
Neural Correlates of Antisocial Behavior: The Victim’s Perspective

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
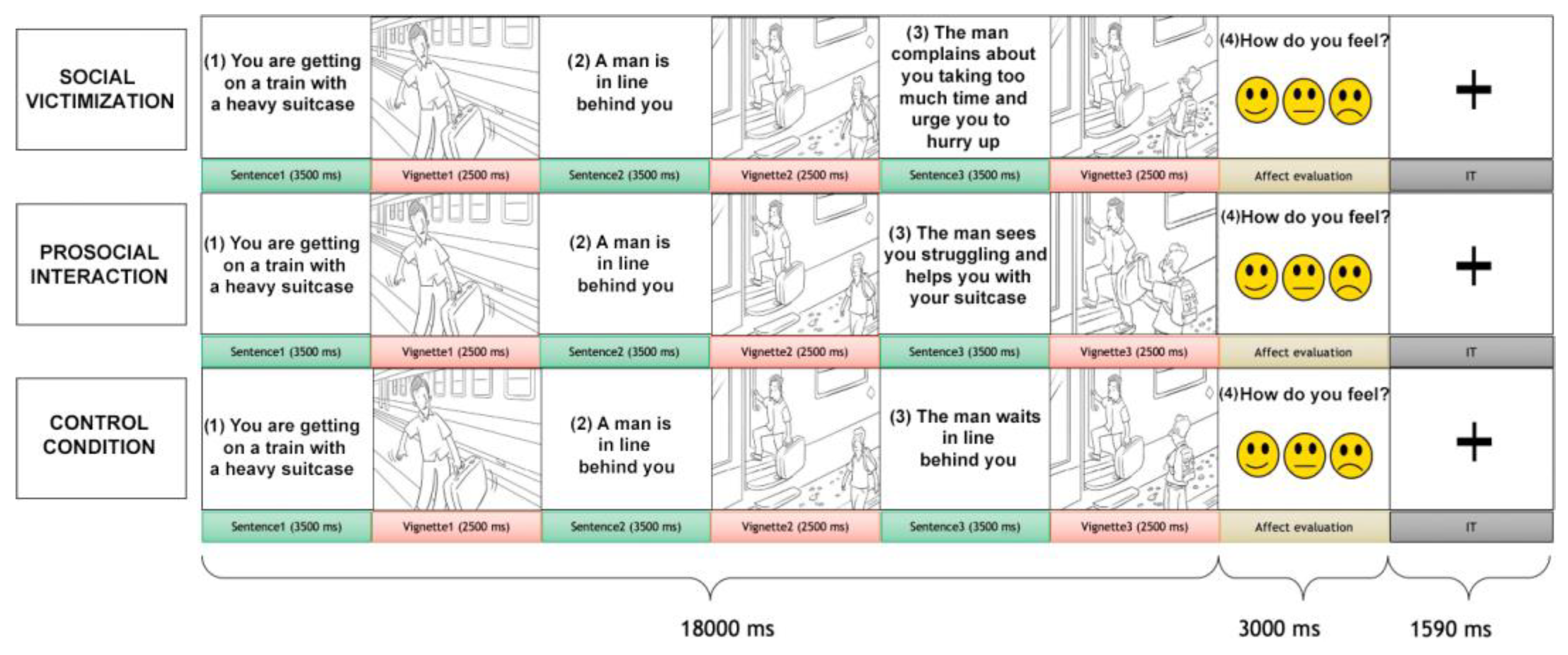
2.2. Social Interaction Task
2.3. Psychological Evaluation
2.4. Image Acquisition and Analysis
2.5. Behavioral and Brain–Behavior Correlation Analyses
3. Results
3.1. Behavioral Results
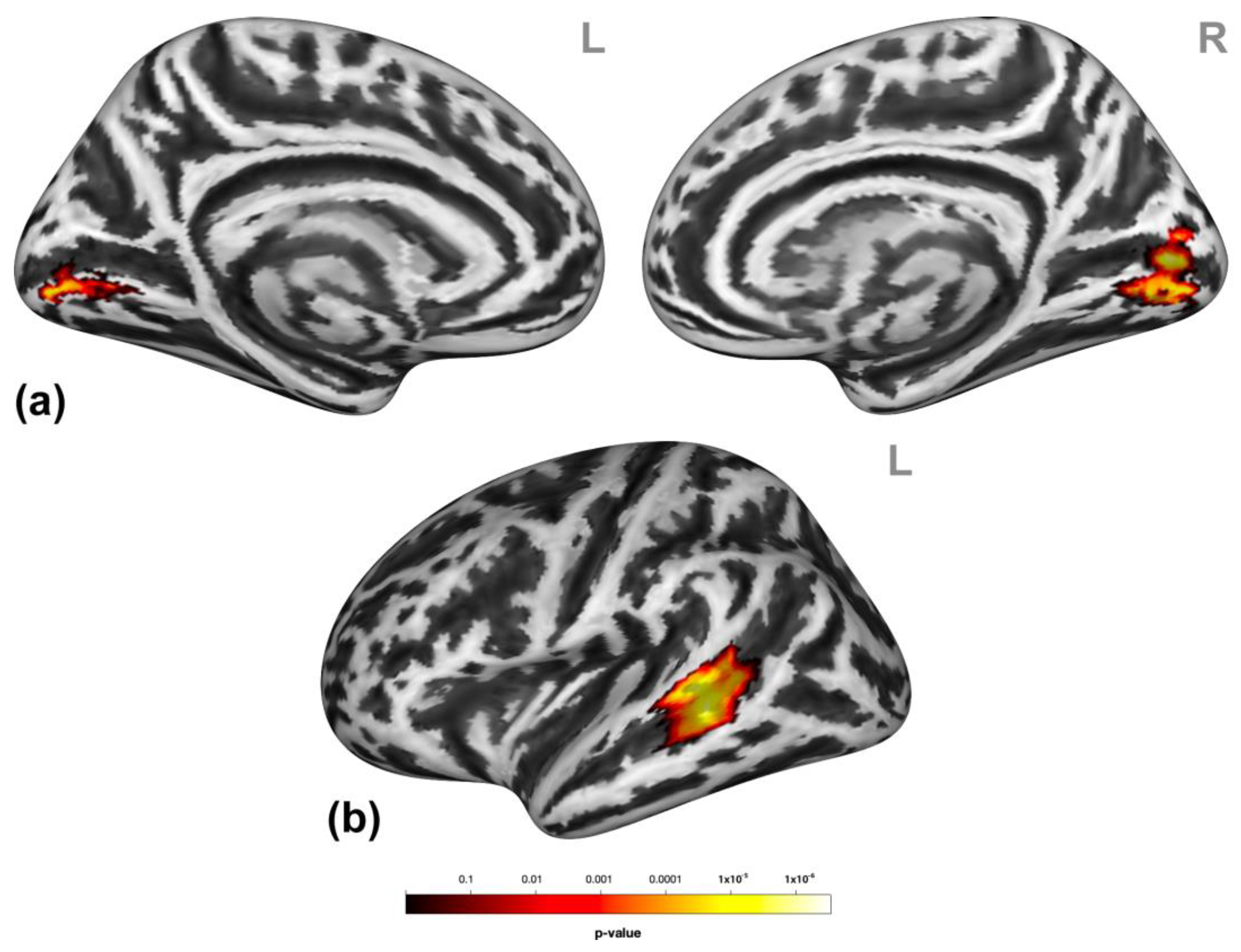
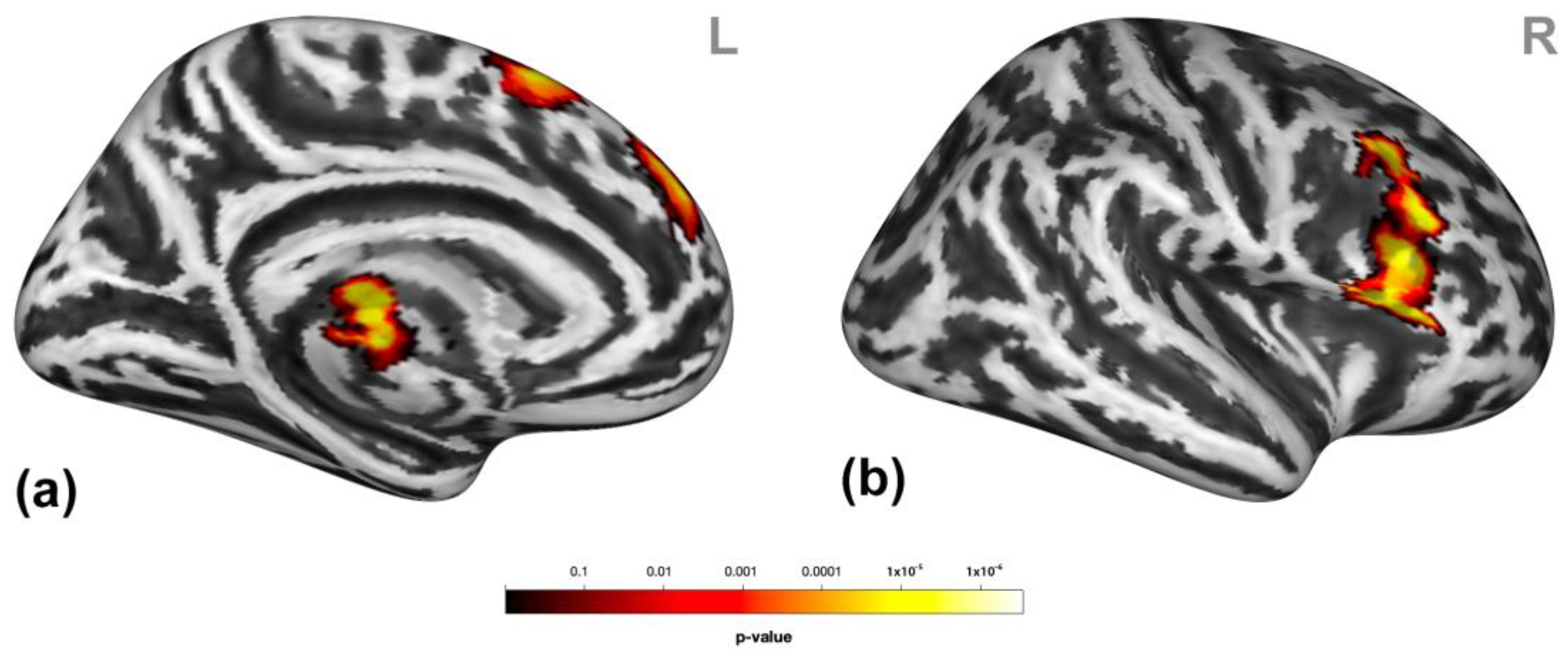
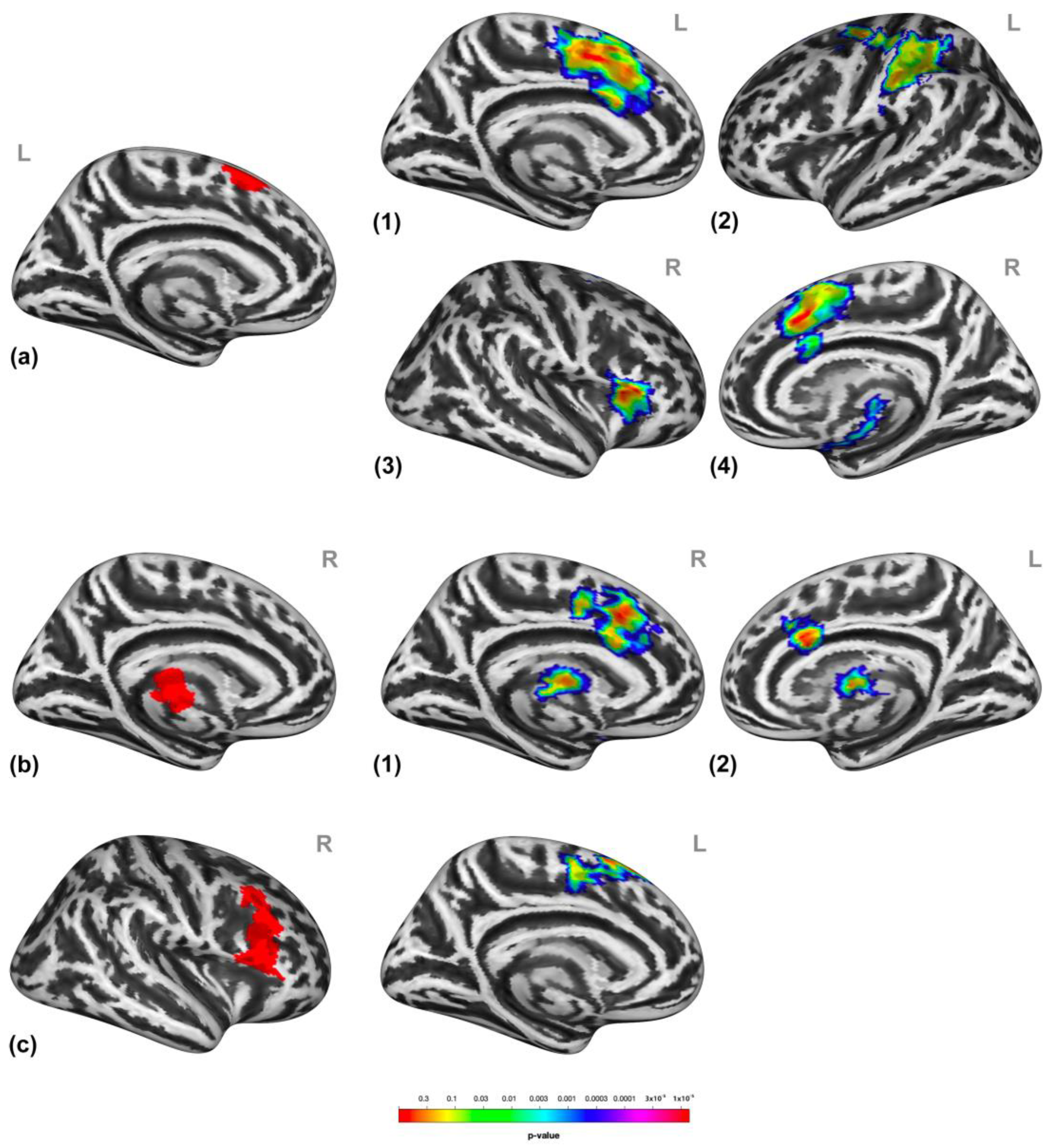
3.2. Brain Activation
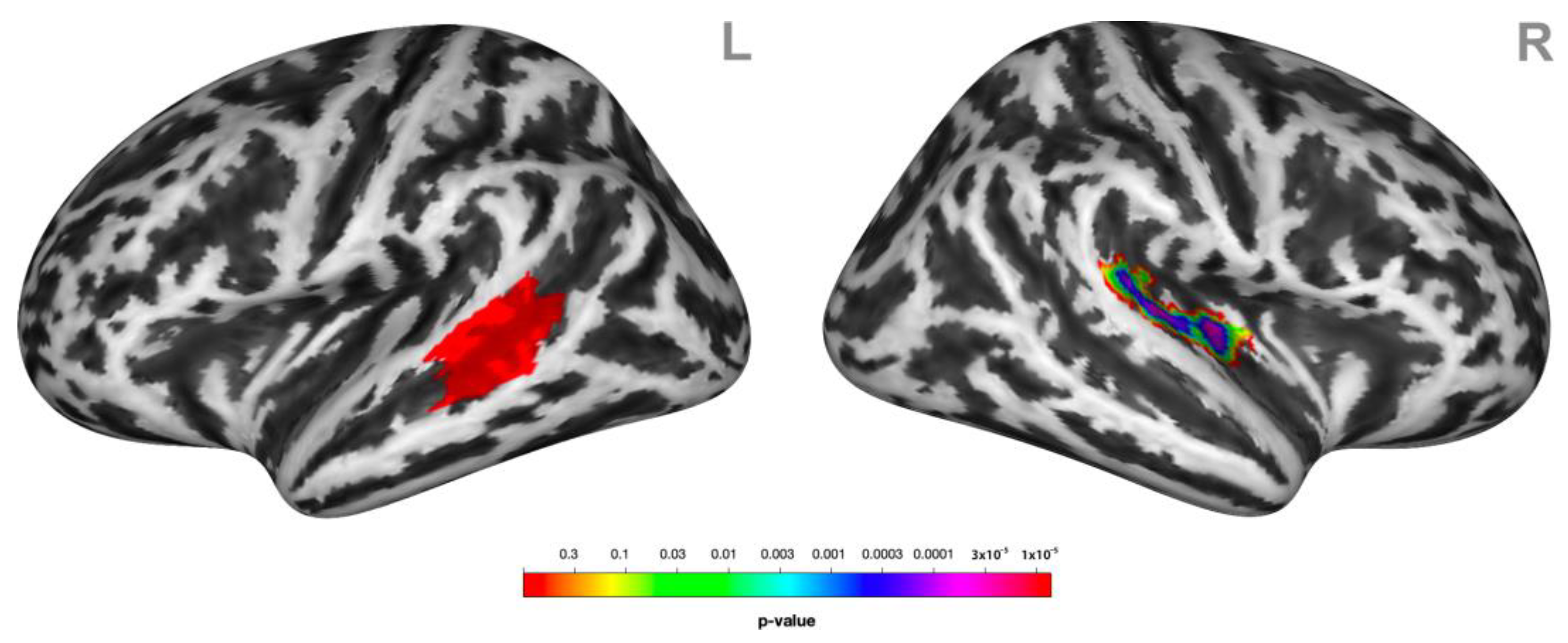
3.3. Psychophysiological Interactions
3.4. Correlations with Psychological Scales
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coan, J.A.; Sbarra, D.A. Social Baseline Theory: The Social Regulation of Risk and Effort. Curr. Opin. Psychol. 2015, 1, 87–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bar-Kalifa, E.; Rafaeli, E. Above and below Baselines: The Nonmonotonic Effects of Dyadic Emotional Support in Daily Life. J. Soc. Pers. Relatsh. 2015, 32, 161–179. [Google Scholar] [CrossRef]
- Calkins, S.D.; Keane, S.P. Developmental Origins of Early Antisocial Behavior. Dev. Psychopathol. 2009, 21, 1095–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, K.D.; Nida, S.A. Ostracism and Social Exclusion: Implications for Separation, Social Isolation, and Loss. Curr. Opin. Psychol. 2022, 47, 101353. [Google Scholar] [CrossRef]
- Leary, M.R. Responses to Social Exclusion: Social Anxiety, Jealousy, Loneliness, Depression, and Low Self-Esteem. J. Soc. Clin. Psychol. 1990, 9, 221–229. [Google Scholar] [CrossRef]
- Mikula, G.; Scherer, K.R.; Athenstaedt, U. The Role of Injustice in the Elicitation of Differential Emotional Reactions. Pers. Soc. Psychol. Bull. 1998, 24, 769–783. [Google Scholar] [CrossRef] [Green Version]
- Miller, D.T. Disrespect and the Experience of Injustice. Annu. Rev. Psychol. 2001, 52, 527–553. [Google Scholar] [CrossRef] [Green Version]
- Iyer-Eimerbrink, P.A.; Scielzo, S.A.; Jensen-Campbell, L.A. The Impact of Peer Victimization. J. Bullying Soc. Aggress. 2015, 1, 1. [Google Scholar]
- Williams, K.D. Chapter 6 Ostracism. In Advances in Experimental Social Psychology; Elsevier: Amsterdam, The Netherlands, 2009; Volume 41, pp. 275–314. ISBN 978-0-12-374472-2. [Google Scholar]
- Williams, K.D.; Cheung, C.K.; Choi, W. Cyberostracism: Effects of Being Ignored over the Internet. J. Pers. Soc. Psychol. 2000, 79, 748–762. [Google Scholar] [CrossRef]
- Zadro, L.; Williams, K.D.; Richardson, R. How Low Can You Go? Ostracism by a Computer Is Sufficient to Lower Self-Reported Levels of Belonging, Control, Self-Esteem, and Meaningful Existence. J. Exp. Soc. Psychol. 2004, 40, 560–567. [Google Scholar] [CrossRef]
- Cloninger, C.R.; Cloninger, K.M.; Zwir, I.; Keltikangas-Järvinen, L. The Complex Genetics and Biology of Human Temperament: A Review of Traditional Concepts in Relation to New Molecular Findings. Transl. Psychiatry 2019, 9, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwir, I.; Del-Val, C.; Arnedo, J.; Pulkki-Råback, L.; Konte, B.; Yang, S.S.; Romero-Zaliz, R.; Hintsanen, M.; Cloninger, K.M.; Garcia, D.; et al. Three Genetic–Environmental Networks for Human Personality. Mol. Psychiatry 2021, 26, 3858–3875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirth, J.H.; Lynam, D.R.; Williams, K.D. When Social Pain Is Not Automatic: Personality Disorder Traits Buffer Ostracism’s Immediate Negative Impact. J. Res. Personal. 2010, 44, 397–401. [Google Scholar] [CrossRef]
- Russo, M.; Mahon, K.; Shanahan, M.; Ramjas, E.; Solon, C.; Braga, R.J.; Burdick, K.E. Affective Temperaments and Neurocognitive Functioning in Bipolar Disorder. J. Affect. Disord. 2014, 169, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Walsh, M.A.; Brown, L.H.; Barrantes-Vidal, N.; Kwapil, T.R. The Expression of Affective Temperaments in Daily Life. J. Affect. Disord. 2013, 145, 179–186. [Google Scholar] [CrossRef]
- Deveney, C.M.; Stoddard, J.; Evans, R.L.; Chavez, G.; Harney, M.; Wulff, R.A. On Defining Irritability and Its Relationship to Affective Traits and Social Interpretations. Personal. Individ. Differ. 2019, 144, 61–67. [Google Scholar] [CrossRef]
- Olivo, D.; Di Ciano, A.; Mauro, J.; Giudetti, L.; Pampallona, A.; Kubera, K.M.; Hirjak, D.; Wolf, R.C.; Sambataro, F. Neural Responses of Benefiting from the Prosocial Exchange: The Effect of Helping Behavior. Front. Psychol 2021, 12, 606858. [Google Scholar] [CrossRef]
- Cloutier, J.; Gabrieli, J.D.E.; O’Young, D.; Ambady, N. An FMRI Study of Violations of Social Expectations: When People Are Not Who We Expect Them to Be. Neuroimage 2011, 57, 583–588. [Google Scholar] [CrossRef] [Green Version]
- Eisenberger, N.I. Social Pain and the Brain: Controversies, Questions, and Where to Go from Here. Annu. Rev. Psychol. 2015, 66, 601–629. [Google Scholar] [CrossRef] [Green Version]
- Bland, A.R.; Zahn, R.; Elliott, R.; Taylor, J.R.; Hill, J. Patrolling the Boundaries of Social Domains: Neural Activations to Violations of Expectations for Romantic and Work Relationships. Soc. Neurosci. 2021, 16, 513–521. [Google Scholar] [CrossRef]
- Knutson, K.M.; Krueger, F.; Koenigs, M.; Hawley, A.; Escobedo, J.R.; Vasudeva, V.; Adolphs, R.; Grafman, J. Behavioral Norms for Condensed Moral Vignettes. Soc. Cogn. Affect. Neurosci. 2010, 5, 378–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyes, M.E.; French, D.J. Having a Cyberball: Using a Ball-Throwing Game as an Experimental Social Stressor to Examine the Relationship between Neuroticism and Coping. Personal. Individ. Differ. 2009, 47, 396–401. [Google Scholar] [CrossRef]
- Caprara, G.V.; Barbaranelli, C.; Borgogni, L.; Perugini, M. The “Big Five Questionnaire”: A New Questionnaire to Assess the Five Factor Model. Personal. Individ. Differ. 1993, 15, 281–288. [Google Scholar] [CrossRef]
- Fico, G.; Luciano, M.; Sampogna, G.; Zinno, F.; Steardo, L.; Perugi, G.; Pompili, M.; Tortorella, A.; Volpe, U.; Fiorillo, A.; et al. Validation of the Brief TEMPS-M Temperament Questionnaire in a Clinical Italian Sample of Bipolar and Cyclothymic Patients. J. Affect. Disord. 2020, 260, 458–462. [Google Scholar] [CrossRef]
- Sebastian, C.; Viding, E.; Williams, K.D.; Blakemore, S.-J. Social Brain Development and the Affective Consequences of Ostracism in Adolescence. Brain Cogn. 2010, 72, 134–145. [Google Scholar] [CrossRef]
- Baars, M.Y.; Müller, M.J.; Gallhofer, B.; Netter, P. Depressive and Aggressive Responses to Frustration: Development of a Questionnaire and Its Validation in a Sample of Male Alcoholics. Depress. Res. Treat. 2011, 2011, 352048. [Google Scholar] [CrossRef]
- Schulz, C.C.; von Klitzing, K.; Deserno, L.; Sheridan, M.A.; Crowley, M.J.; Schoett, M.J.S.; Hoffmann, F.; Villringer, A.; Vrtička, P.; White, L.O. Emotional Maltreatment and Neglect Impact Neural Activation upon Exclusion in Early and Mid-Adolescence: An Event-Related FMRI Study. Dev. Psychopathol. 2022, 34, 573–585. [Google Scholar] [CrossRef]
- Carrington, S.J.; Bailey, A.J. Are There Theory of Mind Regions in the Brain? A Review of the Neuroimaging Literature. Hum. Brain Mapp. 2009, 30, 2313–2335. [Google Scholar] [CrossRef]
- de Greck, M.; Wang, G.; Yang, X.; Wang, X.; Northoff, G.; Han, S. Neural Substrates Underlying Intentional Empathy. Soc. Cogn. Affect. Neurosci. 2012, 7, 135–144. [Google Scholar] [CrossRef]
- Kogler, L.; Müller, V.I.; Chang, A.; Eickhoff, S.B.; Fox, P.T.; Gur, R.C.; Derntl, B. Psychosocial versus Physiological Stress—Meta-Analyses on Deactivations and Activations of the Neural Correlates of Stress Reactions. NeuroImage 2015, 119, 235–251. [Google Scholar] [CrossRef] [Green Version]
- Mwilambwe-Tshilobo, L.; Spreng, R.N. Social Exclusion Reliably Engages the Default Network: A Meta-Analysis of Cyberball. Neuroimage 2021, 227, 117666. [Google Scholar] [CrossRef] [PubMed]
- Zheltyakova, M.; Korotkov, A.; Masharipov, R.; Myznikov, A.; Didur, M.; Cherednichenko, D.; Wagels, L.; Habel, U.; Kireev, M.; Votinov, M. Social Interaction with an Anonymous Opponent Requires Increased Involvement of the Theory of Mind Neural System: An FMRI Study. Front. Behav. Neurosci. 2022, 16, 807599. [Google Scholar] [CrossRef] [PubMed]
- Roxo, M.R.; Franceschini, P.R.; Zubaran, C.; Kleber, F.D.; Sander, J.W. The Limbic System Conception and Its Historical Evolution. Sci. World J. 2011, 11, 2427–2440. [Google Scholar] [CrossRef]
- Britton, J.C.; Phan, K.L.; Taylor, S.F.; Welsh, R.C.; Berridge, K.C.; Liberzon, I. Neural Correlates of Social and Nonsocial Emotions: An FMRI Study. Neuroimage 2006, 31, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Zeki, S.; Romaya, J.P. Neural Correlates of Hate. PLoS ONE 2008, 3, e3556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanderwal, T.; Hunyadi, E.; Grupe, D.W.; Connors, C.M.; Schultz, R.T. Self, Mother and Abstract Other: An FMRI Study of Reflective Social Processing. Neuroimage 2008, 41, 1437–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arioli, M.; Canessa, N. Neural Processing of Social Interaction: Coordinate-Based Meta-Analytic Evidence from Human Neuroimaging Studies. Hum. Brain Mapp. 2019, 40, 3712–3737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochsner, K.N.; Silvers, J.A.; Buhle, J.T. Functional Imaging Studies of Emotion Regulation: A Synthetic Review and Evolving Model of the Cognitive Control of Emotion. Ann. N. Y. Acad. Sci. 2012, 1251, E1–E24. [Google Scholar] [CrossRef] [Green Version]
- Vossel, S.; Geng, J.J.; Fink, G.R. Dorsal and Ventral Attention Systems: Distinct Neural Circuits but Collaborative Roles. Neuroscientist 2014, 20, 150–159. [Google Scholar] [CrossRef]
- Japee, S.; Holiday, K.; Satyshur, M.D.; Mukai, I.; Ungerleider, L.G. A Role of Right Middle Frontal Gyrus in Reorienting of Attention: A Case Study. Front. Syst. Neurosci. 2015, 9, 23. [Google Scholar] [CrossRef] [Green Version]
- Talati, A.; Hirsch, J. Functional Specialization within the Medial Frontal Gyrus for Perceptual Go/No-Go Decisions Based on “What,” “When,” and “Where” Related Information: An FMRI Study. J. Cogn. Neurosci. 2005, 17, 981–993. [Google Scholar] [CrossRef] [PubMed]
- Seok, J.-W.; Bajaj, S.; Soltis-Vaughan, B.; Lerdahl, A.; Garvey, W.; Bohn, A.; Edwards, R.; Kratochvil, C.J.; Blair, J.; Hwang, S. Structural Atrophy of the Right Superior Frontal Gyrus in Adolescents with Severe Irritability. Hum. Brain Mapp. 2021, 42, 4611–4622. [Google Scholar] [CrossRef] [PubMed]
- Nachev, P.; Kennard, C.; Husain, M. Functional Role of the Supplementary and Pre-Supplementary Motor Areas. Nat. Rev. Neurosci. 2008, 9, 856–869. [Google Scholar] [CrossRef] [PubMed]
- Ingvar, M. Pain and Functional Imaging. Philos. Trans. R Soc. Lond. B Biol. Sci. 1999, 354, 1347–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legrain, V.; Iannetti, G.D.; Plaghki, L.; Mouraux, A. The Pain Matrix Reloaded: A Salience Detection System for the Body. Prog. Neurobiol. 2011, 93, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Hampshire, A.; Chamberlain, S.R.; Monti, M.M.; Duncan, J.; Owen, A.M. The Role of the Right Inferior Frontal Gyrus: Inhibition and Attentional Control. Neuroimage 2010, 50, 1313–1319. [Google Scholar] [CrossRef] [Green Version]
- Hughes, M.E.; Johnston, P.J.; Fulham, W.R.; Budd, T.W.; Michie, P.T. Stop-Signal Task Difficulty and the Right Inferior Frontal Gyrus. Behav. Brain Res. 2013, 256, 205–213. [Google Scholar] [CrossRef]
- Eisenberger, N.I.; Lieberman, M.D.; Williams, K.D. Does Rejection Hurt? An FMRI Study of Social Exclusion. Science 2003, 302, 290–292. [Google Scholar] [CrossRef]
- Rosen, L.H.; Beron, K.J.; Underwood, M.K. Social Victimization Trajectories from Middle Childhood Through Late Adolescence. Soc. Dev. 2017, 26, 227–247. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trevisan, N.; Cattarinussi, G.; Olivo, D.; Di Ciano, A.; Giudetti, L.; Pampallona, A.; Kubera, K.M.; Hirjak, D.; Wolf, R.C.; Sambataro, F. Neural Correlates of Antisocial Behavior: The Victim’s Perspective. Brain Sci. 2023, 13, 474. https://doi.org/10.3390/brainsci13030474
Trevisan N, Cattarinussi G, Olivo D, Di Ciano A, Giudetti L, Pampallona A, Kubera KM, Hirjak D, Wolf RC, Sambataro F. Neural Correlates of Antisocial Behavior: The Victim’s Perspective. Brain Sciences. 2023; 13(3):474. https://doi.org/10.3390/brainsci13030474
Chicago/Turabian StyleTrevisan, Nicolò, Giulia Cattarinussi, Daniele Olivo, Andrea Di Ciano, Lucia Giudetti, Alan Pampallona, Katharina M. Kubera, Dusan Hirjak, Robert Christian Wolf, and Fabio Sambataro. 2023. "Neural Correlates of Antisocial Behavior: The Victim’s Perspective" Brain Sciences 13, no. 3: 474. https://doi.org/10.3390/brainsci13030474
APA StyleTrevisan, N., Cattarinussi, G., Olivo, D., Di Ciano, A., Giudetti, L., Pampallona, A., Kubera, K. M., Hirjak, D., Wolf, R. C., & Sambataro, F. (2023). Neural Correlates of Antisocial Behavior: The Victim’s Perspective. Brain Sciences, 13(3), 474. https://doi.org/10.3390/brainsci13030474