar] [CrossRef]
- Lanczos, C. Evaluation of Noisy Data. J. Soc. Ind. Appl. Math. Ser. B Numer. Anal. 1964, 1, 76–85. [Google Scholar] [CrossRef]
- Thomas Yeo, B.T.; Krienen, F.M.; Sepulcre, J.; Sabuncu, M.R.; Lashkari, D.; Hollinshead, M.; Roffman, J.L.; Smoller, J.W.; Zöllei, L.; Polimeni, J.R.; et al. The Organization of the Human Cerebral Cortex Estimated by Intrinsic Functional Connectivity. J. Neurophysiol. 2011, 106, 1125–1165. [Google Scholar] [CrossRef]
- Feng, W.; Liu, G.; Zeng, K.; Zeng, M.; Liu, Y. A Review of Methods for Classification and Recognition of ASD Using fMRI Data. J. Neurosci. Methods 2022, 368, 109456. [Google Scholar] [CrossRef]
- Santana, C.P.; De Carvalho, E.A.; Rodrigues, I.D.; Bastos, G.S.; De Souza, A.D.; De Brito, L.L. Rs-fMRI and Machine Learning for ASD Diagnosis: A Systematic Review and Meta-Analysis. Sci. Rep. 2022, 12, 6030. [Google Scholar] [CrossRef]
- Urchs, S.G.; Tam, A.; Orban, P.; Moreau, C.; Benhajali, Y.; Nguyen, H.D.; Evans, A.C.; Bellec, P. Functional Connectivity Subtypes Associate Robustly with ASD Diagnosis. eLife 2022, 11, e56257. [Google Scholar] [CrossRef] [PubMed]
- Philip, R.C.M.; Dauvermann, M.R.; Whalley, H.C.; Baynham, K.; Lawrie, S.M.; Stanfield, A.C. A Systematic Review and Meta-Analysis of the fMRI Investigation of Autism Spectrum Disorders. Neurosci. Biobehav. Rev. 2012, 36, 901–942. [Google Scholar] [CrossRef] [PubMed]
- Knaus, T.A.; Silver, A.M.; Lindgren, K.A.; Hadjikhani, N.; Tager-Flusberg, H. fMRI Activation during a Language Task in Adolescents with ASD. J. Int. Neuropsychol. Soc. 2008, 14, 967–979. [Google Scholar] [CrossRef] [PubMed]
- Larson, C.; Thomas, H.R.; Crutcher, J.; Stevens, M.C.; Eigsti, I.-M. Language Networks in Autism Spectrum Disorder: A Systematic Review of Connectivity-Based fMRI Studies. Rev. J. Autism. Dev. Disord. 2023, 1–28. [Google Scholar] [CrossRef]
- Cooper, R.A.; Richter, F.R.; Bays, P.M.; Plaisted-Grant, K.C.; Baron-Cohen, S.; Simons, J.S. Reduced Hippocampal Functional Connectivity During Episodic Memory Retrieval in Autism. Cereb. Cortex 2017, 27, 888–902. [Google Scholar] [CrossRef]
- Eigsti, I.-M.; Stevens, M.C.; Schultz, R.T.; Barton, M.; Kelley, E.; Naigles, L.; Orinstein, A.; Troyb, E.; Fein, D.A. Language Comprehension and Brain Function in Individuals with an Optimal Outcome from Autism. NeuroImage Clin. 2016, 10, 182–191. [Google Scholar] [CrossRef]
- Hogeveen, J.; Krug, M.K.; Geddert, R.M.; Ragland, J.D.; Solomon, M. Compensatory Hippocampal Recruitment Supports Preserved Episodic Memory in Autism Spectrum Disorder. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2020, 5, 97–109. [Google Scholar] [CrossRef]
- Tseng, A.; Camchong, J.; Francis, S.M.; Mueller, B.A.; Lim, K.O.; Conelea, C.A.; Jacob, S. Differential Extrinsic Brain Network Connectivity and Social Cognitive Task-Specific Demands in Autism Spectrum Disorder (ASD). J. Psychiatr. Res. 2022, 148, 230–239. [Google Scholar] [CrossRef]
- Pua, E.P.K.; Malpas, C.B.; Bowden, S.C.; Seal, M.L. Different Brain Networks Underlying Intelligence in Autism Spectrum Disorders. Hum. Brain Mapp. 2018, 39, 3253–3262. [Google Scholar] [CrossRef]
- Arutiunian, V.; Gomozova, M.; Minnigulova, A.; Davydova, E.; Pereverzeva, D.; Sorokin, A.; Tyushkevich, S.; Mamokhina, U.; Danilina, K.; Dragoy, O. Structural Brain Abnormalities and Their Association with Language Impairment in School-Aged Children with Autism Spectrum Disorder. Sci. Rep. 2023, 13, 1172. [Google Scholar] [CrossRef]
- Herbert, M.R.; Harris, G.J.; Adrien, K.T.; Ziegler, D.A.; Makris, N.; Kennedy, D.N.; Lange, N.T.; Chabris, C.F.; Bakardjiev, A.; Hodgson, J.; et al. Abnormal Asymmetry in Language Association Cortex in Autism. Ann. Neurol. 2002, 52, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Qiu, M.; Ye, Z.; Li, Q.; Liu, G.; Xie, B.; Wang, J. Changes of Brain Structure and Function in ADHD Children. Brain Topogr. 2011, 24, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Schafer, R.J.; Lacadie, C.; Vohr, B.; Kesler, S.R.; Katz, K.H.; Schneider, K.C.; Pugh, K.R.; Makuch, R.W.; Reiss, A.L.; Constable, R.T.; et al. Alterations in Functional Connectivity for Language in Prematurely Born Adolescents. Brain 2009, 132, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Wolf, R.C.; Plichta, M.M.; Sambataro, F.; Fallgatter, A.J.; Jacob, C.; Lesch, K.-P.; Herrmann, M.J.; Schönfeldt-Lecuona, C.; Connemann, B.J.; Grön, G.; et al. Regional Brain Activation Changes and Abnormal Functional Connectivity of the Ventrolateral Prefrontal Cortex during Working Memory Processing in Adults with Attention-Deficit/Hyperactivity Disorder. Hum. Brain Mapp. 2009, 30, 2252–2266. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Jiang, X.; Wang, X.; Liu, H.; Qian, A.; Yang, C.; Chen, H.; Li, J.; Ye, Q.; Wang, J.; et al. Disrupted Control-Related Functional Brain Networks in Drug-Naive Children with Attention-Deficit/Hyperactivity Disorder. Front. Psychiatry 2017, 8, 246. [Google Scholar] [CrossRef]
- Lee, Y.; Park, B.; James, O.; Kim, S.-G.; Park, H. Autism Spectrum Disorder Related Functional Connectivity Changes in the Language Network in Children, Adolescents and Adults. Front. Hum. Neurosci. 2017, 11, 418. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Children without ASD (n = 19) | Children with ASD (n = 16) | t Value | p | Effect Size | |
|---|---|---|---|---|---|
| Age (years) | |||||
| M (SD) | 9.55 (1.56) | 10.15 (1.32) | −1.22 | 0.23 | 0.42 |
| Range | 7.20–12.90 | 4.60–12.00 | |||
| IQ | |||||
| M (SD) | 110.84 (11.44) | 102.75 (19.81) | −7.86 | 0.16 | 0.50 |
| Range | 82–130 | 62–137 | |||
| Gender | |||||
| Male | 13 | 13 | −0.85 | 0.40 | 0.43 |
| Female | 6 | 3 | |||
| AQ total | |||||
| M (SD) | 13.00 (6.94) | 32.63 (7.83) | −7.86 | p < 0.01 | 2.65 |
| Range | 2–26 | 14–42 |
| Domain | Test | Subtest |
|---|---|---|
| Executive functions | Behavior Rating Inventory of Executive Function (BRIEF-2) | All |
| Behavior mental health | Behavior Assessment System for Children (BASC-2): Parents questionnaire | All |
| Cognitive ability | Wechsler Abbreviated Scale of Intelligence (WASI-II) | Intelligence Quotient (IQ) |
| Social competence | Chandler Cartoons Questionnaire | Sandcastle, Airplane, and Snowman cartoons |
| Social Style Questionnaire (4 to 12 years old) | All | |
| Social Responsiveness Scale (SRS 2) | All | |
| Multidimensional Social Competence Scale (MSCS) | All | |
| Pragmatic language | Comprehensive Assessment of Spoken Language (CASL 2) | Pragmatic languages |
| Scales (t Scores) | Children without ASD (n = 19) | Children with ASD (n = 16) | t Value | p | Effect Size |
|---|---|---|---|---|---|
| BRI | |||||
| M (SD) | 16.58 (5.04) | 26.94 (5.31) | −5.91 | p < 0.01 | 2.00 |
| Range | 12–29.0 | 17–34 | |||
| Inhibit | |||||
| M (SD) | 11.53 (3.64) | 17.63 (3.88) | −4.76 | p < 0.01 | 1.62 |
| Range | 8–20.0 | 10–22.0 | |||
| Self-monitor | |||||
| M (SD) | 5.05 (1.75) | 9.31 (1.70) | −7.29 | p < 0.01 | 2.47 |
| Range | 4–9.0 | 7–12.0 | |||
| ERI | |||||
| M (SD) | 22.68 (7.21) | 37.00 (7.29) | −5.82 | p < 0.01 | 1.97 |
| Range | 16–42 | 21–47 | |||
| Shift | |||||
| M (SD) | 11.32 (3.00) | 18.88 (3.76) | −6.62 | p < 0.01 | 2.22 |
| Range | 8–19.0 | 12–24.0 | |||
| Emotional control | |||||
| M (SD) | 11.37 (4.45) | 18.13 (4.84) | −4.30 | p < 0.01 | 1.45 |
| Range | 8–23.0 | 9–24.0 | |||
| CRI | |||||
| M (SD) | 50.11 (12.75) | 74.19 (14.73) | −5.19 | p < 0.01 | 1.75 |
| Range | 33–77 | 47–94 | |||
| Initiate | |||||
| M (SD) | 7.16 (1.74) | 11.75 (2.35) | −6.46 | p < 0.01 | 2.22 |
| Range | 5–11.0 | 8–15.0 | |||
| Working memory | |||||
| M (SD) | 11.42 (3.19) | 17.69 (4.45) | −4.84 | p < 0.01 | 1.62 |
| Range | 8–18.0 | 8–23.0 | |||
| Plan/organize | |||||
| M (SD) | 12.68 (3.45) | 19.75 (3.30) | −6.16 | p < 0.01 | 2.10 |
| Range | 8–20.0 | 14–24 | |||
| Task-monitor | |||||
| M (SD) | 8.58 (2.71) | 12.38 (3.01) | −3.92 | p < 0.01 | 1.33 |
| Range | 5–13.0 | 7–15.0 | |||
| Organization of materials | |||||
| M (SD) | 10.26 (3.02) | 12.63 (3.69) | −2.09 | 0.45 | 0.7 |
| Range | 6–18.0 | 7–18.0 | |||
| GEC | |||||
| M (SD) | 89.37 (22.96) | 138.13 (24.86) | −6.03 | p < 0.01 | 2.04 |
| Range | 61–144 | 91–169 | |||
| Scales (t Scores) | Children without ASD (n = 19) | Children with ASD (n = 16) | t Value | p | Effect Size |
|---|---|---|---|---|---|
| Social skill | |||||
| M (SD) | 1.74 (1.91) | 5.81 (2.46) | −5.52 | p < 0.01 | 1.85 |
| Range | 0–8 | 1–10.0 | |||
| Attention switching | |||||
| M (SD) | 3.21 (2.18) | 7.13 (2.53) | −4.93 | p < 0.01 | 1.66 |
| Range | 0–7 | 2–10.0 | |||
| Attention to detail | |||||
| M (SD) | 4.53 (2.88) | 6.56 (2.10) | −2.35 | 0.03 | 0.81 |
| Range | 0–10 | 2–10.0 | |||
| Communication | |||||
| M (SD) | 1.58 (1.35) | 7.00 (2.42) | −7.98 | p < 0.01 | 2.77 |
| Range | 0–5 | 1–10.0 | |||
| Imagination | |||||
| M (SD) | 1.95 (1.03) | 6.13 (2.06) | −7.37 | p < 0.01 | 2.57 |
| Range | 0–4 | 2.0–9 | |||
| AQ total | |||||
| M (SD) | 13.00 (6.94) | 32.63 (7.83) | −7.86 | p < 0.01 | 2.65 |
| Range | 2.0–26 | 14–42 | |||
| Scales (t Scores) | Children without ASD (n = 19) | Children with ASD (n = 16) | t Value | p | Effect Size |
|---|---|---|---|---|---|
| Social motivation | |||||
| M (SD) | 43.05 (7.47) | 32.56 (10.47) | 3.45 | p < 0.01 | 1.15 |
| Range | 28–55 | 11–52 | |||
| Social inferencing | |||||
| M (SD) | 44.68 (6.46) | 24.63 (8.35) | 8.01 | p < 0.01 | 2.69 |
| Range | 29–53 | 15–49 | |||
| Empathic concern | |||||
| M (SD) | 46.00 (6.88) | 30.69 (8.74) | 5.80 | p < 0.01 | 1.95 |
| Range | 31–55 | 16–54 | |||
| Social knowledge | |||||
| M (SD) | 47.53 (6.70) | 35.25 (8.20) | 4.88 | p < 0.01 | 1.64 |
| Range | 30–54 | 19–48 | |||
| Verbal conversation | |||||
| M (SD) | 41.68 (8.38) | 27.06 (9.70) | 4.79 | p < 0.01 | 1.61 |
| Range | 22–53 | 13–49 | |||
| Nonverbal conversation | |||||
| M (SD) | 49.95 (4.62) | 37.75 (10.20) | 4.42 | p < 0.01 | 1.54 |
| Range | 39–55 | 19–53 | |||
| Emotional regulation | |||||
| M (SD) | 41.58 (8.09) | 27.69 (9.24) | 4.74 | p < 0.01 | 1.60 |
| Range | 22–51 | 13–43 | |||
| MSCS total | |||||
| M (SD) | 314.47 (39.99) | 215.63 (46.36) | 6.78 | p < 0.01 | 2.28 |
| Range | 234–364 | 134–329 | |||
| Scales (t Scores) | Children without ASD (n = 19) | Children with ASD (n = 16) | t Value | p | Effect Size |
|---|---|---|---|---|---|
| Social awareness | |||||
| M (SD) | 45.11 (9.68) | 67.69 (13.27) | −5.81 | p < 0.01 | 1.94 |
| Range | 34–70 | 35–89 | |||
| Social cognition | |||||
| M (SD) | 45.21 (7.38) | 73.00 (10.55) | −8.87 | p < 0.01 | 3.05 |
| Range | 39–63 | 55–90 | |||
| Social communication | |||||
| M (SD) | 45.89 (7.56) | 73.00 (11.90) | −8.17 | p < 0.01 | 2.72 |
| Range | 38–65 | 44–90 | |||
| Social motivation | |||||
| M (SD) | 49.16 (10.29) | 68.75 (12.48) | −5.09 | p < 0.01 | 1.71 |
| Range | 38–75 | 46–90 | |||
| Restricted interests/repetitiveness | |||||
| M (SD) | 49.00 (8.52) | 74.75 (12.50) | −7.22 | p < 0.01 | 2.41 |
| Range | 41–71 | 50–90 | |||
| Social communication/interaction | |||||
| M (SD) | 46.05 (8.22) | 73.56 (11.29) | −8.33 | p < 0.01 | 2.79 |
| Range | 38–66 | 46–90 | |||
| SRS-2 total | |||||
| M (SD) | 46.37 (8.24) | 74.50 (11.02) | −8.63 | p < 0.01 | 2.89 |
| Range | 38–67 | 47–90 | |||
| Scales (t Scores) | Children without ASD (n = 19) | Children with ASD (n = 16) | t Value | p | Effect Size |
|---|---|---|---|---|---|
| CASL-2 Pragmatic language | |||||
| M (SD) | 109.84 (14.98) | 90.19 (13.32) | 4.07 | p < 0.01 | 1.38 |
| Range | 81–141 | 70–117 | |||
| Scales (t Scores) | Children without ASD (n = 19) | Children with ASD (n = 16) | t Value | p | Effect Size |
|---|---|---|---|---|---|
| WASI-2 IQ | |||||
| M (SD) | 110.84 (11.44) | 102.75 (19.81) | −7.86 | 0.16 | 0.5 |
| Range | 82–130 | 62–137 |
| Scales (t Scores) | Children without ASD (n = 19) | Children with ASD (n = 16) | t Value | p | Effect Size |
|---|---|---|---|---|---|
| Bystander cartoon total | |||||
| M (SD) | 9.00 (4.06) | 6.31 (4.63) | 1.83 | 0.08 | 0.62 |
| Range | 0–12 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Márquez-García, A.V.; Ng, B.K.; Iarocci, G.; Moreno, S.; Vakorin, V.A.; Doesburg, S.M. Atypical Associations between Functional Connectivity during Pragmatic and Semantic Language Processing and Cognitive Abilities in Children with Autism. Brain Sci. 2023, 13, 1448. https://doi.org/10.3390/brainsci13101448
Márquez-García AV, Ng BK, Iarocci G, Moreno S, Vakorin VA, Doesburg SM. Atypical Associations between Functional Connectivity during Pragmatic and Semantic Language Processing and Cognitive Abilities in Children with Autism. Brain Sciences. 2023; 13(10):1448. https://doi.org/10.3390/brainsci13101448
Chicago/Turabian StyleMárquez-García, Amparo V., Bonnie K. Ng, Grace Iarocci, Sylvain Moreno, Vasily A. Vakorin, and Sam M. Doesburg. 2023. "Atypical Associations between Functional Connectivity during Pragmatic and Semantic Language Processing and Cognitive Abilities in Children with Autism" Brain Sciences 13, no. 10: 1448. https://doi.org/10.3390/brainsci13101448
APA StyleMárquez-García, A. V., Ng, B. K., Iarocci, G., Moreno, S., Vakorin, V. A., & Doesburg, S. M. (2023). Atypical Associations between Functional Connectivity during Pragmatic and Semantic Language Processing and Cognitive Abilities in Children with Autism. Brain Sciences, 13(10), 1448. https://doi.org/10.3390/brainsci13101448