

Dyslexia: Causes and Concomitant Impairments

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. What Are Causes of Dyslexia?
2.1. Necessary Conditions, Sufficient Conditions, and Causes
2.2. Dyslexia Is Not Always Due to an Impaired Visual Attention Span, to Lateral Masking, or a Phononological Impairment
- (1)
- If the processing of stimuli improves when a person voluntarily attempts to process visual stimuli optimally at that location;
- (2)
- and/or if the processing of stimuli improves when stimuli are (consciously) expected at that location;
- (3)
- and/or if the processing of stimuli improves when environmental conditions indicate the location where stimuli will appear;
- (4)
- and/or when the visual system attempts to process stimuli optimally at that location even if the person does not voluntarily try to process stimuli optimally at that location.
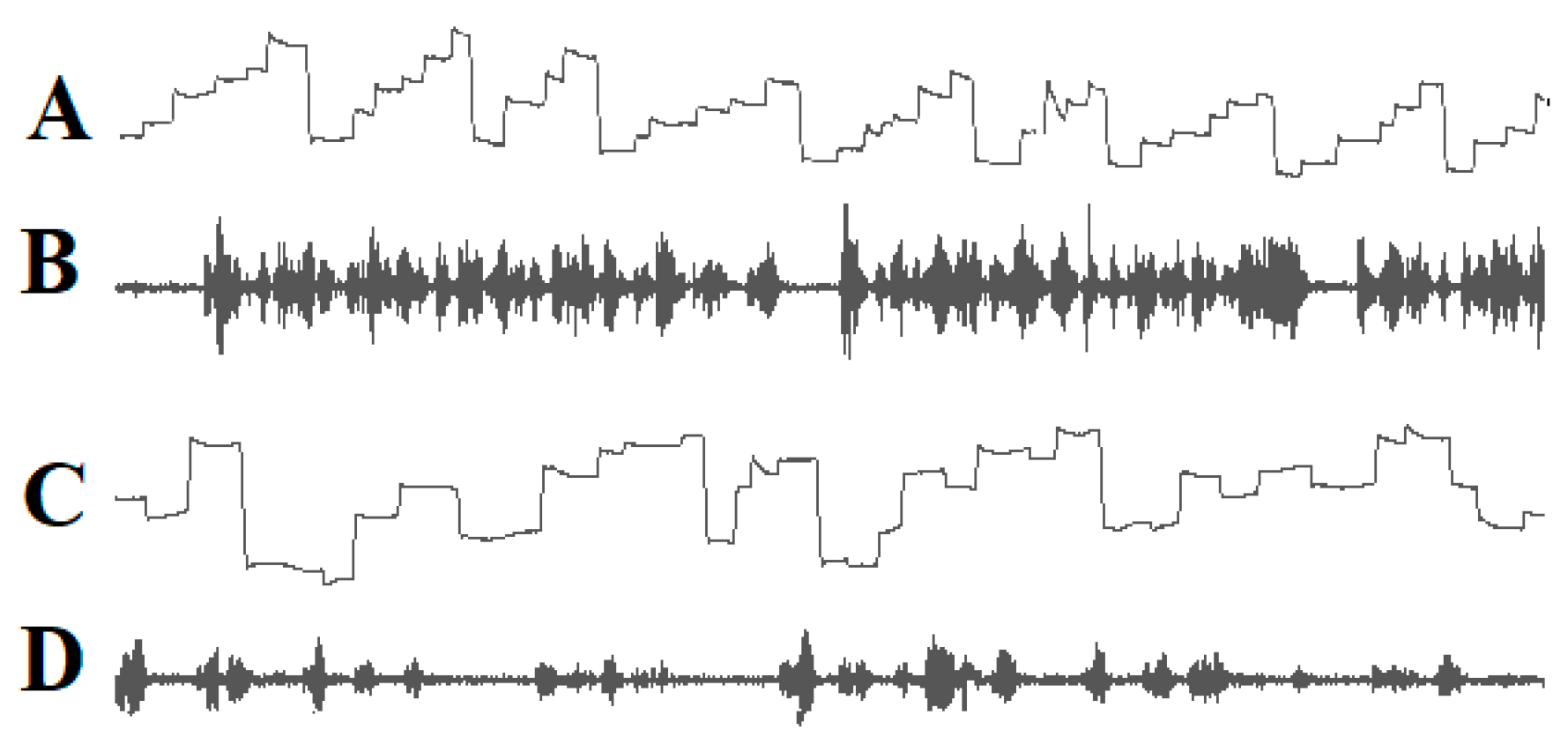
2.3. The Role of Inappropriate Reading Eye Movements in Dyslexia
- (1)
- A yellow cursor indicated the location in the word where the gaze should be directed;
- (2)
- Green cursors to the left and right of the yellow cursor indicated how many letters to the left and right of the fixation point the subject should try to recognize together with the letter at the fixation point;
- (3)
- The onset of the movement of the yellow and green cursors in the reading direction indicated when fixation on a word or word segment should be terminated;
- (4)
- The amplitude of the movement of the cursors in the reading direction indicated the direction and amplitude of the saccade to be made in order to read the next word or word segment;
- (5)
- A sound signal indicated when the subject should begin to pronounce the word or word segment to be read.
- (1)
- Fixation times are long enough to recognize a string of letters;
- (2)
- Readers do not try to recognize more letters in a word at the same time than they can;
- (3)
- Eye movement amplitudes in the reading direction do not exceed the number of letters that the reader can simultaneously recognize;
- (4)
- The reader does not start pronouncing the words or word segments to be read too early.
2.4. DD Is Not Caused by a Phonological Impairment
3. Is Dyslexia Due to an Impaired Learning Capacity?
4. What Functional MRI and Cerebral Lesions Reveal about the Neurobiological Basis of Dyslexia
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berkhan, O. Über Störungen der Sprache und der Schriftsprache: Für Ärzte und Lehrer Dargestellt; August Hirschwald: Berlin, Germany, 1889. [Google Scholar]
- Berlin, R. Über Dyslexie. Arch. Psychiat. Nervenkrankh. 1884, 15, 276–278. [Google Scholar]
- Helland, T. Trends in dyslexia research during the period 1950 to 2020—Theories, definitions, and publications. Brain Sci. 2022, 12, 1323. [Google Scholar] [CrossRef] [PubMed]
- Stein, J. Theories about developmental dyslexia. Brain Sci. 2023, 13, 208. [Google Scholar] [CrossRef] [PubMed]
- Livingstone, M.S.; Rosen, G.D.; Drislane, F.W.; Galaburda, A.M. Physiological and anatomical evidence for a magnocellular defect in developmental dyslexia. Proc. Natl. Acad. Sci. USA 1991, 88, 7943–7947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, J.; Walsh, V. To see but not to read; the magnocellular theory of dyslexia. Trends Neurosci. 1997, 20, 147–152. [Google Scholar] [CrossRef]
- Stein, J.; Walsh, V. Impaired neural timing in developmental dyslexia—The magnocellular hypothesis. Dyslexia 1999, 5, 59–77. [Google Scholar] [CrossRef]
- Stein, J. The magnocellular theory of dyslexia. In Developmental Dyslexia, Early Precursors, Neurobehavioral Markers, and Biological Substrates; Benasich, A.A., Fitch, R.H., Eds.; Brooks: Baltimore, MD, USA; London, UK; Sydney, Australia, 2012. [Google Scholar]
- Stein, J. The current status of the magnocellular theory of developmental dyslexia. Neuropsychologia 2019, 130, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Geiger, G.; Lettvin, J.Y. Peripheral vision in persons with dyslexia. N. Engl. J. Med. 1987, 316, 1238–1243. [Google Scholar] [CrossRef]
- Geiger, G.; Lettvin, J.Y.; Fahle, M. Dyslexic children learn a new visual strategy for reading: A controlled experiment. Vis. Res. 1993, 34, 1223–1233. [Google Scholar] [CrossRef]
- Spinelli, D.; De Luca, M.; Judica, A.; Zoccolotti, P. Crowding effects on word identification in developmental dyslexia. Cortex 2002, 38, 179–200. [Google Scholar] [CrossRef]
- Lorusso, M.L.; Facoetti, A.; Pesenti, S.; Cattaneo, C.; Molteni, M.; Geiger, G. Wider recognition in peripheral vision common to different subtypes of dyslexia. Vis. Res. 2004, 44, 2413–2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitney, D.; Levi, D.M. Visual crowding: A fundamental limit on conscious perception and object recognition. Trends. Cogn. Sci. 2011, 15, 160–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zorzi, M.; Barbiero, C.; Facoetti, A.; Lonciari, I.; Carrozzi, M.; Montico, M.; Bravar, L.; George, F.; Pech-Georgel, C.; Ziegler, J.C. Extra-large letter spacing improves reading in dyslexia. Proc. Natl. Acad. Sci. USA 2012, 109, 11455–11459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gori, S.; Facoetti, A. How the visual aspects can be crucial in reading acquisition? The intriguing case of crowding and developmental dyslexia. J. Vis. 2015, 14, 15.1.8. [Google Scholar] [CrossRef] [Green Version]
- Strasburger, H. Seven Myths on Crowding and Peripheral Vision. Iperception 2020, 11, 2041669520913052. [Google Scholar] [CrossRef]
- Kirkby, J.A.; Barrington, R.S.; Drieghe, D.; Liversedge, S.P. Parafoveal processing and transposed-letter effects in dyslexic reading. Dyslexia 2022, 28, 359–374. [Google Scholar] [CrossRef]
- Kewan-Khalayly, B.; Migó, M.; Yashar, A. Transient attention equally reduces visual crowding in radial and tangential axes. J. Vis. 2022, 22, 3. [Google Scholar] [CrossRef]
- Werth, R. Rapid improvement of reading performance in children with dyslexia by altering the reading strategy: A novel approach to diagnoses und therapy of reading deficiencies. Restor. Neurol. Neurosci. 2018, 36, 679–691. [Google Scholar] [CrossRef] [Green Version]
- Werth, R. What causes Dyslexia? Identifying the causes and effective compensatory therapy. Restor. Neurol. Neurosci. 2019, 37, 591–608. [Google Scholar] [CrossRef] [Green Version]
- Werth, R. Dyslexic readers improve without training when using a computer-guided reading strategy. Brain Sci. 2021, 11, 526. [Google Scholar] [CrossRef]
- Werth, R. Is developmental dyslexia due to a visual and not a phonological impairment? Brain Sci. 2021, 11, 1313. [Google Scholar] [CrossRef] [PubMed]
- Tallal, P. Auditory temporal perception, phonics, and reading disabilities in children. Brain Lang. 1980, 9, 182–198. [Google Scholar] [CrossRef] [PubMed]
- Tallal, P.; Miller, S.; Fitch, R.H. Neurobiological basis of speech: A case for the pre-eminence of temporal processing. Ann. N. Y. Acad. Sci. 1993, 682, 27–47. [Google Scholar] [CrossRef] [PubMed]
- Merzenich, M.M.; Jenkins, W.M.; Johnston, P.; Schreiner, C.; Miller, S.L.; Tallal, P. Temporal processing deficits of language learning impaired children ameliorated by training. Science 1996, 271, 77–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagarajan, S.; Mahnke, H.; Salz, T.; Tallal, P.; Roberts, T.; Merzenich, M.M. Cortical auditory signal processing in poor readers. Proc. Natl. Acad. Sci. USA 1999, 96, 6483–6488. [Google Scholar] [CrossRef] [Green Version]
- Pavlidis, G.T. Do eye movements hold the key to dyslexia? Neuropsychologia 1981, 19, 57–64. [Google Scholar] [CrossRef]
- Rayner, K.; Pollatsek, A. Eye movement control during reading: Evidence for direct control. Q. J. Exp. Psychol. Sect. A 1982, 33, 351–373. [Google Scholar] [CrossRef]
- Pavlidis, G.T. Eye movements in dyslexia: Their diagnostic significance. J. Learn. Disabil. 1985, 18, 42–50. [Google Scholar] [CrossRef]
- Rayner, K. Do faulty eye movements cause dyslexia? Dev. Neuropsychol. 1985, 1, 3–15. [Google Scholar] [CrossRef]
- Rainer, K. Eye movements and the perceptual span in beginning and skilled readers. J. Exp. Child Psychol. 1986, 41, 211–236. [Google Scholar] [CrossRef]
- Fischer, B.; Biscaldi, M.; Otto, P. Saccadic eye movements of dyslexic adult subjects. Neuropsychologia 1993, 31, 887–906. [Google Scholar] [CrossRef] [PubMed]
- Eden, G.F.F.; Stein, J.F.F.; Wood, H.M.M.; Wood, F.B.B. Differences in eye movements and reading problems in dyslexic and normal children. Vis. Res. 1994, 34, 1345–1358. [Google Scholar] [CrossRef] [PubMed]
- Hyönä, J.; Olson, R.K. Eye fixation patterns among dyslexic and normal readers: Effect of word length and word frequency. J. Exp. Psychol. Learn. Mem. Cogn. 1995, 21, 1430–1440. [Google Scholar] [CrossRef] [PubMed]
- Biscaldi, M.; Gezeck, S.; Stuhr, V. Poor saccadic control correlates with dyslexia. Neuropsychologia 1998, 36, 1189–1202. [Google Scholar] [CrossRef] [PubMed]
- De Luca, M.; Di Pace, E.; Judica, A.; Spinelli, D.; Zoccolotti, P. Eye movement patterns in linguistic and non-linguistic tasks in developmental surface dyslexia. Neuropsychologia 1999, 37, 1407–1420. [Google Scholar] [CrossRef]
- Biscaldi, M.; Fischer, B.; Hartnegg, K. Voluntary saccadic control in dyslexia. Perception 2000, 29, 509–521. [Google Scholar] [CrossRef] [Green Version]
- Rayner, K.; Slattery, T.J.; Bélanger, N.N. Eye movements, the percepzual span, and reading speed. Bull. Rev. 2010, 17, 834–839. [Google Scholar] [CrossRef] [Green Version]
- Ward, L.M.; Kapoula, Z. Dyslexics’ Fragile Oculomotor Control Is Further Destabilized by Increased Text Difficulty. Brain Sci. 2021, 11, 990. [Google Scholar] [CrossRef]
- Temelturk, R.D.; Ozer, E. Binocular coordination of children with dyslexia and typically developing children in linguistic and non-linguistic tasks: Evidence from eye movements. Ann. Dyslexia 2022, 72, 426–444. [Google Scholar] [CrossRef] [PubMed]
- Premeti, A.; Bucci, M.P.; Isel, F. Evidence from ERP and eye movements as markers of language dysfunction in dyslexia. Brain Sci. 2022, 12, 73. [Google Scholar] [CrossRef]
- Caldani, S.; Acquaviva, E.; Moscoso, A.; Peyre, H.; Delorme, R.; Bucci, M.P. Reading performance in children with ADHD: An eye-tracking study. Ann. Dyslexia 2022, 72, 552–565. [Google Scholar] [CrossRef] [PubMed]
- Wagner, R.K.; Torgesen, J.K. The nature of phonological processing and its causal role in the acquisition of reading skills. Psychol. Bull. 1987, 101, 192–212. [Google Scholar] [CrossRef]
- Bruce, D. An analysis of word sounds by young children. Br. J. Educ. Psychol. 1964, 34, 158–170. [Google Scholar] [CrossRef]
- Bryant, P.E.; MacLean, M.; Bradley, L.L.; Crossland, J. Rhyme and alliteration, phoneme detection, and learning to read. Dev. Psychol. 1990, 26, 429–438. [Google Scholar] [CrossRef]
- Torgesen, J.K.; Wagner, R.K.; Rashotte, C.A. Longitudinal studies of phonological processing and reading. J. Learn. Disabil. 1994, 27, 276–286. [Google Scholar] [CrossRef] [PubMed]
- McBride-Chang, C. What is phonological awareness? J. Educ. Psychol. 1995, 87, 179–192. [Google Scholar] [CrossRef]
- Yopp, H.K. A test for assessing phonemic awareness in young children. Read. Teach. 1995, 49, 20–29. [Google Scholar] [CrossRef]
- Torgesen, J.K.; Wagner, R.K.; Rashotte, C.A. Prevention and remediation of severe reading disabilities: Keeping the end in mind. Sci. Stud. Read. 1997, 1, 217–234. [Google Scholar] [CrossRef]
- Muter, V.; Hulme, C.; Snowling, M.J.; Taylor, S. Segmentation, not rhyming, predicts early progress in learning to read. J. Exp. Child. Psychol. 1998, 71, 3–27. [Google Scholar] [CrossRef]
- Ehri, L.C. Research on learning to read and spell: A personal historical perspective. Sci. Stud. Read. 1998, 2, 97–114. [Google Scholar] [CrossRef]
- Snowling, M. From language to reading and dyslexia. Dyslexia. Dyslexia 2001, 7, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Goswami, U. Phonology, reading development, and dyslexia: A cross-linguistic perspective. Ann. Dyslexia 2002, 52, 139–163. [Google Scholar] [CrossRef]
- Vellutino, F.R.; Fletcher, J.M.; Snowling, M.J. Scanlon, D.M. Specific reading disability (dyslexia): What have we learned in the past four decades. J. Child Psychol. Psychiat. 2004, 45, 2–40. [Google Scholar] [CrossRef] [PubMed]
- Goswami, U. Phonology, reading and reading difficulties. In Interdisciplinary Perspectives on Learning to Read. Culture, Cognition and Pedagogy; Hall, K., Goswami, U., Harrison, C., Ellis, S., Soler, Eds.; Routledge: London, UK, 2010; pp. 103–116. [Google Scholar]
- Ligges, C.; Lehmann, T. Multiple case study in German children with dyslexia: Characterization of phonological, auditory, visual, and cerebellar processing on the group and individual level. Brain Sci. 2022, 12, 1292. [Google Scholar] [CrossRef] [PubMed]
- Mackie, J.L. Causes and conditions. Am. Philos. Q. 1965, 2, 245–264. [Google Scholar]
- Spirtes, P.; Glymour, C.; Scheines, R. Causation, Prediction, and Search; MIT Press: Cambridge, UK, 1992. [Google Scholar]
- Lewis, D. Causation as influence. J. Philos. 2000, 97, 182–197. [Google Scholar] [CrossRef] [Green Version]
- Spohn, W. Causation: An alternative. Brit. J. Philos. Sci. 2006, 57, 93–119. [Google Scholar] [CrossRef] [Green Version]
- Pearl, J.; Glymour, M.; Jewell, N.P. Causal Inference in Statistics; Wiley: Chichester, UK, 2016. [Google Scholar]
- Pearl, J. Causality—Models, Reasoning and Inference; Cambridge University Press: Cambridge, UK; New York, NY, USA; Port Melbourne, Australia, 2018; pp. 316–320. [Google Scholar]
- Bressler, S.L.; Seth, A.K. Wiener-Granger causality: A well established methodology. Neuroimage 2011, 58, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Stokes, P.A.; Purdon, P.L. A study of problems encountered in Granger causality analysis from a neuroscience perspective. Proc. Natl. Acad. Sci. USA 2017, 114, E7063–E7072. [Google Scholar] [CrossRef] [Green Version]
- Ingthorson, R. The Logical vs. the Ontological Understanding of Conditions. Metaphysica 2008, 9, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Douven, I. The Evidential Support Theory of Conditionals. Synthese 2008, 164, 19–44. [Google Scholar] [CrossRef]
- Gomes, G. Are Necessary and Sufficient Conditions Converse Relations? Australas. J. Philos. 2009, 87, 375–387. [Google Scholar] [CrossRef]
- Gomes, G. Meaning Preserving Contraposition of Natural Language Conditionals. J. Prag. 2019, 152, 46–60. [Google Scholar] [CrossRef]
- Gomes, G. Concessive conditionals without Even if and non-concessive conditionals with Even if. Acta Analyt. 2020, 35, 1–21. [Google Scholar] [CrossRef]
- Crupi, V.; Iacona, A. The Evidential Conditional. Erkenntnis 2020, 87, 2897–2921. [Google Scholar] [CrossRef]
- Raidl, E.; Iacona, A.; Crupi, V. The Logic of the Evidential Conditional. Rev. Symb. Logic 2021, 15, 758–770. [Google Scholar] [CrossRef]
- Crupi, V. On the Logical Form of Concessive Conditionals. J. Philos. Logic 2022, 51, 633–651. [Google Scholar] [CrossRef]
- Farah, M.; Stowe, R.M.; Levinson, K.L. Phonological dyslexia. Loss of a reading specific component of cognitive architecture? Cogn. Neuropsychol. 1996, 13, 849–868. [Google Scholar] [CrossRef]
- Hari, R.; Renvall, H. Impaired processing of rapid stimulus sequences in dyslexia. Trends Cogn. Sci. 2001, 5, 525–532. [Google Scholar] [CrossRef]
- Roach, N.W.; Hogben, J.H. Attentional modulation of visual processing in adult dyslexia: A spatial cuing deficit. Psychol. Sci. 2004, 15, 650–654. [Google Scholar] [CrossRef]
- Buchholz, J.; Davis, A.A. Adults with dyslexia demonstrate space-based and object-based covert attention deficits: Shifting attention to the periphery and shifting attention between objects in the left visual field. Brain Cogn. 2005, 57, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Heim, S.; Tschierse, J.; Amunts, K.; Wilms, M.; Vossel, S.; Willmes, K.; Grabowska, A.; Huber, W. Cognitive subtypes of dyslexia. Acta Neurobiol. Exp. 2008, 68, 73–82. [Google Scholar]
- Daniel, A.; Abrams, D.A.; Nicol, T.; Zecker, S.; Kraus, N. Abnormal cortical processing of the syllable rate of speech in poor readers. J. Neurosci. 2009, 29, 7686–7693. [Google Scholar] [CrossRef] [Green Version]
- Menghini, D.; Finzi, A.; Benassi, M.; Bolzani, R.; Facoetti, A.; Giovagnoli, S.; Ruffino, M.; Vicari, S. Different underlying neurocognitive deficits in developmental dyslexia: A comparative study. Neuropsychologia 2010, 48, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Hornickel, J.; Kraus, N. Unstable representation of sound: A biological marker of dyslexia. J. Neurosci. 2013, 33, 3500–3504.67. [Google Scholar] [CrossRef] [Green Version]
- Lorusso, M.L.; Cantiani, C.; Molteni, M. Age, dyslexia subtype and comorbidity modulate rapid auditory processing in developmental dyslexia. Front. Hum. Neurosci. 2014, 8, 313. [Google Scholar] [CrossRef] [Green Version]
- van Bergen, E.; de Jong, P.F.; Maassen, B.; Krikhaar, E.; Plakas, A.; van der Leij, A. IQ of four-year-olds who go on to develop dyslexia. J. Learn. Disabil. 2014, 47, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Hendren, R.L.; Haft, S.L.; Black, J.M.; White, N.C.; Hoeft, F. Recognizing Psychiatric Comorbidity With Reading Disorders. Front. Psychiatry 2018, 9, 101. [Google Scholar] [CrossRef] [Green Version]
- Peters, L.; Bulthé, J.; Daniels, N.; Op de Beeck, H.; De Smedt, B. Dyscalculia and dyslexia: Different behavioral, yet similar brain activity profiles during arithmetic. Neuroimage Clin. 2018, 18, 663–674. [Google Scholar] [CrossRef]
- Moll, K.; Landerl, K.; Snowling, M.J.; Schulte-Körne, G. Understanding comorbidity of learning disorders: Task-dependent estimates of prevalence. J. Child. Psychol. Psychiat. 2019, 60, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Maziero, S.; Tallet, J.; Bellocchi, S.; Jover, M.; Chaix, Y.; Jucla, M. Influence of comorbidity on working memory profile in dyslexia and developmental coordination disorder. J. Clin. Exp. Neuropsychol. 2020, 42, 660–674. [Google Scholar] [CrossRef] [PubMed]
- Facoetti, A.; Paganoni, P.; Turatto, M.; Marzola, V.; Mascetti, G.G. Visual spatial attention in developmental dyslexia. Cortex 2000, 36, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, M.; Williams, P.E.; Yeshurun, Y. Covert attention increases spatial resolution with or without masks: Support for signal enhancement. J. Vision 2002, 2, 467–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosse, M.L.; Tainturier, M.J.; Valdois, S. Developmental dyslexia: The visual attention span deficit hypothesis. Cognition 2007, 104, 198–230. [Google Scholar] [CrossRef] [Green Version]
- Franceschini, S.; Gori, M.; Ruffino, K.; Pedrolli, A.; Facoetti, A. A causal link between visual spatial attention and reading acquisition. Curr. Biol. 2012, 22, 814–819. [Google Scholar] [CrossRef] [Green Version]
- Ruffino, M.; Gori, S.; Boccardi, D.; Molteni, M.; Facoetti, A. Spatial and temporal attention in developmental dyslexia. Front. in Human Neurosci. 2014, 8, 8–331. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Schneps, M.H.; Masyn, K.E.; Thomson, J.M. The effects of visual attention span and phonological decoding in reading comprehension in dyslexia: A path analysis. Dyslexia 2016, 22, 322–344. [Google Scholar] [CrossRef]
- Facoetti, A.; Turatto, M.; Lorusso, M.L.; Mascetti, G.G. Orienting of visual attention in dyslexia: Evidence for asymmetric hemispheric control of attention. 2001, 138, 46–53. Exp. Brain Res. 2001, 138, 46–53. [Google Scholar] [CrossRef]
- Facoetti, A.; Lorusso, M.L.; Paganoni, P.; Cattaneo, C.; Galli, R.; Umiltà, C.; Mascetti, G.G. Auditory and visual automatic attention deficits in developmental dyslexia. Brain Res. Cogn. Brain Res. 2003, 16, 185–191. [Google Scholar] [CrossRef]
- Roach, N.W.; Hogben, J.H. Spatial cueing deficits in dyslexia reflect generalised difficulties with attentional selection. Vision Res. 2008, 48, 193–207. [Google Scholar] [CrossRef] [Green Version]
- Facoetti, A.; Zorzi, M.; Cestnick, L.; Lorusso, M.L.; Molteni, L.M.; Paganoni, P.; Umilta, C.; Mascetti, G.G. The relationship between visuo-spatial attention and nonword reading in developmental dyslexia. Cogn. Neuropsych. 2006, 23, 841–855. [Google Scholar] [CrossRef] [PubMed]
- Pina Rodrigues, A.; Castelo-Branco, M.; van Asselen, M. Disrupted spatial organization of cued exogenous attention persists into adulthood in developmental dyslexia. Front. Psychol. 2021, 12, 769237. [Google Scholar] [CrossRef]
- Skottun, B.C.; Skoyles, J. Dyslexia: Sensory deficits or inattention? Perception 2007, 36, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Skottun, B.C. On the use of cues to assess attention in dyslexia. Front. Hum. Neurosci. 2014, 8, 983. [Google Scholar] [CrossRef] [Green Version]
- De Luca, M.; Burani, C.; Paizi, D.; Spinelli, D.; Zoccolotti, P. Letter and letter-string processing in developmental dyslexia. Cortex 2010, 46, 1272–1283. [Google Scholar] [CrossRef] [PubMed]
- Zoccolotti, P.; De Luca, M.; Spinelli, D. Discrete versus multiple word displays: A re-analysis of studies comparing dyslexic and typically developing children. Front. Psychol. 2015, 6, 1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burani, C.; Marcolini, S.; Traficante, D.; Zoccolotti, P. Reading derived words by Italian children with and without dyslexia: The effect of root length. Front. Psychol. 2018, 9, 647. [Google Scholar] [CrossRef] [Green Version]
- Franceschini, S.; Bertoni, S.; Puccio, G.; Gori, S.; Termine, C.; Facoetti, A. Visuo-spatial attention deficit in children with reading difficulties. Sci. Rep. 2022, 12, 13930. [Google Scholar] [CrossRef]
- Valdois, S. The visual-attention span deficit in developmental dyslexia: Review of evidence for a visual-attention-based deficit. Dyslexia 2022, 28, 397–415. [Google Scholar] [CrossRef] [PubMed]
- Perry, C.; Long, H. What is going on with visual attention in reading and dyslexia? A critical review of recent studies. Brain Sci. 2022, 12, 87. [Google Scholar] [CrossRef]
- Stenneken, P.; Egetemeir, J.; Schulte-Körne, G.; Müller, H.J.; Schneider, W.X.; Finke, K. Slow perceptual processing at the core of developmental dyslexia: A parameter-based assessment of visual attention. Neuropsychologia 2011, 49, 3454–3465. [Google Scholar] [CrossRef] [PubMed]
- Pammer, K.; Lavis, R.; Hansen, P.; Cornelissen, P.L. Symbol-string sensitivity and children’s reading. Brain Lang. 2004, 89, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Vidyasagar, R.R.; Pammer, K. Dyslexia: A deficit in visuo-spatial attention, not a phonological processing. Trends Cogn. Sci. 2010, 14, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Graham, C.H.; Cook, C. Visual acuity as a function of intensity and exposure-time. Am. J. Psychol. 1937, 49, 654–661. [Google Scholar] [CrossRef]
- Barlow, H.B. Temporal and spatial summation in human vision at different background intensities. J. Physiol. Lond. 1958, 141, 337–350. [Google Scholar] [CrossRef] [Green Version]
- Niwa, K.; Tokoro, T. Measurement of temporal summation of visual acuity with use of modified tachistoscope. Jpn. J. Ophthalmol. 1997, 41, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Sandberg, K.; Bibby, B.M.; Timmermans, B.; Cleeremans, A.; Overgaard, M. Measuring consciousness: Task accuracy and awareness as sigmoid functions of stimulus duration. Conscious. Cogn. 2011, 20, 1659–1675. [Google Scholar] [CrossRef]
- McAnany, J.J. The effect of exposure duration on visual acuity for letter optotypes and gratings. Vision Res. 2014, 105, 86–91. [Google Scholar] [CrossRef] [Green Version]
- Windey, B.; Vermeiren, A.; Atas, A.; Cleeremans, A. The graded and dichotomous nature of visual awareness. Philos. Trans. R.Soc. B. Biol. Sci. 2014, 369, 20130282. [Google Scholar] [CrossRef] [Green Version]
- Mulholland, P.J.; Redmond, T.; Garway-Heath, D.F.; Zlatkova, M.B.; Anderson, R.S. The Effect of Age on the Temporal Summation of Achromatic Perimetric Stimuli. Invest. Ophthalmol. Vis. Sci. 2015, 56, 6467–6472. [Google Scholar] [CrossRef] [Green Version]
- Holmes, R.; Victora, M.; Wang, R.F.; Kwiat, P.G. Measuring temporal summation in visual detection with a single-photon source. Vis. Res. 2017, 140, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Stigliani, A.; Jeska, B.; Grill-Spector, K. Encoding model of temporal processing in human visual cortex. Proc. Natl. Acad. Sc.i USA 2017, 114, E11047–E11056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Benson, N.C.; Kay, K.N. Winawer, J. Compressive temporal summation in human visual cortex. J. Neurosci. 2018, 38, 691–709. [Google Scholar] [CrossRef] [Green Version]
- Beauny, A.; de Heering, A.; Muñoz Moldes, S.; Martin, J.-R.; de Beir, A.; Cleeremans, A. Unconscious categorization of sub-millisecond complex images. PLoS One 2020, 15, e0236467. [Google Scholar] [CrossRef]
- Heinrich, S.P.; Blechenberg, T.; Reichel, C.; Bach, M. The "speed" of acuity in scotopic vs. photopic vision. Graefes Arch. Clin. Exp. Ophthalmol. 2020, 258, 2791–2798. [Google Scholar] [CrossRef] [PubMed]
- Harvey, B.M.; Dumoulin, S.O.; Fracasso, A.; Paul, J.M. A Network of Topographic Maps in Human Association Cortex Hierarchically Transforms Visual Timing-Selective Responses. Curr. Biol. 2020, 30, 1424–1434.e6. [Google Scholar] [CrossRef] [PubMed]
- Hendrikx, E.; Paul, J.M.; van Ackooij, M.; van der Stoep, N.; Harvey, B.M. Visual timing-tuned responses in human association cortices and response dynamics in early visual cortex. Nat. Commun. 2022, 8, 3952. [Google Scholar] [CrossRef]
- Paul, J.M.; van Ackooij, M.; Ten Cate, T.C.; Harvey, B.M. Numerosity tuning in human association cortices and local image contrast representations in early visual cortex. Nat. Commun. 2022, 13, 1340. [Google Scholar] [CrossRef]
- Weiner, K.S.; Sayres, R.; Vinberg, J.; Grill-Spector, K. fMRI-adaptation and category selectivity in human ventral temporal cortex: Regional differences across time scales. J. Neurophysiol. 2010, 103, 3349–3365. [Google Scholar] [CrossRef] [Green Version]
- Fornaciai, M.; Brannon, E.M.; Woldorff, M.G.; Park, J. Numerosity processing in early visual cortex. Neuroimage 2017, 157, 429–438. [Google Scholar] [CrossRef]
- DeWind, N.K.; Park, J.; Woldorff, M.G.; Brannon, E.M. Numerical encoding in early visual cortex. Cortex 2019, 114, 76–89. [Google Scholar] [CrossRef] [PubMed]
- Mechelli, A.; Humphreys, G.W.; Mayall, K.; Olson, A.; Price, C.J. Differential effects of word length and visual contrast in the fusiform and lingual gyri during reading. Proc. Royal Soc. Lond. Series B. 2000, 267, 1909–1913. [Google Scholar] [CrossRef] [PubMed]
- Schurz, M.; Sturm, D.; Richlan, F.; Kronbichler, M.; Ladurner, G.; Wimmer, H. A dual-route perspective on brain activation in response to visual words: Evidence for a length by lexicality interaction in the visual word form area (VWFA). Neuroimage 2010, 49, 2649–2661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bálint, D. Seelenlähmung des “Schauens”, optische Ataxie, räumliche Störung der Aufmerksamkeit. Mschr. Psychiat. Neurol. 1909, 25, 51–81. [Google Scholar] [CrossRef] [Green Version]
- Poppelreuter, W. Die Psychischen Schädigungen durch Kopfschuß im Kriege 1914–1918. Bd. I. Die Störungen der Niederen und Höheren Sehleistungen durch Verletzungen des Okzipitalhirns; L. Voss: Leipzig, Germany, 1917. [Google Scholar]
- Luria, A.R. Disorders of “simultaneous perception” in a case of bilateral occipito-parietal brain injury. Brain 1959, 82, 437–449. [Google Scholar] [CrossRef]
- Baylis, G.C.; Driver, J.; Baylis, L.L.; Rafal, R.D. Reading of letters and words in a patient with Balint’s syndrome. Neuropsychologia 1994, 32, 1273–1286. [Google Scholar] [CrossRef] [PubMed]
- Rizzo., M.; Vecera, S.P. Psychoanatomical substrates of Bálint’s syndrome. J. Neurol. Neurosurg. Psychiatry 2002, 72, 162–178. [Google Scholar] [CrossRef] [Green Version]
- Moreaud, O. Balint syndrome. Arch. Neurol. 2003, 60, 1329–1331. [Google Scholar] [CrossRef]
- Michel, F.; Henaff, M.A. Seeing without the occipito-parietal cortex: Simultagnosia as a shrinkage of the attentional visual field. Behav. Neurol. 2004, 15, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Farah, M.J. Visual Agnosia, 2nd ed.; MIT Press: Cambridge, MA, USA; London, UK, 2004; pp. 43–58. [Google Scholar]
- Chechlacz, M.; Rotshtein, P.; Hansen, P.C.; Riddoch, J.M.; Deb, S.; Humphreys, G.W. The neural underpinings of simultanagnosia: Disconnecting the visuospatial attention network. J. Cogn. Neurosci. 2012, 247, 18–35. [Google Scholar] [CrossRef]
- Chechlacz, M.; Humphreys, G.W. The enigma of Balint’s syndrome: Neural substrates and cognitive deficits. Front. Hum. Neurosci. 2014, 8, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chechlacz, M. Bilateral parietal dysfunctions and disconnections in simultanagnosia and Bálint syndrome. Handb. Clin. Neurol. 2018, 151, 249–267. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.G.; Haslerud, G.M. Influence on extreme peripheral vision of attention to a visual or auditory task. J. Exp. Psychol. 1964, 68, 269–272. [Google Scholar] [CrossRef] [PubMed]
- Engel, F.L. Visual conspicuity, directed attention and retinal locus. Vision Res. 1971, 11, 563–576. [Google Scholar] [CrossRef]
- Ikeda, M.; Takeuchi, T. Influence of foveal load on the functional visual field. Perc. Psychophys. 1975, 18, 225–260. [Google Scholar] [CrossRef]
- Henderson, J.M.; Ferreira, F. Effects of foveal processing difficulty on the perceptual span in reading: Implication for attention and eye movement control. J. Exp. Psychol. Learn. Mem. Cogn. 1990, 16, 417–429. [Google Scholar] [CrossRef]
- Handy, T.C.; Kingstone, A.; Mangun, G.R. Spatial distribution of visual attention: Perceptual sensitivity and response latency. Percept. Psychophys. 1996, 58, 613–627. [Google Scholar] [CrossRef]
- Werth, R. Therapie von Lesestörungen durch Erkennen und Beheben der Ursachen. Ergother. Rehabil. 2006, 9, 6–11. [Google Scholar]
- Klische, A. Leseschwächen gezielt beheben. Individuelle Diagnose und Therapie mit dem Programm Celeco; Tectum: Marburg, Germany, 2007. [Google Scholar]
- Gabay, Y.; Thiessen, E.D.; Holt, L.L. Impaired statistical learning in developmental dyslexia. J. Speech Lang. Hear. Res. 2015, 58, 934–945. [Google Scholar] [CrossRef]
- Adlof, S.M.; Baron, L.S.; Bell, B.A.; Scoggins, J. Spoken Word Learning in Children With Developmental Language Disorder or Dyslexia. J. Speech Lang. Hear. Res. 2021, 64, 2734–2749. [Google Scholar] [CrossRef]
- Alt, M.; Gray, S.; Hogan, T.P.; Schlesinger., N.; Cowan, N. Spoken Word Learning Differences Among Children With Dyslexia, Concomitant Dyslexia and Developmental Language Disorder, and Typical Development. Lang. Speech. Hear. Serv. Sch. 2019, 50, 540–561. [Google Scholar] [CrossRef] [PubMed]
- Melby-Lervåg, M.; Lyster, S.A.; Hulme, C. Phonological skills and their role in learning to read: A meta-analytic review. Psychol. Bull. 2012, 138, 322–352. [Google Scholar] [CrossRef] [PubMed]
- Kimel, E.; Lieder, I.; Ahissar, M. Repeated series learning revisited with a novel prediction on the reduced effect of item frequency in dyslexia. Sci. Rep. 2022, 12, 13521. [Google Scholar] [CrossRef] [PubMed]
- Dębska, A.; Łuniewska, M.; Zubek, J.; Chyl, K.; Dynak, A.; Dzięgiel-Fivet, G.; Plewko, J.; Jednoróg, K.; Grabowska, A. The cognitive basis of dyslexia in school-aged children: A multiple case study in a transparent orthography. Dev. Sci. 2022, 25, e13173. [Google Scholar] [CrossRef] [PubMed]
- van Rijthoven, R.; Kleemans, T.; Segers, E.; Verhoeven, L. Compensatory role of verbal learning and consolidation in reading and spelling of children with dyslexia. Ann. Dyslexia. 2022, 72, 461–486. [Google Scholar] [CrossRef] [PubMed]
- Lazzaro, G.; Varuzza, C.; Costanzo, F.; Fucà, E.; Di Vara, S.; De Matteis, M.E.; Vicari, S.; Menghini, D. Memory Deficits in Children with Developmental Dyslexia: A Reading-Level and Chronological-Age Matched Design. Brain Sci. 2021, 11, 40. [Google Scholar] [CrossRef]
- Torgesen, J.K.; Wagner, R.K.; Rashotte, C.A.; Herron, J.; Lindamood, P. Computer-assisted instruction to prevent early reading difficulties in students at risk for dyslexia: Outcomes from two instructional approaches. Ann. Dyslexia 2010, 60, 40–56. [Google Scholar] [CrossRef] [Green Version]
- Galuschka, K.; Ise, E.; Krick, K.; Schulte-Körne, G. Effectiveness of treatment approaches for children and adolescents with reading disabilities: A meta-analysis of randomized controlled trials. PLoS ONE 2014, 9, e89900. [Google Scholar] [CrossRef] [Green Version]
- Tilanus, E.A.T.; Segers, E.; Verhoeven, L. Predicting responsiveness to a sustained reading and spelling intervention in children with dyslexia. Dyslexia 2019, 25, 190–206. [Google Scholar] [CrossRef]
- Forné, S.; López-Sala, A.; Mateu-Estivill, R.; Adan, A.; Caldú, X.; Rifà-Ros, X.; Serra-Grabulosa, J.M. Improving Reading Skills Using a Computerized Phonological Training Program in Early Readers with Reading Difficulties. Int. J. Environ. Res. Public Health 2022, 19, 11526. [Google Scholar] [CrossRef]
- Hughes, J.A.; Phillips, G.; Reed, P. Brief exposure to a self-paced computer-based reading programme and how it impacts reading ability and behaviour problems. PLoS ONE 2013, 8, e77867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulvermüller, F. The case of CAUSE: Neurobiological mechanisms for grounding an abstract concept. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2018, 373, 20170129. [Google Scholar] [CrossRef] [PubMed]
- Weichwald, S.; Peters, J. Causality in Cognitive Neuroscience: Concepts, Challenges, and Distributional Robustness. J. Cogn. Neurosci. 2021, 33, 226–247. [Google Scholar] [CrossRef] [PubMed]
- Cohen, L.; Dehaene, S.; Naccache, L.; Lehéricy, S.; Dehaene-Lambertz, G.; Hénaff, M.A.; Michel, F. The visual word form area: Spatial and temporal characterization of an initial stage of reading in normal subjects and posterior split-brain patients. Brain 2000, 123, 291–307. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Grafman, J.; Gaillard, W.D.; Ishii, K.; Vega-Bermudez, F.; Pietrini, P.; Reeves-Tyer, P.; DiCamillo, P.; Theodore, W. Conjoint and extended neural networks for the computation of speechcodes: The neural basis of selective impairment in reading words and pseudowords. Cereb. Cortex 2001, 11, 267–277. [Google Scholar] [CrossRef] [Green Version]
- Cohen, L.; Lehericy, S.; Chochon, F.; Lemer, C.; Rivaud, S.; Dehaene, S. Language-specific tuning of visual cortex? Functional properties of the visual word form area. Brain 2002, 125, 1054–1069. [Google Scholar]
- Polk, T.A.; Stallcup, M.; Aguirre, G.K.; Alsop, D.C.; D’Esposito, M.; Detre, J.A.; Farah, M.J. Neural specialization for letter recognition. J. Cogn. Neurosci. 2002, 14, 145–159. [Google Scholar] [CrossRef]
- Dehaene, S.; Le Clec´H., G.; Poline, J.B.; Le Bihan, D.; Cohen, L. The visual word-form area: A prelexical representation of visualwords in the fusiform gyrus. Neuroreport 2002, 13, 321–325. [Google Scholar] [CrossRef]
- Wydell, T.N.; Vuorinen, T.; Helenius, P.; Salmelin, R. Neural correlates of letter-string length and lexicality during reading in a regular orthography. J. Cogn. Neurosci. 2003, 15, 1052–1062. [Google Scholar] [CrossRef] [Green Version]
- McCandliss, B.D.; Cohen, L.; Dehaene, S. The visual word form area: Expertise for reading in the fusiform gyrus. Trends Cogn. Sci. 2003, 7, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Cohen, L.; Martinaud, O.; Lemer, C.; Lehéricy, S.; Samson, Y.; Obadia, M.; Slachevsky, A.; Dehaene, S. Visual word recognition in the left and right hemispheres: Anatomical and functional correlates of peripheral alexias. Cereb. Cortex 2003, 13, 1313–1333. [Google Scholar] [CrossRef] [Green Version]
- Binder, J.R.; Medler, D.A.; Desai, R.; Conant, L.L.; Liebenthal, E. Some neurophysiological constraints on models of word naming. Neuroimage 2005, 27, 677–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binder, J.R.; Medler, D.A.; Westbury, C.F.; Liebenthal, E.; Buchanan, L. Tuning of the human left fusiform gyrus to sublexical orthographic structure. Neuroimage 2006, 33, 739–748. [Google Scholar] [CrossRef] [Green Version]
- Martens, V.E.; de Jong, P.F. The effect of word length on lexical decision in dyslexic and normal reading children. Brain Lang. 2006, 98, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Kronbichler, M.; Bergmann, J.; Hutzler, F.; Staffen, W.; Mair, A.; Ladurner, G.; Wimmer, H. Taxi vs. Taksi: On orthographic word recognition in the left ventral occitiotemporal cortex. J. Cogn. Neurosci. 2007, 19, 1584–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, C.I.; Liu, J.; Wald, L.L.; Kwong, K.K.; Benner, T.; Kanwisher, N. Visual word processing and experiential origins of functional selectivity in human extrastriate cortex. Proc. Natl. Acad. Sci. USA 2007, 104, 9087–9092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epelbaum, S.; Pinel, P.; Gaillard, R.; Delmaire, C.; Perrin, M.; Dupont, S.; Dehaene, S.; Cohen, L. Pure alexia as a disconnection syndrome: New diffusion imaging evidence for an old concept. Cortex 2008, 44, 962–974. [Google Scholar] [CrossRef]
- Graves, W.W.; Desai, R.; Humphries, C.; Seidenberg, M.S.; Binder, J.R. Neural systems for reading aloud: A multiparametric approach. Cereb. Cortex 2010, 20, 1799–1815. [Google Scholar] [CrossRef] [Green Version]
- Church, J.A.; Balota, D.A.; Petersen, S.E.; Schlaggar, B.L. Manipulation of length and lexicality localizes the functional neuroanatomy of phonological processing in adult readers. J. Cogn. Neurosci. 2011, 23, 1475–1493. [Google Scholar] [CrossRef] [Green Version]
- Ben-Shachar, M.; Dougherty, R.F.; Deutsch, G.K.; Wandell, B.A. The development of cortical sensitivity to visual word forms. J. Cogn. Neurosci. 2011, 23, 2387–2399. [Google Scholar] [CrossRef] [Green Version]
- Stigliani, A.; Weiner, K.S.; Grill-Spector, K. Temporal Processing Capacity in High-Level Visual Cortex Is Domain Specific. J. Neurosci. 2015, 35, 12412–12424. [Google Scholar] [CrossRef] [PubMed]
- Stevens, W.D.; Kravitz, D.J.; Peng, C.S.; Tessler, M.H.; Martin, A. Privileged Functional Connectivity between the Visual Word Form Area and the Language System. J. Neurosci. 2017, 37, 5288–5297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer-Baum, S.; Kook, J.H.; Lee, Y.; Ramos-Nuñez, A.; Vannucci, M. Individual Differences in the Neural and Cognitive Mechanisms of Single Word Reading. Front. Hum. Neurosci. 2018, 12, 271. [Google Scholar] [CrossRef] [PubMed]
- Dębska, A.; Chyl, K.; Dzięgiel, G.; Kacprzak, A.; Łuniewska, M.; Plewko, J.; Marchewka, A.; Grabowska, A.; Jednoróg, K. Reading and spelling skills are differentially related to phonological processing: Behavioral and fMRI study. Dev. Cogn. Neurosci. 2019, 39, 100683. [Google Scholar] [CrossRef] [PubMed]
- Ostertag, C.; Reynolds, J.E.; Dewey, D.; Landman, B.; Huo, Y.; Lebel, C. Altered gray matter development in pre-reading children with a family history of reading disorder. Dev Sci. 2022, 25, e13160. [Google Scholar] [CrossRef]
- Eckert, M.A.; Vaden, K.I., Jr.; Iuricich, F. Dyslexia Data Dyslexia Data Consortium. Cortical asymmetries at different spatial hierarchies relate to phonological processing ability. PLoS Biol. 2022, 20, e3001591. [Google Scholar] [CrossRef]
- Hedenius, M.; Persson, J. Neural correlates of sequence learning in children with developmental dyslexia. Hum. Brain Mapp. 2022, 43, 3559–3576. [Google Scholar] [CrossRef]
- Yu, X.; Ferradal, S.; Dunstan, J.; Carruthers, C.; Sanfilippo, J.; Zuk, J.; Zöllei, L.; Gagoski, B.; Ou, Y.; Grant, P.E.; et al. Patterns of Neural Functional Connectivity in Infants at Familial Risk of Developmental Dyslexia. JAMA Netw. Open 2022, 5, e2236102. [Google Scholar] [CrossRef]
- Zhao, J.; Song, Z.; Zhao, Y.; Thiebaut de Schotten, M.; Altarelli, I.; Ramus, F. White matter connectivity in uncinate fasciculus accounts for visual attention span in developmental dyslexia. Neuropsychologia 2022, 177, 108414. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Peng, P. Reading real words versus pseudowords: A meta-analysis of research in developmental dyslexia. Dev. Psychol. 2022, 58, 1035–1050. [Google Scholar] [CrossRef] [PubMed]
- Braid, J.; Richlan, F. The Functional Neuroanatomy of Reading Intervention. Front. Neurosci. 2022, 16, 921931. [Google Scholar] [CrossRef] [PubMed]
- Logothetis, N.K. The neural basis of the blood-oxygen-level-dependent functional magnetic resonance imaging signal. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2002, 357, 1003–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goense, J.B.; Logothetis, N.K. Neurophysiology of the BOLD fMRI signal in awake monkeys. Curr. Biol. 2008, 18, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Werth, R. Visual functions without the occipital lobe or after cerebral hemispherectomy in infancy. Eur. J. Neurosci. 2006, 24, 2932–2944. [Google Scholar] [CrossRef] [PubMed]
- Werth, R. Cerebral blindness and plasticity oft he visual system in children. A review of visual capacities in patients with occipital lesions, hemispherectomy or hydranencephaly. Rest. Neurol. Neurosci. 2008, 26, 377–389. [Google Scholar]
- Seghier, M.L.; Neufeld, N.H.; Zeidman, P.; Leff, A.P.; Mechelli, A.; Nagendran, A.; Riddoch, J.M.; Humphreys, G.W.; Price, C.J. Reading without the left ventral occipito-temporal cortex. Neuropsychologia 2012, 50, 3621–3635. [Google Scholar] [CrossRef] [Green Version]
- Krafnick, A.J.; Napoliello, E.M.; Flowers, D.L.; Eden, G.F. The role of brain activity in characterizing successful reading intervention in children with dyslexia. Front. Neurosci. 2022, 16, 898661. [Google Scholar] [CrossRef]
- Warrington, E.K.; Shallice, T. Word-form dyslexia. Brain 1980, 103, 99–112. [Google Scholar] [CrossRef]
- Salmelin, R.; Service, E.; Kiesilä, P.; Uutela, K.; Salonen, O. Impaired visual word processing in dyslexia revealed with magnetoencephalography. Ann. Neurol. 1996, 40, 157–162. [Google Scholar] [CrossRef]
- Rumsey, J.M.; Horwitz, B.; Donohue, B.C.; Nace, K.; Maisog, J.M.; Andreason, P. Phonological and orthographic components of word recognition. A PET-rCBF study. Brain 1997, 120, 739–759. [Google Scholar] [CrossRef] [Green Version]
- Tarkiainen, A.; Helenius, P.; Hansen, P.C.; Cornelissen, P.L.; Salmelin, R. Dynamics of letter string perception in the human occipitotemporal cortex. Brain 1999, 122, 2119–2131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulesu, E.; Démonet, J.F.; Fazio, F.; McCrory, E.; Chanoine, V.; Brunswick, N.; Cappa, S.F.; Cossu, G.; Habib, M.; Frith, C.D.; et al. Dyslexia: Cultural diversity and biological unity. Science 2001, 291, 2165–2167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaywitz, B.A.; Shaywitz, S.E.; Pugh, K.R.; Mencl, W.E.; Fulbright, R.K.; Skudlarski, P.; Constable, R.T.; Marchione, K.E.; Fletcher, J.M.; Lyon, G.R.; et al. Disruption of posterior brain systems for reading in children with developmental dyslexia. Biol. Psychiatry 2002, 52, 101–110. [Google Scholar] [CrossRef]
- Polk, T.A.; Farah, M.J. Functional MRI evidence for an abstract, not perceptual, word-form area. J. Exp. Psychol. Gen. 2002, 131, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Cohen, L.; Dehaene, S. Specialization within the ventral stream: The case for the visual word form area. Neuroimage 2004, 22, 466–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mechelli, A.; Crinion, J.T.; Long, S.; Friston, K.J.; Ralph, M.A.L.; Patterson, K.; McClelland, J.L.; Price, C.J. Dissociating reading processes on the basis of neuronal interactions. J. Cogn. Neurosci. 2005, 17, 1753–1765. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.; Bitan, T.; Chou, T.; Burman, D.D.; Booth, J.R. Deficient orthographic and phonological representations in children with dyslexia revealed by brain activation patterns. J. Child Psychol. Psychiatry 2006, 47, 1041–1050. [Google Scholar] [CrossRef] [Green Version]
- Maurer, U.; Brem, S.; Bucher, K.; Kranz, F.; Benz, R.; Steinhausen, H.C.; Brandeis, D. Impaired tuning of a fast occipito-temporal response for print in dyslexic children learning to read. Brain 2007, 130, 3200–3210. [Google Scholar] [CrossRef]
- Ben-Shachar, M.; Dougherty, R.F.; Deutsch, G.K.; Wandell, B.A. Differential sensitivity to words and shapes in ventral occipito-temporal cortex. Cereb. Cortex 2007, 17, 1604–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maisog, J.M.; Einbinder, E.R.; Flowers, D.L.; Turkeltaub, P.E.; Eden, G.F. A meta-analysis of functional neuroimaging studies of dyslexia. Ann. N. Y. Acad. Sci. 2008, 1145, 237–259. [Google Scholar] [CrossRef]
- Glezer, L.S.; Jiang, X.; Riesenhuber, M. Evidence for highly selective neuronal tuning to whole words in the "visual word form area". Neuron 2009, 62, 199–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulesu, E.; Danelli, L.; Berlingeri, M. Reading the dyslexic brain: Multiple dysfunctional routes revealed by a new meta-analysis of PET and fMRI activation studies. Front. Hum. Neurosci. 2014, 8, 830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olulade, O.A.; Flowers, D.L.; Napoliello, E.M.; Eden, G.F. Dyslexic children lack word selectivity gradients in occipito-temporal and inferior frontal cortex. Neuroimage Clin. 2015, 7, 742–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boros, M.; Anton, J.-L.; Pech, C.; Grainger, J.; Szwed, M.; Ziegler, J.C. Orthographic processing deficits in developmental dyslexia: Beyond the ventral visual stream. Neuroimage 2016, 128, 316–327. [Google Scholar] [CrossRef]
- Saygin, Z.M.; Osher, D.E.; Norton, E.S.; Youssoufian, D.A.; Beach, S.D.; Feather, J.; Gaab, N.; Gabrieli, J.D.; Kanwisher, N. Connectivity precedes function in the development of the visual word form area. Nat. Neurosci 2016, 19, 1250–1255. [Google Scholar] [CrossRef] [Green Version]
- Kronbichler, L.; Kronbichler, M. The importance of the left occipitotemporal cortex in developmental dyslexia. Curr. Dev. Disord. Rep. 2018, 5, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, A.P.; Rebola, J.; Pereira, M.; van Asselen, M.; Castelo-Branco, M. Neural Responses of the AnteriorVentral Occipitotemporal Cortex in Developmental Dyslexia: Beyond the Visual Word Form Area. IOVS Vis. Neurosci. 2019, 60, 1063–1068. [Google Scholar]
- Dickens, J.V.; Fama, M.E.; DeMarco, A.T.; Lacey, E.H.; Friedman, R.B.; Turkeltaub, P.E. Localization of Phonological and Semantic Contributions to Reading. J. Neurosci. 2019, 39, 5361–5368. [Google Scholar] [CrossRef] [Green Version]
- Brem, S.; Maurer, U.; Kronbichler, M.; Schurz, M.; Richlan, F.; Blau, V.; Reithler, J.; van der Mark, S.; Schulz, E.; Bucher, K.; et al. Visual word form processing deficits driven by severity of reading impairments in children with developmental dyslexia. Sci. Rep. 2020, 10, 18728. [Google Scholar] [CrossRef]
- Liu, T.; Thiebaut de Schotten, M.; Altarelli, I.; Ramus, F.; Zhao, J. Neural dissociation of visual attention span and phonological deficits in developmental dyslexia: A hub-based white matter network analysis. Hum. Brain. Mapp. 2022, 43, 5210–5219. [Google Scholar] [CrossRef]
- Taran, N.; Farah, R.; DiFrancesco, M.; Altaye, M.; Vannest, J.; Holland, S.; Rosch, K.; Schlaggar, B.L.; Horowitz-Kraus, T. The role of visual attention in dyslexia: Behavioral and neurobiological evidence. Hum. Brain Mapp. 2022, 43, 1720–1737. [Google Scholar] [CrossRef] [PubMed]
- Gaillard, R.; Naccache, L.; Pinel, P.; Clémenceau, S.; Volle, E.; Hasboun, D.; Dupont, S.; Baulac, M.; Dehaene, S.; Adam, C.; et al. Direct intracranial, FMRI, and lesion evidence for the causal role of left inferotemporal cortex in reading. Neuron 2006, 50, 191–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Werth, R. Dyslexia: Causes and Concomitant Impairments. Brain Sci. 2023, 13, 472. https://doi.org/10.3390/brainsci13030472
Werth R. Dyslexia: Causes and Concomitant Impairments. Brain Sciences. 2023; 13(3):472. https://doi.org/10.3390/brainsci13030472
Chicago/Turabian StyleWerth, Reinhard. 2023. "Dyslexia: Causes and Concomitant Impairments" Brain Sciences 13, no. 3: 472. https://doi.org/10.3390/brainsci13030472
APA StyleWerth, R. (2023). Dyslexia: Causes and Concomitant Impairments. Brain Sciences, 13(3), 472. https://doi.org/10.3390/brainsci13030472