

Dopaminergic System in Promoting Recovery from General Anesthesia

Abstract
:
1. Introduction
2. Dopaminergic System
2.1. Dopaminergic Neurons
2.2. Dopaminergic Neuron Projections
2.3. Dopamine Receptors
3. Dopaminergic System and Arousal
4. Dopaminergic System for General Anesthesia Recovery
4.1. Nervous Nuclei and General Anesthesia
4.1.1. VTA
4.1.2. NAc
4.1.3. vPAG
4.2. Neural Circuit and General Anesthesia
4.3. Dopamine Receptor and General Anesthesia
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brown, E.N.; Purdon, P.L.; Van Dort, C.J. General Anesthesia and Altered States of Arousal: A Systems Neuroscience Analysis. Annu. Rev. Neurosci. 2011, 34, 601–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antkowiak, B. How Do General Anaesthetics Work? Naturwissenschaften 2001, 88, 201–213. [Google Scholar] [CrossRef]
- Franks, N.P. General Anaesthesia: From Molecular Targets to Neuronal Pathways of Sleep and Arousal. Nat. Rev. Neurosci. 2008, 9, 370–386. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Pan, J.; Yu, Y. Regulation of Neural Circuitry under General Anesthesia: New Methods and Findings. Biomolecules 2022, 12, 898. [Google Scholar] [CrossRef] [PubMed]
- Moody, O.A.; Zhang, E.R.; Vincent, K.F.; Kato, R.; Melonakos, E.D.; Nehs, C.J.; Solt, K. The Neural Circuits Underlying General Anesthesia and Sleep. Anesth. Analg. 2021, 132, 1254–1264. [Google Scholar] [CrossRef]
- Pérez-Fernández, J.; Barandela, M.; Jiménez-López, C. The Dopaminergic Control of Movement-Evolutionary Considerations. Int. J. Mol. Sci. 2021, 22, 11284. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Liang, J.; Wang, R.; Yan, T.; Zhou, Y.; Liu, Y.; Feng, Q.; Sun, F.; Li, Y.; Li, A.; et al. The Raphe Dopamine System Controls the Expression of Incentive Memory. Neuron 2020, 106, 498–514.e498. [Google Scholar] [CrossRef]
- Dong, H.; Wang, J.; Yang, Y.F.; Shen, Y.; Qu, W.M.; Huang, Z.L. Dorsal Striatum Dopamine Levels Fluctuate across the Sleep-Wake Cycle and Respond to Salient Stimuli in Mice. Front. Neurosci. 2019, 13, 242. [Google Scholar] [CrossRef] [PubMed]
- Engelhard, B.; Finkelstein, J.; Cox, J.; Fleming, W.; Jang, H.J.; Ornelas, S.; Koay, S.A.; Thiberge, S.Y.; Daw, N.D.; Tank, D.W.; et al. Specialized Coding of Sensory, Motor and Cognitive Variables in Vta Dopamine Neurons. Nature 2019, 570, 509–513. [Google Scholar] [CrossRef]
- Berke, J.D. What Does Dopamine Mean? Nat. Neurosci. 2018, 21, 787–793. [Google Scholar] [CrossRef]
- Monti, J.M.; Monti, D. The Involvement of Dopamine in the Modulation of Sleep and Waking. Sleep Med. Rev. 2007, 11, 113–133. [Google Scholar] [CrossRef] [PubMed]
- Islam, K.U.S.; Meli, N.; Blaess, S. The Development of the Mesoprefrontal Dopaminergic System in Health and Disease. Front. Neural Circuits 2021, 15, 746582. [Google Scholar] [CrossRef]
- Lu, J.; Jhou, T.C.; Saper, C.B. Identification of Wake-Active Dopaminergic Neurons in the Ventral Periaqueductal Gray Matter. J. Neurosci. 2006, 26, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Monti, J.M. The Neurotransmitters of Sleep and Wake, a Physiological Reviews Series. Sleep Med. Rev. 2013, 17, 313–315. [Google Scholar] [CrossRef]
- Leung, L.S.; Luo, T.; Ma, J.; Herrick, I. Brain Areas That Influence General Anesthesia. Prog. Neurobiol. 2014, 122, 24–44. [Google Scholar] [CrossRef]
- Wang, D.; Guo, Q.; Zhou, Y.; Xu, Z.; Hu, S.W.; Kong, X.X.; Yu, Y.M.; Yang, J.X.; Zhang, H.; Ding, H.L.; et al. Gabaergic Neurons in the Dorsal-Intermediate Lateral Septum Regulate Sleep-Wakefulness and Anesthesia in Mice. Anesthesiology 2021, 135, 463–481. [Google Scholar] [CrossRef] [PubMed]
- Daubner, S.C.; Le, T.; Wang, S. Tyrosine Hydroxylase and Regulation of Dopamine Synthesis. Arch. Biochem. Biophys. 2011, 508, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Christenson, J.G.; Dairman, W.; Udenfriend, S. Preparation and Properties of a Homogeneous Aromatic L-Amino Acid Decarboxylase from Hog Kidney. Arch. Biochem. Biophys. 1970, 141, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Garris, P.A.; Ciolkowski, E.L.; Pastore, P.; Wightman, R.M. Efflux of Dopamine from the Synaptic Cleft in the Nucleus Accumbens of the Rat Brain. J. Neurosci. 1994, 14, 6084–6093. [Google Scholar] [CrossRef] [Green Version]
- Eriksen, J.; Jørgensen, T.N.; Gether, U. Regulation of Dopamine Transporter Function by Protein-Protein Interactions: New Discoveries and Methodological Challenges. J. Neurochem. 2010, 113, 27–41. [Google Scholar] [CrossRef]
- Chinta, S.J.; Andersen, J.K. Dopaminergic Neurons. Int. J. Biochem. Cell Biol. 2005, 37, 942–946. [Google Scholar] [CrossRef]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.E.; Lang, A.E. Parkinson Disease. Nat. Rev. Dis. Prim. 2017, 3, 17013. [Google Scholar] [CrossRef] [PubMed]
- Kempadoo, K.A.; Mosharov, E.V.; Choi, S.J.; Sulzer, D.; Kandel, E.R. Dopamine Release from the Locus Coeruleus to the Dorsal Hippocampus Promotes Spatial Learning and Memory. Proc. Natl. Acad. Sci. USA 2016, 113, 14835–14840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Condon, M.D.; Platt, N.J.; Zhang, Y.F.; Roberts, B.M.; Clements, M.A.; Vietti-Michelina, S.; Tseu, M.Y.; Brimblecombe, K.R.; Threlfell, S.; Mann, E.O.; et al. Plasticity in Striatal Dopamine Release Is Governed by Release-Independent Depression and the Dopamine Transporter. Nat. Commun. 2019, 10, 4263. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.R.; Treweek, J.B.; Robinson, J.E.; Xiao, C.; Bremner, L.R.; Greenbaum, A.; Gradinaru, V. Dorsal Raphe Dopamine Neurons Modulate Arousal and Promote Wakefulness by Salient Stimuli. Neuron 2017, 94, 1205–1219.e1208. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wei, J.; Ma, F.; Shan, Q.; Gao, D.; Jin, Y.; Sun, P. Melatonin Modulates Lactation by Regulating Prolactin Secretion Via Tuberoinfundibular Dopaminergic Neurons in the Hypothalamus- Pituitary System. Curr. Protein Pept. Sci. 2020, 21, 744–750. [Google Scholar] [CrossRef] [PubMed]
- Björklund, A.; Dunnett, S.B. Dopamine Neuron Systems in the Brain: An Update. Trends Neurosci. 2007, 30, 194–202. [Google Scholar] [CrossRef]
- Liu, Y.L.; Qin, L.; Wilson, B.; Wu, X.; Qian, L.; Granholm, A.C.; Crews, F.T.; Hong, J.S. Endotoxin Induces a Delayed Loss of Th-Ir Neurons in Substantia Nigra and Motor Behavioral Deficits. Neurotoxicology 2008, 29, 864–870. [Google Scholar] [CrossRef] [Green Version]
- Fields, H.L.; Hjelmstad, G.O.; Margolis, E.B.; Nicola, S.M. Ventral Tegmental Area Neurons in Learned Appetitive Behavior and Positive Reinforcement. Annu. Rev. Neurosci. 2007, 30, 289–316. [Google Scholar] [CrossRef] [Green Version]
- Obeso, J.A.; Marin, C.; Rodriguez-Oroz, C.; Blesa, J.; Benitez-Temiño, B.; Mena-Segovia, J.; Rodríguez, M.; Olanow, C.W. The Basal Ganglia in Parkinson’s Disease: Current Concepts and Unexplained Observations. Ann. Neurol. 2008, 64 (Suppl. S2), S30–S46. [Google Scholar] [CrossRef]
- Wise, R.A. Dopamine, Learning and Motivation. Nat. Rev. Neurosci. 2004, 5, 483–494. [Google Scholar] [CrossRef]
- Smargiassi, A.; Mutti, A. Peripheral Biomarkers and Exposure to Manganese. Neurotoxicology 1999, 20, 401–406. [Google Scholar]
- Klein, M.O.; Battagello, D.S.; Cardoso, A.R.; Hauser, D.N.; Bittencourt, J.C.; Correa, R.G. Dopamine: Functions, Signaling, and Association with Neurological Diseases. Cell Mol. Neurobiol. 2019, 39, 31–59. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Field, G.D. Dopaminergic Modulation of Retinal Processing from Starlight to Sunlight. J. Pharmacol. Sci. 2019, 140, 86–93. [Google Scholar] [CrossRef]
- Indrieri, A.; Pizzarelli, R.; Franco, B.; De Leonibus, E. Dopamine, Alpha-Synuclein, and Mitochondrial Dysfunctions in Parkinsonian Eyes. Front. Neurosci. 2020, 14, 567129. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, J.M.; Espinoza, S.; Gainetdinov, R.R. Dopamine Receptors—Iuphar Review 13. Br. J. Pharmacol. 2015, 172, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, H.; Kato, M.; Takano, H.; Arakawa, R.; Okumura, M.; Otsuka, T.; Kodaka, F.; Hayashi, M.; Okubo, Y.; Ito, H.; et al. Differential Contributions of Prefrontal and Hippocampal Dopamine D(1) and D(2) Receptors in Human Cognitive Functions. J. Neurosci. 2008, 28, 12032–12038. [Google Scholar] [CrossRef] [Green Version]
- Gingrich, J.A.; Caron, M.G. Recent Advances in the Molecular Biology of Dopamine Receptors. Annu. Rev. Neurosci. 1993, 16, 299–321. [Google Scholar] [CrossRef]
- Gurevich, E.V.; Gainetdinov, R.R.; Gurevich, V.V. G Protein-Coupled Receptor Kinases as Regulators of Dopamine Receptor Functions. Pharmacol. Res. 2016, 111, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyson, S.J.; McGonigle, P.; Molinoff, P.B. Quantitative Autoradiographic Localization of the D1 and D2 Subtypes of Dopamine Receptors in Rat Brain. J. Neurosci. 1986, 6, 3177–3188. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, C.; Okamura, H.; Nakajima, T.; Taguchi, J.; Ibata, Y. Autoradiographic Distribution of [3h]Ym-09151-2, a High-Affinity and Selective Antagonist Ligand for the Dopamine D2 Receptor Group, in the Rat Brain and Spinal Cord. J. Comp. Neurol. 1994, 344, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Boyd, K.N.; Mailman, R.B. Dopamine Receptor Signaling and Current and Future Antipsychotic Drugs. Handb. Exp. Pharmacol. 2012, 212, 53–86. [Google Scholar]
- Albert, P.R.; Neve, K.A.; Bunzow, J.R.; Civelli, O. Coupling of a Cloned Rat Dopamine-D2 Receptor to Inhibition of Adenylyl Cyclase and Prolactin Secretion. J. Biol. Chem. 1990, 265, 2098–2104. [Google Scholar] [CrossRef] [PubMed]
- Channer, B.; Matt, S.M.; Nickoloff-Bybel, E.A.; Pappa, V.; Agarwal, Y.; Wickman, J.; Gaskill, P.J. Dopamine, Immunity, and Disease. Pharmacol. Rev. 2023, 75, 62–158. [Google Scholar] [CrossRef] [PubMed]
- Swift, J.L.; Godin, A.G.; Doré, K.; Freland, L.; Bouchard, N.; Nimmo, C.; Sergeev, M.; De Koninck, Y.; Wiseman, P.W.; Beaulieu, J.M. Quantification of Receptor Tyrosine Kinase Transactivation through Direct Dimerization and Surface Density Measurements in Single Cells. Proc. Natl. Acad. Sci. USA 2011, 108, 7016–7021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kisilevsky, A.E.; Zamponi, G.W. D2 Dopamine Receptors Interact Directly with N-Type Calcium Channels and Regulate Channel Surface Expression Levels. Channels 2008, 2, 269–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazelwood, L.A.; Free, R.B.; Cabrera, D.M.; Skinbjerg, M.; Sibley, D.R. Reciprocal Modulation of Function between the D1 and D2 Dopamine Receptors and the Na+,K+-Atpase. J. Biol. Chem. 2008, 283, 36441–36453. [Google Scholar] [CrossRef] [Green Version]
- Noseda, R.; Borsook, D.; Burstein, R. Neuropeptides and Neurotransmitters That Modulate Thalamo-Cortical Pathways Relevant to Migraine Headache. Headache 2017, 57 (Suppl. 2), 97–111. [Google Scholar] [CrossRef] [Green Version]
- Yanagihara, S.; Ikebuchi, M.; Mori, C.; Tachibana, R.O.; Okanoya, K. Arousal State-Dependent Alterations in Neural Activity in the Zebra Finch Vta/Snc. Front. Neurosci. 2020, 14, 897. [Google Scholar] [CrossRef]
- Qiu, M.H.; Zhong, Z.G.; Chen, M.C.; Lu, J. Nigrostriatal and Mesolimbic Control of Sleep-Wake Behavior in Rat. Brain. Struct. Funct. 2019, 224, 2525–2535. [Google Scholar] [CrossRef]
- Wisor, J.P.; Nishino, S.; Sora, I.; Uhl, G.H.; Mignot, E.; Edgar, D.M. Dopaminergic Role in Stimulant-Induced Wakefulness. J. Neurosci. 2001, 21, 1787–1794. [Google Scholar] [CrossRef]
- Rye, D.B.; Jankovic, J. Emerging Views of Dopamine in Modulating Sleep/Wake State from an Unlikely Source: Pd. Neurology 2002, 58, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.E.; Bobillier, P.; Pin, C.; Jouvet, M. The Effect of Lesions of Catecholamine-Containing Neurons Upon Monoamine Content of the Brain and Eeg and Behavioral Waking in the Cat. Brain Res. 1973, 58, 157–177. [Google Scholar] [CrossRef] [PubMed]
- Behbehani, M.M. Functional Characteristics of the Midbrain Periaqueductal Gray. Prog. Neurobiol. 1995, 46, 575–605. [Google Scholar] [CrossRef] [PubMed]
- Porter-Stransky, K.A.; Centanni, S.W.; Karne, S.L.; Odil, L.M.; Fekir, S.; Wong, J.C.; Jerome, C.; Mitchell, H.A.; Escayg, A.; Pedersen, N.P.; et al. Noradrenergic Transmission at Alpha1-Adrenergic Receptors in the Ventral Periaqueductal Gray Modulates Arousal. Biol. Psychiatry 2019, 85, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C.; Bjorkum, A.A.; Gaus, S.E.; Lu, J.; Scammell, T.E.; Saper, C.B. Afferents to the Ventrolateral Preoptic Nucleus. J. Neurosci. 2002, 22, 977–990. [Google Scholar] [CrossRef] [Green Version]
- Verret, L.; Fort, P.; Gervasoni, D.; Léger, L.; Luppi, P.H. Localization of the Neurons Active During Paradoxical (Rem) Sleep and Projecting to the Locus Coeruleus Noradrenergic Neurons in the Rat. J. Comp. Neurol. 2006, 495, 573–586. [Google Scholar] [CrossRef]
- Omelchenko, N.; Sesack, S.R. Periaqueductal Gray Afferents Synapse onto Dopamine and Gaba Neurons in the Rat Ventral Tegmental Area. J. Neurosci. Res. 2010, 88, 981–991. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Chu, R.; Cao, F.; Wang, Y.; Liu, Y.; Cao, J.; Guo, Y.; Mi, W.; Tong, L. Dopaminergic Neurons in the Ventral Tegmental-Prelimbic Pathway Promote the Emergence of Rats from Sevoflurane Anesthesia. Neurosci. Bull. 2022, 38, 417–428. [Google Scholar] [CrossRef]
- Vittoz, N.M.; Berridge, C.W. Hypocretin/Orexin Selectively Increases Dopamine Efflux within the Prefrontal Cortex: Involvement of the Ventral Tegmental Area. Neuropsychopharmacology 2006, 31, 384–395. [Google Scholar] [CrossRef] [Green Version]
- Eban-Rothschild, A.; Rothschild, G.; Giardino, W.J.; Jones, J.R.; de Lecea, L. Vta Dopaminergic Neurons Regulate Ethologically Relevant Sleep-Wake Behaviors. Nat. Neurosci. 2016, 19, 1356–1366. [Google Scholar] [CrossRef] [PubMed]
- Gui, H.; Liu, C.; He, H.; Zhang, J.; Chen, H.; Zhang, Y. Dopaminergic Projections from the Ventral Tegmental Area to the Nucleus Accumbens Modulate Sevoflurane Anesthesia in Mice. Front. Cell Neurosci. 2021, 15, 671473. [Google Scholar] [CrossRef] [PubMed]
- Monti, J.M.; Jantos, H. The Effects of Local Microinjection of Selective Dopamine D1 and D2 Receptor Agonists and Antagonists into the Dorsal Raphe Nucleus on Sleep and Wakefulness in the Rat. Behav. Brain Res. 2018, 339, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gui, H.; Hu, L.; Li, C.; Zhang, J.; Liang, X. Dopamine D1 Receptor in the Nac Shell Is Involved in Delayed Emergence from Isoflurane Anesthesia in Aged Mice. Brain Behav. 2021, 11, e01913. [Google Scholar] [CrossRef]
- Li, Y.D.; Luo, Y.J.; Xu, W.; Ge, J.; Cherasse, Y.; Wang, Y.Q.; Lazarus, M.; Qu, W.M.; Huang, Z.L. Ventral Pallidal Gabaergic Neurons Control Wakefulness Associated with Motivation through the Ventral Tegmental Pathway. Mol. Psychiatry 2021, 26, 2912–2928. [Google Scholar] [CrossRef]
- Clark, M.; Bracci, E. Dichotomous Dopaminergic Control of Ventral Pallidum Neurons. Front. Cell Neurosci. 2018, 12, 260. [Google Scholar] [CrossRef]
- Oishi, Y.; Xu, Q.; Wang, L.; Zhang, B.J.; Takahashi, K.; Takata, Y.; Luo, Y.J.; Cherasse, Y.; Schiffmann, S.N.; de Kerchove, A.; et al. Slow-Wave Sleep Is Controlled by a Subset of Nucleus Accumbens Core Neurons in Mice. Nat. Commun. 2017, 8, 734. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.J.; Li, Y.D.; Wang, L.; Yang, S.R.; Yuan, X.S.; Wang, J.; Cherasse, Y.; Lazarus, M.; Chen, J.F.; Qu, W.M.; et al. Nucleus Accumbens Controls Wakefulness by a Subpopulation of Neurons Expressing Dopamine D(1) Receptors. Nat. Commun. 2018, 9, 1576. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Liu, T.; Dong, Y.; Kondoh, K.; Lu, Z. Trans-Synaptic Neural Circuit-Tracing with Neurotropic Viruses. Neurosci. Bull. 2019, 35, 909–920. [Google Scholar] [CrossRef]
- Li, J.; Liu, Y.; Yuan, L.; Zhang, B.; Bishop, E.S.; Wang, K.; Tang, J.; Zheng, Y.Q.; Xu, W.; Niu, S.; et al. A Tissue-Like Neurotransmitter Sensor for the Brain and Gut. Nature 2022, 606, 94–101. [Google Scholar] [CrossRef]
- Liu, C.; Zhou, X.; Zhu, Q.; Fu, B.; Cao, S.; Zhang, Y.; Zhang, L.; Zhang, Y.; Yu, T. Dopamine Neurons in the Ventral Periaqueductal Gray Modulate Isoflurane Anesthesia in Rats. CNS Neurosci. Ther. 2020, 26, 1121–1133. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gui, H.; Duan, Z.; Yu, T.; Zhang, J.; Liang, X.; Liu, C. Dopamine D1 Receptor in the Nucleus Accumbens Modulates the Emergence from Propofol Anesthesia in Rat. Neurochem. Res. 2021, 46, 1435–1446. [Google Scholar] [CrossRef] [PubMed]
- Swanson, L.W. The Projections of the Ventral Tegmental Area Adjacent Regions: A Combined Fluorescent Retrograde Tracer and Immunofluorescence Study in the Rat. Brain Res. Bull. 1982, 9, 321–353. [Google Scholar] [CrossRef]
- Efimova, E.V.; Gainetdinov, R.R.; Budygin, E.A.; Sotnikova, T.D. Dopamine Transporter Mutant Animals: A Translational Perspective. J. Neurogenet. 2016, 30, 5–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Xu, K.; Yin, J.W.; Zhang, H.; Yin, J.T.; Li, Y. Dopamine Transporter in the Ventral Tegmental Area Modulates Recovery from Propofol Anesthesia in Rats. J. Chem. Neuroanat. 2022, 121, 102083. [Google Scholar] [CrossRef] [PubMed]
- Solt, K.; Van Dort, C.J.; Chemali, J.J.; Taylor, N.E.; Kenny, J.D.; Brown, E.N. Electrical Stimulation of the Ventral Tegmental Area Induces Reanimation from General Anesthesia. Anesthesiology 2014, 121, 311–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, N.E.; Van Dort, C.J.; Kenny, J.D.; Pei, J.; Guidera, J.A.; Vlasov, K.Y.; Lee, J.T.; Boyden, E.S.; Brown, E.N.; Solt, K. Optogenetic Activation of Dopamine Neurons in the Ventral Tegmental Area Induces Reanimation from General Anesthesia. Proc. Natl. Acad. Sci. USA 2016, 113, 12826–12831. [Google Scholar] [CrossRef] [Green Version]
- Soares-Cunha, C.; Coimbra, B.; David-Pereira, A.; Borges, S.; Pinto, L.; Costa, P.; Sousa, N.; Rodrigues, A.J. Activation of D2 Dopamine Receptor-Expressing Neurons in the Nucleus Accumbens Increases Motivation. Nat. Commun. 2016, 7, 11829. [Google Scholar] [CrossRef] [Green Version]
- Kostrzewa, R.M. Neonatal 6-Hydroxydopamine Lesioning of Rats Dopaminergic Neurotoxicity: Proposed Animal Model of Parkinson’s Disease. J. Neural Transm. 2022, 129, 445–461. [Google Scholar] [CrossRef]
- Li, J.; Yu, T.; Shi, F.; Zhang, Y.; Duan, Z.; Fu, B.; Zhang, Y. Involvement of Ventral Periaqueductal Gray Dopaminergic Neurons in Propofol Anesthesia. Neurochem. Res. 2018, 43, 838–847. [Google Scholar] [CrossRef]
- Qiu, G.; Wu, Y.; Yang, Z.; Li, L.; Zhu, X.; Wang, Y.; Sun, W.; Dong, H.; Li, Y.; Hu, J. Dexmedetomidine Activation of Dopamine Neurons in the Ventral Tegmental Area Attenuates the Depth of Sedation in Mice. Anesthesiology 2020, 133, 377–392. [Google Scholar] [CrossRef]
- Solt, K.; Cotten, J.F.; Cimenser, A.; Wong, K.F.; Chemali, J.J.; Brown, E.N. Methylphenidate Actively Induces Emergence from General Anesthesia. Anesthesiology 2011, 115, 791–803. [Google Scholar] [CrossRef] [Green Version]
- Kenny, J.D.; Taylor, N.E.; Brown, E.N.; Solt, K. Dextroamphetamine (but Not Atomoxetine) Induces Reanimation from General Anesthesia: Implications for the Roles of Dopamine and Norepinephrine in Active Emergence. PLoS ONE 2015, 10, e0131914. [Google Scholar] [CrossRef]
- Kato, R.; Zhang, E.R.; Mallari, O.G.; Moody, O.A.; Vincent, K.F.; Melonakos, E.D.; Siegmann, M.J.; Nehs, C.J.; Houle, T.T.; Akeju, O.; et al. D-Amphetamine Rapidly Reverses Dexmedetomidine-Induced Unconsciousness in Rats. Front. Pharmacol. 2021, 12, 668285. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, Y.; Zhang, C.; Wang, M.; Zhang, M.; Yu, L.; Yan, M. The Role of Dopaminergic Vta Neurons in General Anesthesia. PLoS ONE 2015, 10, e0138187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Li, H.; Wang, D.; Guo, Y.; Zhang, X.; Ran, M.; Yang, C.; Yang, Q.; Dong, H. Orexin Activated Emergence from Isoflurane Anaesthesia Involves Excitation of Ventral Tegmental Area Dopaminergic Neurones in Rats. Br. J. Anaesth. 2019, 123, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Gangarossa, G.; Espallergues, J.; de Kerchove, A.; El Mestikawy, S.; Gerfen, C.R.; Hervé, D.; Girault, J.A.; Valjent, E. Distribution and Compartmental Organization of Gabaergic Medium-Sized Spiny Neurons in the Mouse Nucleus Accumbens. Front. Neural Circuits 2013, 7, 22. [Google Scholar] [CrossRef] [Green Version]
- Salgado, S.; Kaplitt, M.G. The Nucleus Accumbens: A Comprehensive Review. Stereotact. Funct. Neurosurg. 2015, 93, 75–93. [Google Scholar] [CrossRef]
- Pain, L.; Gobaille, S.; Schleef, C.; Aunis, D.; Oberling, P. In Vivo Dopamine Measurements in the Nucleus Accumbens after Nonanesthetic and Anesthetic Doses of Propofol in Rats. Anesth. Analg. 2002, 95, 915–919. [Google Scholar] [CrossRef] [Green Version]
- Bao, W.W.; Xu, W.; Pan, G.J.; Wang, T.X.; Han, Y.; Qu, W.M.; Li, W.X.; Huang, Z.L. Nucleus Accumbens Neurons Expressing Dopamine D1 Receptors Modulate States of Consciousness in Sevoflurane Anesthesia. Curr. Biol. 2021, 31, 1893–1902.e1895. [Google Scholar] [CrossRef]
- Zhang, L.N.; Li, Z.J.; Tong, L.; Guo, C.; Niu, J.Y.; Hou, W.G.; Dong, H.L. Orexin-a Facilitates Emergence from Propofol Anesthesia in the Rat. Anesth. Analg. 2012, 115, 789–796. [Google Scholar] [CrossRef]
- Yuan, J.; Luo, Z.; Zhang, Y.; Zhang, Y.; Wang, Y.; Cao, S.; Fu, B.; Yang, H.; Zhang, L.; Zhou, W.; et al. Gabaergic Ventrolateral Pre-Optic Nucleus Neurons Are Involved in the Mediation of the Anesthetic Hypnosis Induced by Propofol. Mol. Med. Rep. 2017, 16, 3179–3186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhang, Y.; Qian, K.; Zhang, L.; Yu, T. Histaminergic H1 and H2 Receptors Mediate the Effects of Propofol on the Noradrenalin-Inhibited Neurons in Rat Ventrolateral Preoptic Nucleus. Neurochem. Res. 2017, 42, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Li, D.D.; Wang, G.Q.; Wu, Q.; Shi, J.S.; Zhang, F. Dendrobium Nobile Lindl Alkaloid Attenuates 6-Ohda-Induced Dopamine Neurotoxicity. Biotechnol. Appl. Biochem. 2021, 68, 1501–1507. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.E.; Chemali, J.J.; Brown, E.N.; Solt, K. Activation of D1 Dopamine Receptors Induces Emergence from Isoflurane General Anesthesia. Anesthesiology 2013, 118, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Ao, Y.; Liu, Y.; Zhang, X.; Li, Y.; Tang, F.; Xu, H. Activation of Dopamine Signals in the Olfactory Tubercle Facilitates Emergence from Isoflurane Anesthesia in Mice. Neurochem. Res. 2021, 46, 1487–1501. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Animal | Nervous Nuclei/ Neural Circuit | General Anesthetics | Intervention /Methods | Findings in Consciousness during General Anesthesia | Reference |
|---|---|---|---|---|---|
| SD rats | VTA-PrL | Sevoflurane | D1R agonist and antagonist were intraperitoneally injected or microinjected into PrL. Chemogenetics and Optogenetics | The dopaminergic pathway of VTA-PrL promotes the emergence of sevoflurane anesthesia. | [58] |
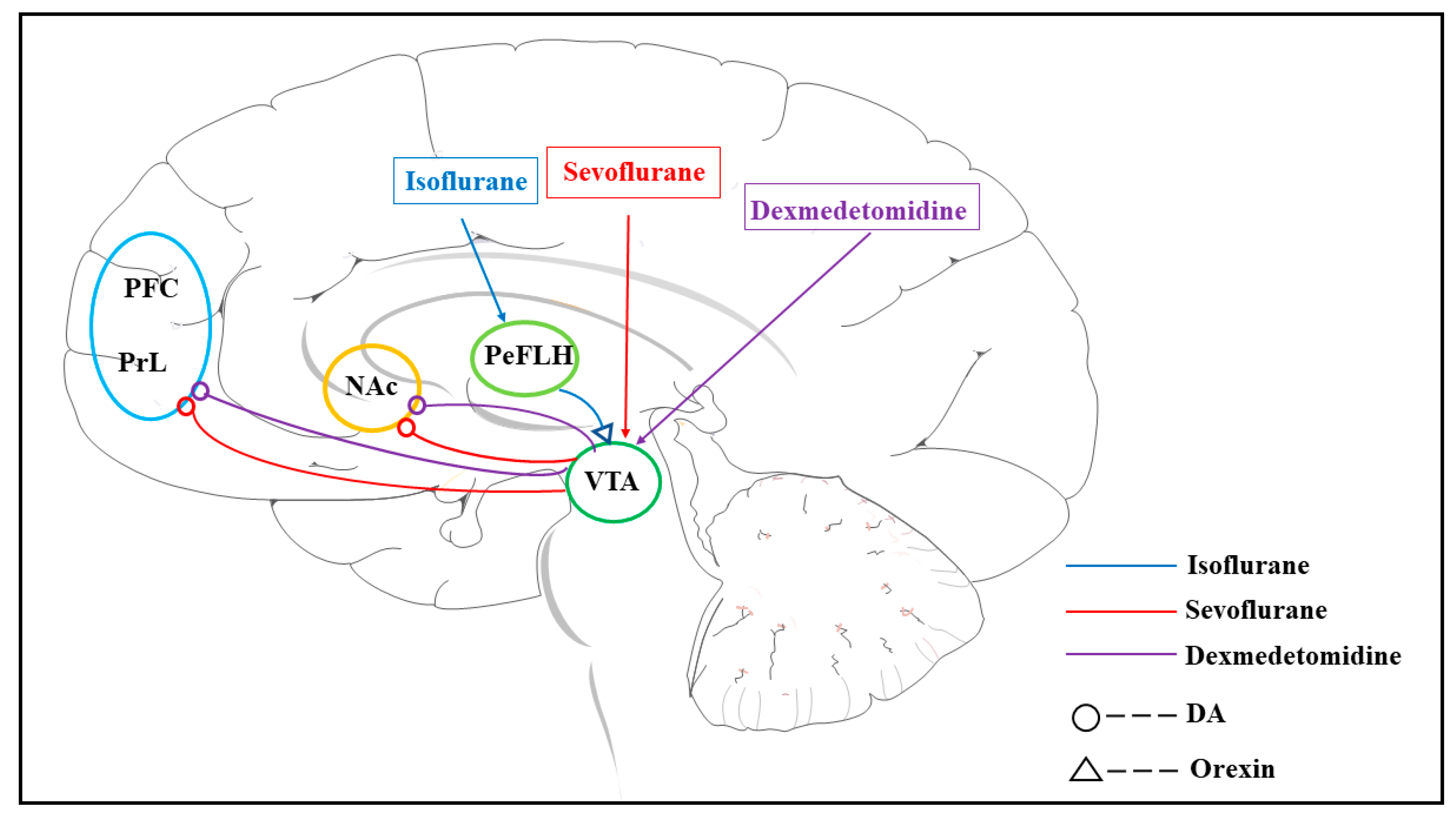
| DAT-cre mice | VTA, VTA-NAc | Sevoflurane, isoflurane | Chemogenetics, Optogenetics, and DA sensor based genetically encoded | Activation of the VTA-NAc dopaminergic pathway delayed the induction and promoted the emergence from general anesthesia | [61] |
| SD rats | vPAG | Isoflurane | Microinjection of 6-OHDA into vPAG. Whole-cell patch clamp recording | Lesion to the vPAG-DA neurons shortens the induction time and prolongs the recovery time in isoflurane anesthesia by activating the GABAA receptor | [70] |
| SD rats | NAc | Propofol | Microinjection of DR agonists and antagonists into NAc. Whole-cell patch clamp recording | The D1R of the NAc MSNs is involved in regulating the emergence of propofol-induced unconsciousness. | [71] |
| SD rats | VTA | Isoflurane, propofol, ketamine | 6-OHDA lesioned Bilateral VTA-DA neurons | VTA-DA neurons may be involved in the emergence of propofol | [73] |
| SD rats | VTA | Propofol | Knockdown DAT | DAT inhibition in VTA enhances PFC neurons activity and promotes recovery after propofol anesthesia. | [74] |
| SD rats | VTA, SN | Isoflurane, propofol | Electrostimulation | Electrical stimulation of VTA, but not SN, induced recovery from anesthesia | [75] |
| DAT-cre mice | VTA | Isoflurane | Optogenetics activation of VTA-DA. DR antagonists were used to intervening | Selective stimulation of VTA-DA neurons induced emergence from general anesthesia | [76] |
| Hcrt-cre rats | VTA PeFLH- VTA | Isoflurane | Microinjection of Orexins into VTA. Identification of Orexin receptors and DA neurons in VTA by immunofluorescence. Optogenetics | Orexin promotes the emergence of isoflurane anesthesia by activating DA neurons in VTA. | [77] |
| D1R-cre mice | NAc | Sevoflurane | Chemogenetics and Optogenetics | Activation of NAc D1R neurons induced cortical activation and behavioral emergence during sevoflurane anesthesia | [78] |
| SD rats | vPAG | Propofol | Microinjection of 6-OHDA into vPAG. Whole-cell patch clamp recording | Lesion to the vPAG-DA neurons shortens the induction time and prolongs the recovery time during propofol anesthesia | [79] |
| DAT-cre mice | VTA-NAc, VTA-mPFC | Dex | DA sensor based genetically encoded. Chemogenomics | Dex activates DA neurons in the VTA and increases DA concentration in the NAc and mPFC. | [80] |
| SD rats | / | Isoflurane | Intraperitoneal injection of DR agonists and antagonists | Activation of D1R mediates emergence from isoflurane | [81] |
| SD rats | / | Sevoflurane, propofol | Dextroamphetamine or atomoxetine is injected intraperitoneally during anesthesia. | Dextroamphetamine induced recovery from general anesthesia, whereas atomoxetine did not | [82] |
| SD rats | / | Dex, ketamine | Dextroamphetamine is administered intravenously during anesthesia | Dextroamphetamine induced recovery after anesthesia with Dex via D1R or D5R, but not with ketamine | [83] |
| TH-IRES-cre mice | OT | Isoflurane | Microinjection of DR agonists and antagonists into OT. Optogenetics | The dopaminergic pathway in OT accelerated emergence from isoflurane anesthesia | [84] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Miao, X.; Sun, Y.; Li, S.; Wu, A.; Wei, C. Dopaminergic System in Promoting Recovery from General Anesthesia. Brain Sci. 2023, 13, 538. https://doi.org/10.3390/brainsci13040538
Wang J, Miao X, Sun Y, Li S, Wu A, Wei C. Dopaminergic System in Promoting Recovery from General Anesthesia. Brain Sciences. 2023; 13(4):538. https://doi.org/10.3390/brainsci13040538
Chicago/Turabian StyleWang, Jinxu, Xiaolei Miao, Yi Sun, Sijie Li, Anshi Wu, and Changwei Wei. 2023. "Dopaminergic System in Promoting Recovery from General Anesthesia" Brain Sciences 13, no. 4: 538. https://doi.org/10.3390/brainsci13040538
APA StyleWang, J., Miao, X., Sun, Y., Li, S., Wu, A., & Wei, C. (2023). Dopaminergic System in Promoting Recovery from General Anesthesia. Brain Sciences, 13(4), 538. https://doi.org/10.3390/brainsci13040538