
Network-Based Differences in Top–Down Multisensory Integration between Adult ADHD and Healthy Controls—A Diffusion MRI Study
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. MRI Protocol
2.3. Diffusion MRI Data Analysis
2.4. Graph Theory
3. Results
3.1. Demographic Variables
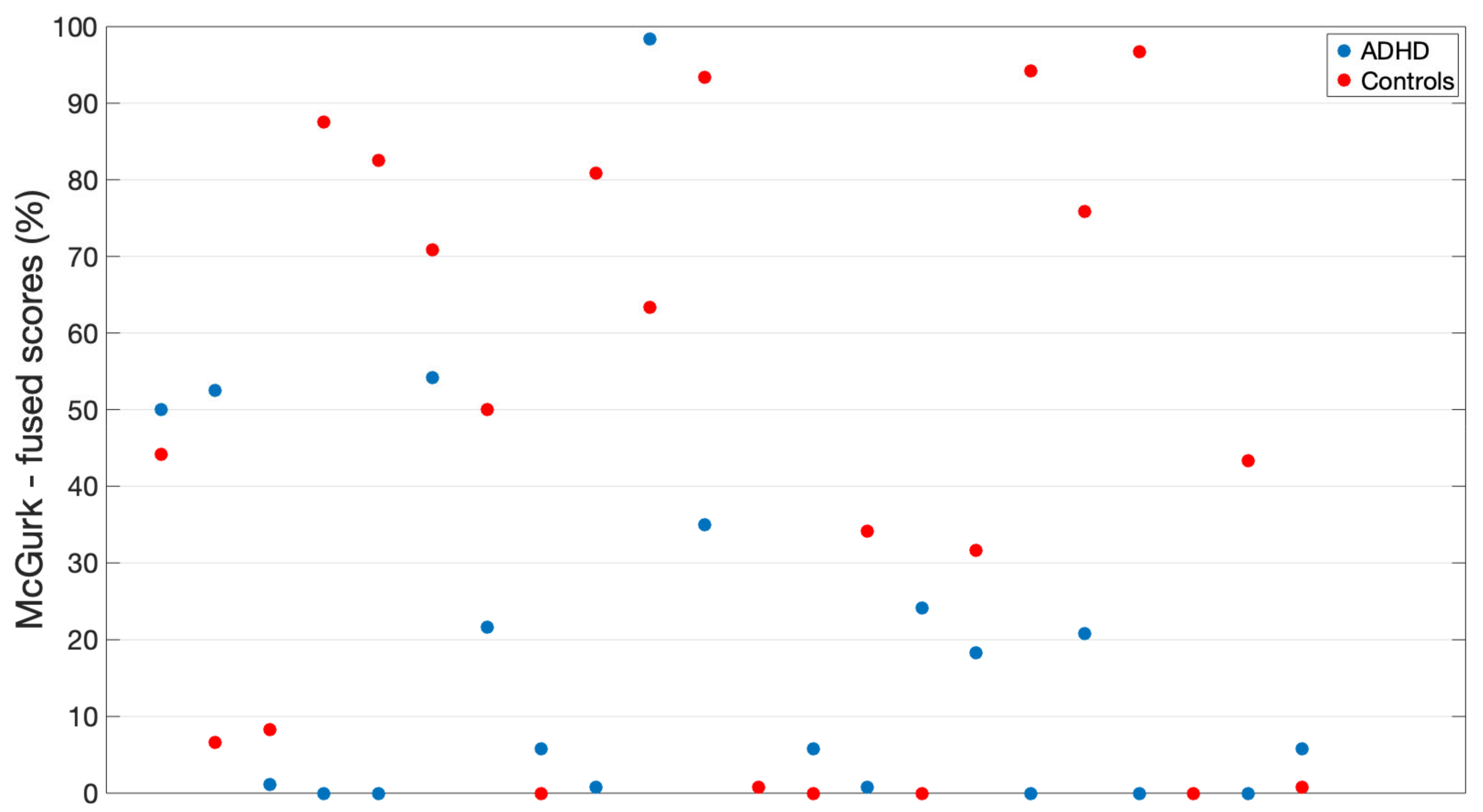
3.2. McGurk Audiovisual Integration
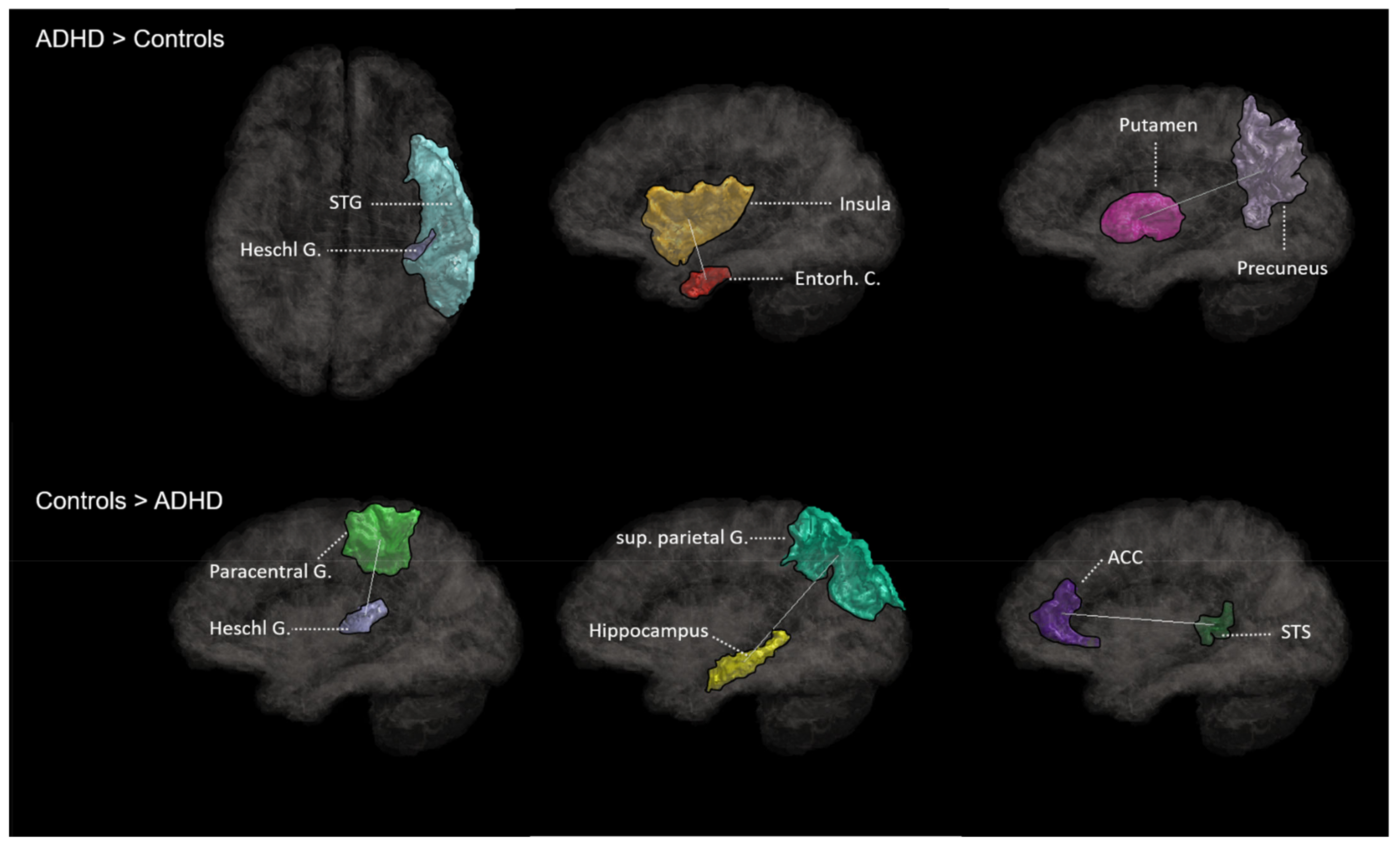
3.3. Network-Based WM Connectivity
3.3.1. ADHD > Controls
3.3.2. ADHD < Controls
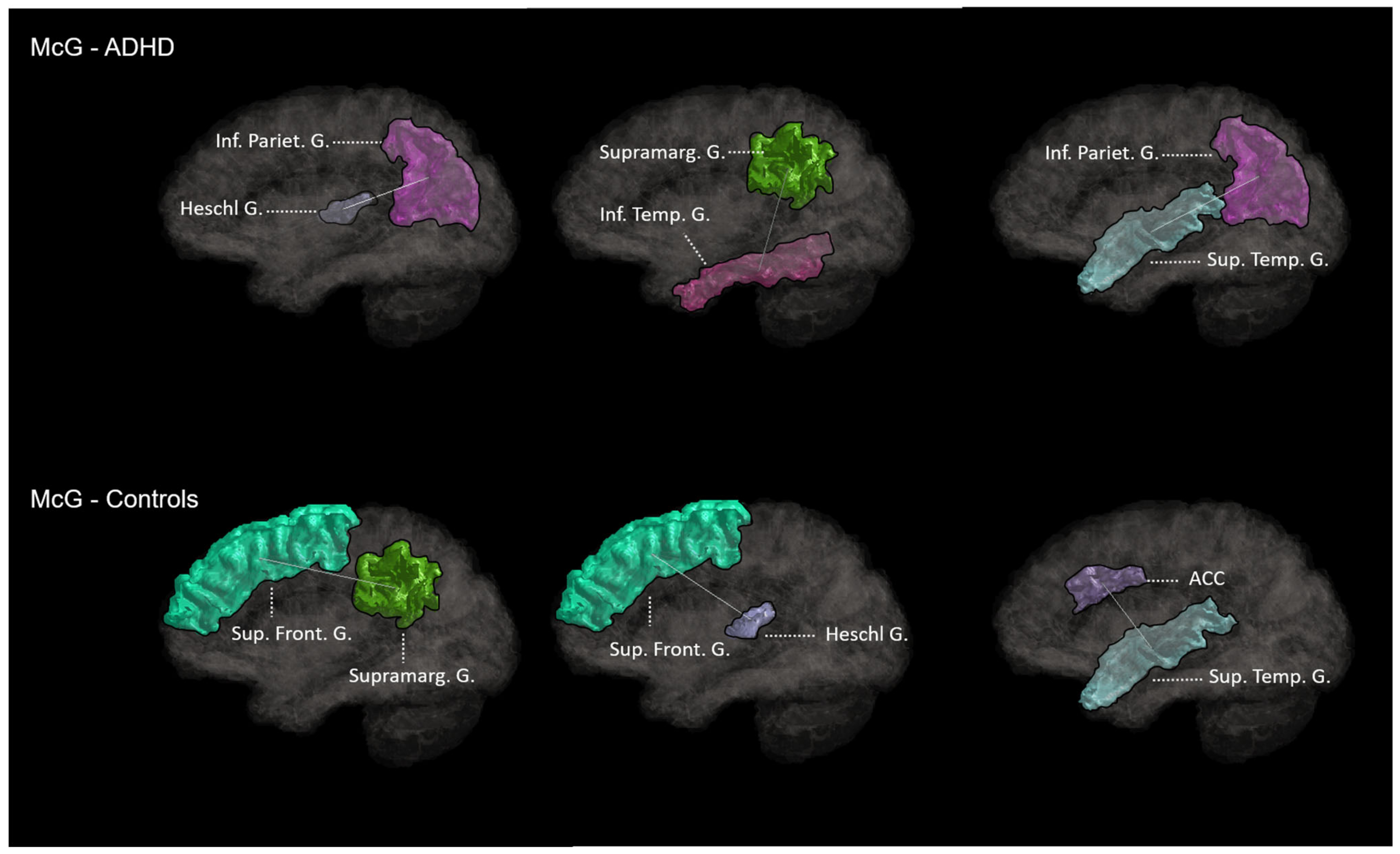
3.4. Association McGurk-Effect–WM Connectivity
3.4.1. ADHD
3.4.2. Controls
3.5. Graph Theory
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Willcutt, E.G.; Nigg, J.T.; Pennington, B.F.; Solanto, M.V.; Rohde, L.A.; Tannock, R.; Loo, S.K.; Carlson, C.L.; McBurnett, K.; Lahey, B.B. Validity of DSM-IV Attention Deficit/Hyperactivity Disorder Symptom Dimensions and Subtypes. J. Abnorm. Psychol. 2012, 121, 991–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sibley, M.H.; Mitchell, J.T.; Becker, S.P. Method of Adult Diagnosis Influences Estimated Persistence of Childhood ADHD: A Systematic Review of Longitudinal Studies. Lancet Psychiatry 2016, 3, 1157–1165. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.E. ADD/ADHD and Impaired Executive Function in Clinical Practice. Curr. Atten. Disord. Rep. 2009, 1, 37–41. [Google Scholar] [CrossRef]
- Ghanizadeh, A. Sensory Processing Problems in Children with ADHD, a Systematic Review. Psychiatry Investig. 2011, 8, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze, M.; Lux, S.; Philipsen, A. Sensory Processing in Adult ADHD—A Systematic Review. Res. Sq. 2020. PREPRINT (Version 1). [Google Scholar] [CrossRef]
- Barry, R.J.; Clarke, A.R.; McCarthy, R.; Selikowitz, M.; Brown, C.R.; Heaven, P.C.L. Event-Related Potentials in Adults with Attention-Deficit/Hyperactivity Disorder: An Investigation Using an Inter-Modal Auditory/Visual Oddball Task. International J. Psychophysiol. Off. J. Int. Organ. Psychophysiol. 2009, 71, 124–131. [Google Scholar] [CrossRef]
- Fostick, L. The Effect of Attention-Deficit/Hyperactivity Disorder and Methylphenidate Treatment on the Adult Auditory Temporal Order Judgment Threshold. J. Speech Lang. Hear. Res. 2017, 60, 2124–2128. [Google Scholar] [CrossRef] [Green Version]
- Dhamala, M.; Assisi, C.G.; Jirsa, V.K.; Steinberg, F.L.; Scott Kelso, J.A. Multisensory Integration for Timing Engages Different Brain Networks. NeuroImage 2007, 34, 764–773. [Google Scholar] [CrossRef] [Green Version]
- Schulze, M.; Aslan, B.; Jung, P.; Lux, S.; Philipsen, A. Robust Perceptual-Load-Dependent Audiovisual Integration in Adult ADHD. Eur. Arch. Psychiatry Clin. Neurosci. 2022, 272, 1443–1451. [Google Scholar] [CrossRef] [PubMed]
- McCracken, H.S.; Murphy, B.A.; Glazebrook, C.M.; Burkitt, J.J.; Karellas, A.M.; Yielder, P.C. Audiovisual Multisensory Integration and Evoked Potentials in Young Adults with and without Attention-Deficit/Hyperactivity Disorder. Front. Hum. Neurosci. 2019, 13, 95. [Google Scholar] [CrossRef]
- Michalek, A.M.P.; Watson, S.M.; Ash, I.; Ringleb, S.; Raymer, A. Effects of Noise and Audiovisual Cues on Speech Processing in Adults with and without ADHD. Int. J. Audiol. 2014, 53, 145–152. [Google Scholar] [CrossRef]
- Schulze, M.; Aslan, B.; Stöcker, T.; Stirnberg, R.; Lux, S.; Philipsen, A. Disentangling Early versus Late Audiovisual Integration in Adult ADHD: A Combined Behavioural and Resting-State Connectivity Study. J. Psychiatry Neurosci. 2021, 46, E528–E537. [Google Scholar] [CrossRef]
- MacAluso, E.; Noppeney, U.; Talsma, D.; Vercillo, T.; Hartcher-O’Brien, J.; Adam, R. The Curious Incident of Attention in Multisensory Integration: Bottom-up vs. Top-Down. Multisens. Res. 2016, 29, 557–583. [Google Scholar] [CrossRef] [Green Version]
- Raichle, M.E. The Brain’s Default Mode Network. Annu. Rev. Neurosci. 2015, 38, 443–447. [Google Scholar] [CrossRef] [Green Version]
- Saad, J.F.; Griffiths, K.R.; Korgaonkar, M.S. A Systematic Review of Imaging Studies in the Combined and Inattentive Subtypes of Attention Deficit Hyperactivity Disorder. Front. Integr. Neurosci. 2020, 14, 31. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Sanchez, V.; Castellanos, F.X. Neuroimaging in Attention-Deficit/Hyperactivity Disorder. Curr. Opin. Psychiatry 2021, 34, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Albajara Sáenz, A.; Villemonteix, T.; Massat, I. Structural and Functional Neuroimaging in Attention-Deficit/Hyperactivity Disorder. Dev. Med. Child Neurol. 2019, 61, 399–405. [Google Scholar] [CrossRef] [Green Version]
- Mcgurk, H.; Macdonald, J. Hearing Lips and Seeing Voices. Nature 1976, 264, 746–748. [Google Scholar] [CrossRef]
- Stropahl, M.; Schellhardt, S.; Debener, S. McGurk Stimuli for the Investigation of Multisensory Integration in Cochlear Implant Users: The Oldenburg Audio Visual Speech Stimuli (OLAVS). Psychon. Bull. Rev. 2017, 24, 863–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- First, M.B. Structured Clinical Interview for the DSM (SCID). In The Encyclopedia of Clinical Psychology; Baskı, B., Cautin, R.L., Lilienfeld, S.O., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2015. [Google Scholar]
- Heinzl, S. Neue S3-Leitlinie “ADHS Bei Kindern, Jugendlichen Und Erwachsenen“. DNP—Der Neurol. Psychiater 2018, 19, 60. [Google Scholar] [CrossRef]
- Atkinson, M.; Hollis, C. NICE Guideline: Attention Deficit Hyperactivity Disorder. Arch. Dis. Child.—Educ. Pract. 2010, 95, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, H.; Kis, B.; Hirsch, O.; Matthies, S.; Hebebrand, J.; Uekermann, J.; Abdel-Hamid, M.; Kraemer, M.; Wiltfang, J.; Graf, E.; et al. German Validation of the Conners Adult ADHD Rating Scales (CAARS) II: Reliability, Validity, Diagnostic Sensitivity and Specificity. Eur. Psychiatry 2012, 27, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Retz-Junginger, P.; Retz, W.; Blocher, D.; Weijers, H.G.; Trott, G.E.; Wender, P.H.; Rössler, M. Wender Utah Rating Scale (WURS-k): Die Deutsche Kurzform Zur Retrospektiven Erfassung Des Hyperkinetischen Syndroms Bei Erwachsenen. Nervenarzt 2002, 73, 830–838. [Google Scholar] [CrossRef] [PubMed]
- Breuer, F.A.; Blaimer, M.; Mueller, M.F.; Seiberlich, N.; Heidemann, R.M.; Griswold, M.A.; Jakob, P.M. Controlled Aliasing in Volumetric Parallel Imaging (2D CAIPIRINHA). Magn. Reson. Med. 2006, 55, 549–556. [Google Scholar] [CrossRef]
- Brenner, D.; Stirnberg, R.; Pracht, E.D.; Stöcker, T. Two-Dimensional Accelerated MP-RAGE Imaging with Flexible Linear Reordering. Magn. Reson. Mater. Phys. Biol. Med. 2014, 27, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Setsompop, K.; Gagoski, B.A.; Polimeni, J.R.; Witzel, T.; Wedeen, V.J.; Wald, L.L. Blipped-Controlled Aliasing in Parallel Imaging for Simultaneous Multislice Echo Planar Imaging with Reduced g-Factor Penalty. Magn. Reson. Med. 2012, 67, 1210–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tournier, J.D.; Smith, R.; Raffelt, D.; Tabbara, R.; Dhollander, T.; Pietsch, M.; Christiaens, D.; Jeurissen, B.; Yeh, C.H.; Connelly, A. MRtrix3: A Fast, Flexible and Open Software Framework for Medical Image Processing and Visualisation. NeuroImage 2019, 202, 116137. [Google Scholar] [CrossRef] [PubMed]
- Veraart, J.; Novikov, D.S.; Christiaens, D.; Ades-aron, B.; Sijbers, J.; Fieremans, E. Denoising of Diffusion MRI Using Random Matrix Theory. NeuroImage 2016, 142, 394–406. [Google Scholar] [CrossRef] [Green Version]
- Veraart, J.; Fieremans, E.; Jelescu, I.O.; Knoll, F.; Novikov, D.S. Gibbs Ringing in Diffusion MRI. Magn. Reson. Med. 2016, 76, 301–314. [Google Scholar] [CrossRef] [Green Version]
- Andersson, J.L.R.; Sotiropoulos, S.N. An Integrated Approach to Correction for Off-Resonance Effects and Subject Movement in Diffusion MR Imaging. NeuroImage 2016, 125, 1063–1078. [Google Scholar] [CrossRef] [Green Version]
- Jenkinson, M.; Beckmann, C.F.; Behrens, T.E.J.; Woolrich, M.W.; Smith, S.M. FSL. NeuroImage 2012, 62, 782–790. [Google Scholar] [CrossRef] [Green Version]
- Tustison, N.J.; Avants, B.B.; Cook, P.A.; Zheng, Y.; Egan, A.; Yushkevich, P.A.; Gee, J.C. N4ITK: Improved N3 Bias Correction. IEEE Trans. Med. Imaging 2010, 29, 1310–1320. [Google Scholar] [CrossRef] [Green Version]
- Dhollander, T.; Raffelt, D.; Connelly, A. Unsupervised 3-Tissue Response Function Estimation from Single-Shell or Multi-Shell Diffusion MR Data without a Co-Registered T1 Image. Predicting Stroke Impairment Using Machine Learning Techniques View Project A Novel Sparse Partial Correlation Method for Simultaneous Estimation of Functional Networks in Group Comparisons ViewProject; ISMRM Workshop on Breaking the Barriers of DiffusionMRI. (ISMRM) 2016. Available online: https://www.researchgate.net/publication/307863133_Unsupervised_3-tissue_response_function_estimation_from_single-shell_or_multi-shell_diffusion_MR_data_without_a_co-registered_T1_image (accessed on 19 February 2023).
- Jeurissen, B.; Tournier, J.D.; Dhollander, T.; Connelly, A.; Sijbers, J. Multi-Tissue Constrained Spherical Deconvolution for Improved Analysis of Multi-Shell Diffusion MRI Data. NeuroImage 2014, 103, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.E.; Tournier, J.D.; Calamante, F.; Connelly, A. Anatomically-Constrained Tractography: Improved Diffusion MRI Streamlines Tractography through Effective Use of Anatomical Information. NeuroImage 2012, 62, 1924–1938. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.E.; Tournier, J.D.; Calamante, F.; Connelly, A. SIFT: Spherical-Deconvolution Informed Filtering of Tractograms. NeuroImage 2013, 67, 298–312. [Google Scholar] [CrossRef] [PubMed]
- Baggio, H.C.; Abos, A.; Segura, B.; Campabadal, A.; Garcia-Diaz, A.; Uribe, C.; Compta, Y.; Marti, M.J.; Valldeoriola, F.; Junque, C. Statistical Inference in Brain Graphs Using Threshold-Free Network-Based Statistics. Hum. Brain Mapp. 2018, 39, 2289–2302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, M.; Shu, N.; Cao, Q.; Wang, Y.; He, Y. Imaging Functional and Structural Brain Connectomics in Attention-Deficit/Hyperactivity Disorder. Mol. Neurobiol. 2014, 50, 1111–1123. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, X.; Xia, M.; Liao, X.; Evans, A.; He, Y. GRETNA: A Graph Theoretical Network Analysis Toolbox for Imaging Connectomics. Front. Hum. Neurosci. 2015, 9, 386. [Google Scholar] [CrossRef]
- He, Y.; Evans, A. Graph Theoretical Modeling of Brain Connectivity. Curr. Opin. Neurol. 2010, 23, 341–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, W.; Bennett, D. Patterns of Sensory Processing in Children with Attention Deficit Hyperactivity Disorder. OTJR-Occup. Particip. Health 2002, 22, 4–15. [Google Scholar] [CrossRef]
- Cheung, P.P.P.; Siu, A.M.H. A Comparison of Patterns of Sensory Processing in Children with and without Developmental Disabilities. Res. Dev. Disabil. 2009, 30, 1468–1480. [Google Scholar] [CrossRef]
- Beer, A.L.; Plank, T.; Greenlee, M.W. Diffusion Tensor Imaging Shows White Matter Tracts between Human Auditory and Visual Cortex. Exp. Brain Res. 2011, 213, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, M.S.; Argall, B.D.; Bodurka, J.; Duyn, J.H.; Martin, A. Unraveling Multisensory Integration: Patchy Organization within Human STS Multisensory Cortex. Nat. Neurosci. 2004, 7, 1190–1192. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, M.S. Using Multisensory Integration to Understand the Human Auditory Cortex. In Multisensory Processes; Springer: Cham, Switzerland, 2019; pp. 161–176. [Google Scholar] [CrossRef]
- Beauchamp, M.S.; Lee, K.E.; Argall, B.D.; Martin, A. Integration of Auditory and Visual Information about Objects in Superior Temporal Sulcus. Neuron 2004, 41, 809–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, I.; Lee, J.Y.; Lee, S.H. Bottom-up and Top-down Modulation of Multisensory Integration. Curr. Opin. Neurobiol. 2018, 52, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Pflugshaupt, T.; Nosberger, M.; Gutbrod, K.; Weber, K.P.; Linnebank, M.; Brugger, P. Bottom-up Visual Integration in the Medial Parietal Lobe. Cereb. Cortex 2016, 26, 943–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talsma, D. Predictive Coding and Multisensory Integration: An Attentional Account of the Multisensory Mind. Front. Integr. Neurosci. 2015, 117, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Talsma, D.; Senkowski, D.; Soto-Faraco, S.; Woldorff, M.G. The Multifaceted Interplay between Attention and Multisensory Integration. Trends Cogn. Sci. 2010, 14, 400–410. [Google Scholar] [CrossRef] [Green Version]
- Michail, G.; Senkowski, D.; Niedeggen, M.; Keil, J. Memory Load Alters Perception-Related Neural Oscillations during Multisensory Integration. J. Neurosci. 2021, 41, 1505–1515. [Google Scholar] [CrossRef]
- Firouzabadi, F.D.; Ramezanpour, S.; Firouzabadi, M.D.; Yousem, I.J.; Puts, N.A.J.; Yousem, D.M. Neuroimaging in Attention-Deficit/Hyperactivity Disorder: Recent Advances. AJR Am. J. Roentgenol. 2021, 218, 321–332. [Google Scholar] [CrossRef]
- Uddin, L.Q.; Kelly, A.M.C.; Biswal, B.B.; Margulies, D.S.; Shehzad, Z.; Shaw, D.; Ghaffari, M.; Rotrosen, J.; Adler, L.A.; Castellanos, F.X.; et al. Network Homogeneity Reveals Decreased Integrity of Default-Mode Network in ADHD. J. Neurosci. Methods 2008, 169, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, K.R.; Grieve, S.M.; Kohn, M.R.; Clarke, S.; Williams, L.M.; Korgaonkar, M.S. Altered Gray Matter Organization in Children and Adolescents with ADHD: A Structural Covariance Connectome Study. Transl. Psychiatry 2016, 6, e947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Wang, G.; Wang, X.; Cao, R.; Xiang, J.; Yan, T.; Li, H.; Yoshimura, S.; Toichi, M.; Zhao, S. Rich-Club Analysis in Adults With ADHD Connectomes Reveals an Abnormal Structural Core Network. J. Atten. Disord. 2021, 25, 1068–1079. [Google Scholar] [CrossRef] [PubMed]
- Stoodley, C.J. The Cerebellum and Neurodevelopmental Disorders. Cerebellum 2016, 15, 34–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Network | z-Value | p-Value |
|---|---|---|
| ADHD > Controls | ||
| R entorhinal C.–R Insula | 2.15 | 0.032 |
| L entorhinal C.–R Cerebellum | 2.05 | 0.045 |
| L superior temp. G.–L Heschl G. | 2.05 | 0.039 |
| R Putamen–R Precuneus | 1.95 | 0.033 |
| L parsopercularis–R Precuneus | 1.94 | 0.032 |
| ADHD < Controls | ||
| R anterior cing. C.–R superior temp. sulcus | 2.73 | 0.018 |
| R Cuneus–L lingual G. | 2.67 | 0.048 |
| R Insula–R parsorbitalis | 2.6 | 0.033 |
| L Accumbens–L paracentral G. | 2.44 | 0.009 |
| R paracentral G.–L Heschl G. | 2.29 | 0.009 |
| L Insula–L parsorbitalis | 2.27 | 0.002 |
| Super. parietal G.–Hippocampus | 2.19 | 0.003 |
| McG–ADHD | ||
| R anterior cing. C. (rostral) –L anterior cing. C. (caudal) | 2.2 | 0.033 |
| R super temp. G.–R inferior parietal G. | 2.19 | 0.031 |
| L inferior temp. G.–L supramarginal G. | 2.2 | 0.035 |
| R Heschl G.–R inferior parietal G. | 2.3 | 0.032 |
| McG–Controls | ||
| L Caudate–L supramarginal G. | 2.29 | 0.044 |
| R Thalamus–L supramarginal G. | 2.28 | 0.042 |
| L supramarginal G.–L superior front. G. | 2.28 | 0.041 |
| R superior temp. G.–R anterior cing. C. | 2.08 | 0.044 |
| L superior temp. S.–R Precuneus | 2.07 | 0.043 |
| L Heschl G.–L superior front. G. | 2.06 | 0.042 |
| Measure | t-Value | p-Value |
|---|---|---|
| Degree Centrality | ||
| R postcentral G. | 2.11 | 0.042 |
| Nodal Local Efficiency | ||
| L isthmuscingulate | 2.19 | 0.034 |
| L lateraloccipital C. | 2.19 | 0.031 |
| L pericalcarine Fissure | 1.92 | 0.006 |
| R inferiorparietal G. | 2.16 | 0.033 |
| R lateraloccipital C. | 2.09 | 0.042 |
| Clustering Coefficient | ||
| R lateraloccipital C. | 2.04 | 0.046 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schulze, M.; Aslan, B.; Farrher, E.; Grinberg, F.; Shah, N.; Schirmer, M.; Radbruch, A.; Stöcker, T.; Lux, S.; Philipsen, A. Network-Based Differences in Top–Down Multisensory Integration between Adult ADHD and Healthy Controls—A Diffusion MRI Study. Brain Sci. 2023, 13, 388. https://doi.org/10.3390/brainsci13030388
Schulze M, Aslan B, Farrher E, Grinberg F, Shah N, Schirmer M, Radbruch A, Stöcker T, Lux S, Philipsen A. Network-Based Differences in Top–Down Multisensory Integration between Adult ADHD and Healthy Controls—A Diffusion MRI Study. Brain Sciences. 2023; 13(3):388. https://doi.org/10.3390/brainsci13030388
Chicago/Turabian StyleSchulze, Marcel, Behrem Aslan, Ezequiel Farrher, Farida Grinberg, Nadim Shah, Markus Schirmer, Alexander Radbruch, Tony Stöcker, Silke Lux, and Alexandra Philipsen. 2023. "Network-Based Differences in Top–Down Multisensory Integration between Adult ADHD and Healthy Controls—A Diffusion MRI Study" Brain Sciences 13, no. 3: 388. https://doi.org/10.3390/brainsci13030388
APA StyleSchulze, M., Aslan, B., Farrher, E., Grinberg, F., Shah, N., Schirmer, M., Radbruch, A., Stöcker, T., Lux, S., & Philipsen, A. (2023). Network-Based Differences in Top–Down Multisensory Integration between Adult ADHD and Healthy Controls—A Diffusion MRI Study. Brain Sciences, 13(3), 388. https://doi.org/10.3390/brainsci13030388