

Genetic Insights into Obesity and Brain: Combine Mendelian Randomization Study and Gene Expression Analysis
 and
and
Abstract
:1. Introduction
2. Materials and Methods
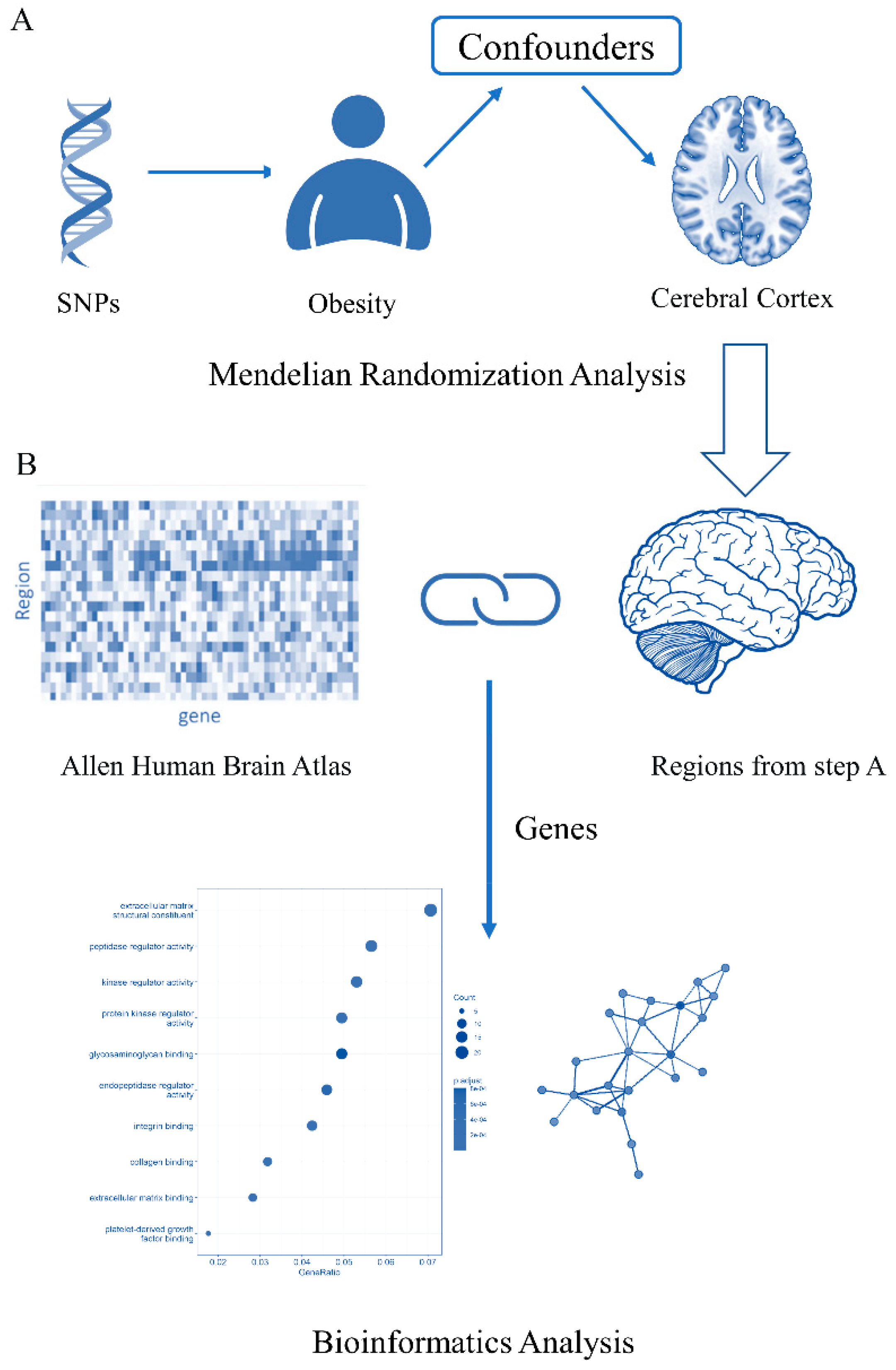
2.1. Study Design
2.2. Data Sources for Obesity and Cortical Structure Phenotype
2.3. Selection of Genetic Variants
2.4. Mendelian Randomization
2.5. Genetic Associations with Brain-Imaging Measurement
3. Results
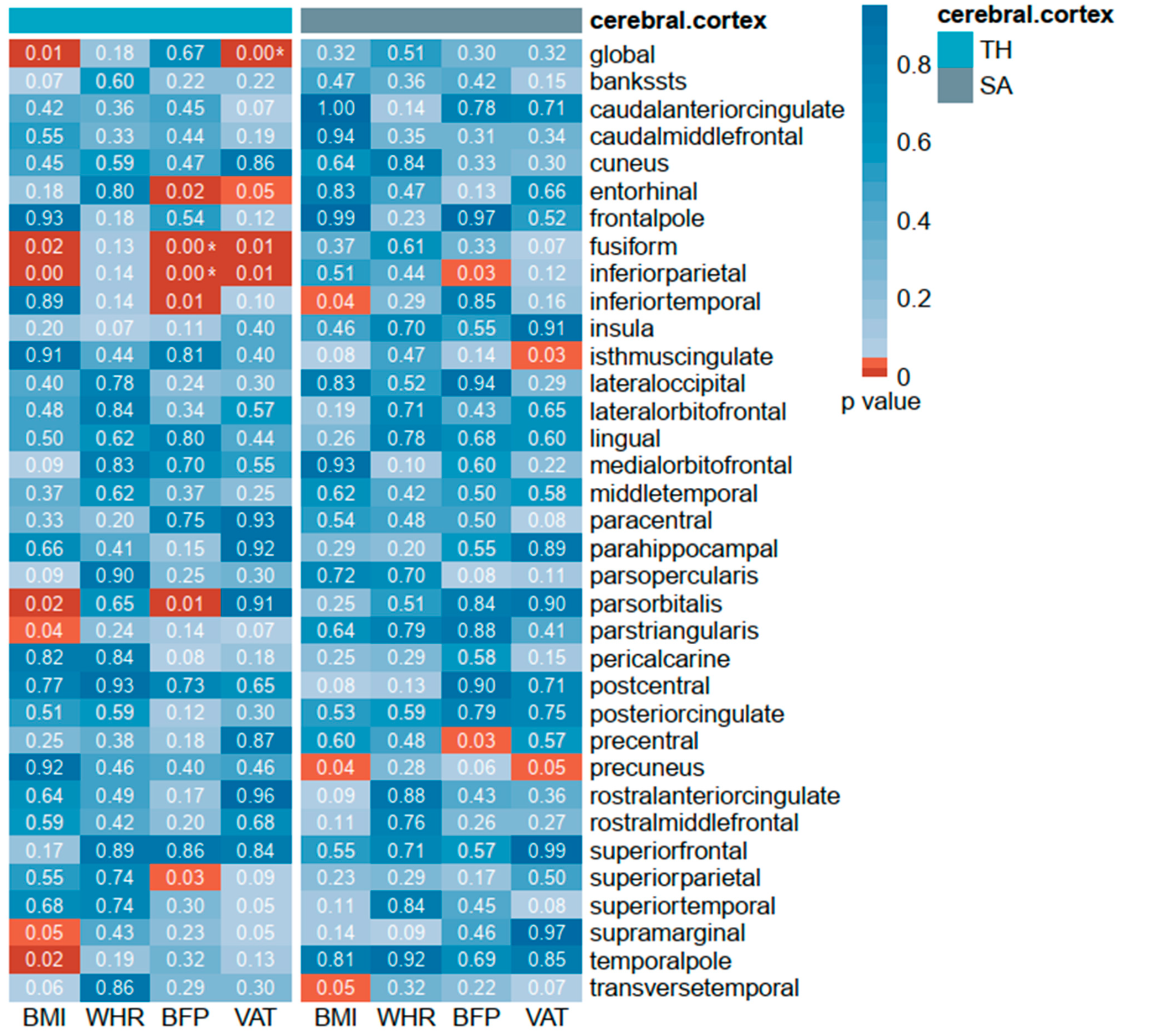
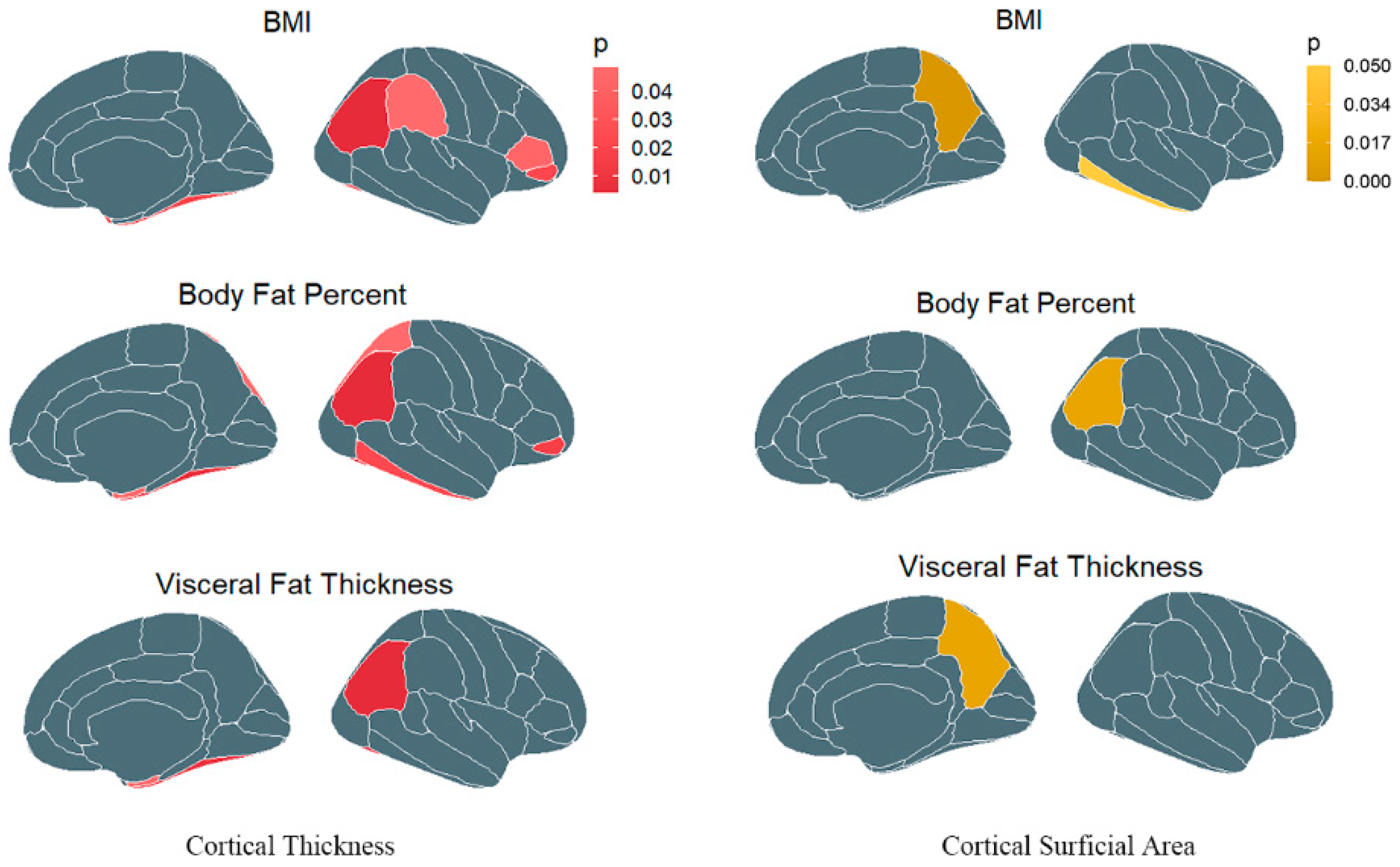
3.1. Causal Association of Obesity with Cerebral Cortex
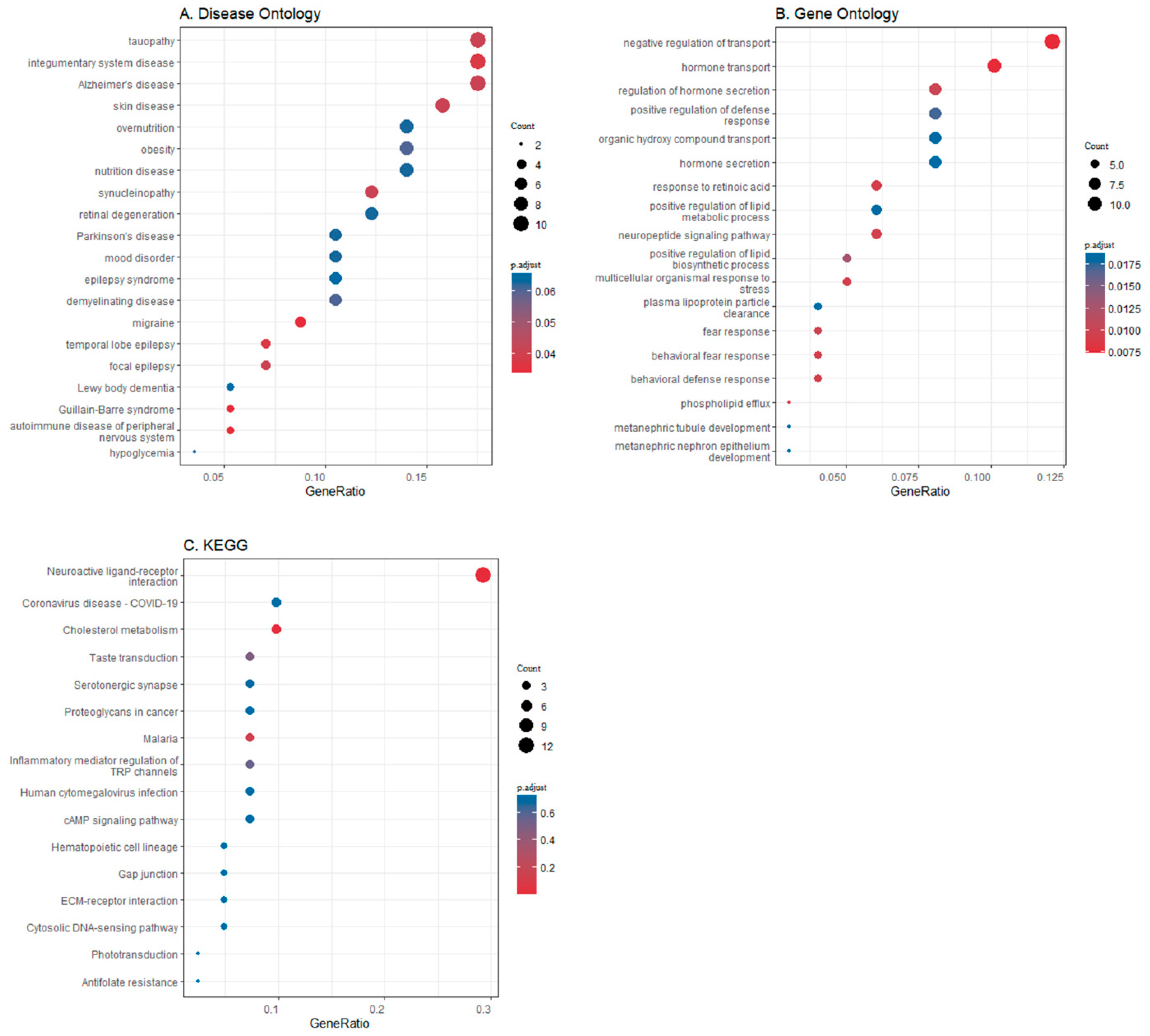
3.2. Differential Gene Expression in Brain Regions
4. Discussion
5. Limitations and Future Research
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AHBA | Allen Human Brain Atlas |
| BFP | body fat percentage |
| BMI | body fat index |
| BP | biological processes |
| CT | cortical thickness |
| DO | Disease Ontology |
| ENIGMA | Enhancing NeuroImaging Genetics through Meta-Analysis |
| GIANT | Genetic Investigation of ANthropometric Traits |
| GO | Gene Ontology |
| GWAS | genome-wide association study |
| IVW | inverse-variance weighting |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| MR | Mendelian randomization |
| MRI | magnetic resonance imaging |
| PPI | protein–protein interaction |
| SA | surface area |
| SNPs | single nucleotide polymorphisms |
| WHR | waist-to-hip ratio |
| VAT | visceral adipose tissue |
References
- James, W.P.T. Obesity: A Global Public Health Challenge. Clin. Chem. 2018, 64, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Yan, A.F.; Shi, Z.; Zhao, B.; Yan, N.; Li, K.; Gao, L.; Xue, H.; Peng, W.; Cheskin, L.J.; et al. Health consequences of obesity and projected future obesity health burden in China. Obesity 2022, 30, 1724–1751. [Google Scholar] [CrossRef] [PubMed]
- Dye, L.; Boyle, N.B.; Champ, C.; Lawton, C. The relationship between obesity and cognitive health and decline. Proc. Nutr. Soc. 2017, 76, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Livingston, G.; Huntley, J.; Sommerlad, A.; Ames, D.; Ballard, C.; Banerjee, S.; Brayne, C.; Burns, A.; Cohen-Mansfield, J.; Cooper, C.; et al. Dementia prevention, intervention, and care: 2020 report of the Lancet Commission. Lancet 2020, 396, 413–446. [Google Scholar] [CrossRef]
- Hamer, M.; Batty, G.D. Association of Body Mass Index and Waist-to-Hip Ratio with Brain Structure: UK Biobank Study. Neurology 2019, 92, e594–e600. [Google Scholar] [CrossRef]
- Raiko, J.R.H.; Tuulari, J.J.; Saari, T.; Parkkola, R.; Savisto, N.; Nuutila, P.; Virtanen, K. Associations between Brain Gray Matter Volumes and Adipose Tissue Metabolism in Healthy Adults. Obesity 2021, 29, 543–549. [Google Scholar] [CrossRef]
- Pflanz, C.P.; Tozer, D.J.; Harshfield, E.L.; Tay, J.; Farooqi, S.; Markus, H.S. Central Obesity Is Selectively Associated with Cerebral Gray Matter Atrophy in 15,634 Subjects in the Uk Biobank. Int. J. Obes. 2022, 46, 1059–1067. [Google Scholar] [CrossRef]
- Medic, N.; Ziauddeen, H.; Ersche, K.D.; Farooqi, I.S.; Bullmore, E.T.; Nathan, P.J.; Ronan, L.; Fletcher, P.C. Increased body mass index is associated with specific regional alterations in brain structure. Int. J. Obes. 2016, 40, 1177–1182. [Google Scholar] [CrossRef]
- Fernández-Andújar, M.; Morales-García, E.; García-Casares, N. Obesity and Gray Matter Volume Assessed by Neu-roimaging: A Systematic Review. Brain Sci. 2021, 11, 999. [Google Scholar] [CrossRef]
- Tanaka, H.; Gourley, D.D.; Dekhtyar, M.; Haley, A.P. Cognition, Brain Structure, and Brain Function in Individuals with Obesity and Related Disorders. Curr. Obes. Rep. 2020, 9, 544–549. [Google Scholar] [CrossRef]
- Saute, R.L.; Soder, R.B.; Filho, J.O.A.; Baldisserotto, M.; Franco, A.R. Increased Brain Cortical Thickness Associated with Visceral Fat in Adoles-cents. Pediatr. Obes. 2018, 13, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Gonzales, M.M.; Strasser, B.; Pasha, E.; McNeely, J.; Tanaka, H.; Haley, A.P. Central Adiposity and Cortical Thickness in Midlife. Psychosom. Med. 2015, 77, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Syan, S.K.; Owens, M.M.; Goodman, B.; Epstein, L.H.; Meyre, D.; Sweet, L.H.; MacKillop, J. Deficits in executive function and suppression of default mode network in obesity. NeuroImage Clin. 2019, 24, 102015. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Ouyang, M.; Wang, J.; Xie, M.; Huang, Y.; Yuan, F.; Jia, Y.; Zhang, X.; Liu, N.; Zhang, N. Shared genetics between classes of obesity and psychiatric disorders: A large-scale genome-wide cross-trait analysis. J. Psychosom. Res. 2022, 162, 111032. [Google Scholar] [CrossRef]
- Yao, S.; Wu, H.; Ding, J.-M.; Wang, Z.-X.; Ullah, T.; Dong, S.-S.; Chen, H.; Guo, Y. Transcriptome-wide association study identifies multiple genes associated with childhood body mass index. Int. J. Obes. 2021, 45, 1105–1113. [Google Scholar] [CrossRef]
- Rossi, M.A.; Basiri, M.L.; McHenry, J.A.; Kosyk, O.; Otis, J.M.; Munkhof, H.E.V.D.; Bryois, J.; Hübel, C.; Breen, G.; Guo, W.; et al. Obesity remodels activity and transcriptional state of a lateral hypothalamic brake on feeding. Science 2019, 364, 1271–1274. [Google Scholar] [CrossRef]
- Lyu, P.; Huang, Z.; Feng, Q.; Su, Y.; Zheng, M.; Hong, Y.; Cai, X.; Lu, Z. Unveiling the transcriptome alteration of POMC neuron in diet-induced obesity. Exp. Cell Res. 2020, 389, 111848. [Google Scholar] [CrossRef]
- Fornito, A.; Arnatkevičiūtė, A.; Fulcher, B.D. Bridging the Gap between Connectome and Transcriptome. Trends Cogn. Sci. 2019, 23, 34–50. [Google Scholar] [CrossRef]
- Lawlor, D.A.; Harbord, R.M.; Sterne, J.A.C.; Timpson, N.; Smith, G.D. Mendelian randomization: Using genes as instruments for making causal inferences in epidemiology. Stat. Med. 2008, 27, 1133–1163. [Google Scholar] [CrossRef]
- Sekula, P.; Del Greco, M.F.; Pattaro, C.; Köttgen, A. Mendelian Randomization as an Approach to Assess Causality Using Observa-tional Data. J. Am. Soc. Nephrol. 2016, 27, 3253–3265. [Google Scholar] [CrossRef]
- Skrivankova, V.W.; Richmond, R.C.; Woolf, B.A.R.; Yarmolinsky, J.; Davies, N.M.; Swanson, S.A.; VanderWeele, T.J.; Higgins, J.P.T.; Timpson, N.J.; Dimou, N.; et al. Strengthening the Reporting of Observational Studies in Epidemi-ology Using Mendelian Randomization: The Strobe-Mr Statement. JAMA 2021, 326, 1614–1621. [Google Scholar] [CrossRef] [PubMed]
- Yengo, L.; Sidorenko, J.; Kemper, K.E.; Zheng, Z.; Wood, A.R.; Weedon, M.N.; Frayling, T.M.; Hirschhorn, J.; Yang, J.; Visscher, P.M.; et al. Meta-analysis of genome-wide association studies for height and body mass index in ∼700000 individuals of European ancestry. Hum. Mol. Genet. 2018, 27, 3641–3649. [Google Scholar] [CrossRef]
- Pulit, S.L.; Stoneman, C.; Morris, A.P.; Wood, A.R.; Glastonbury, C.A.; Tyrrell, J.; Yengo, L.; Ferreira, T.; Marouli, E.; Ji, Y.; et al. Meta-analysis of genome-wide association studies for body fat distribution in 694 649 individuals of European ancestry. Hum. Mol. Genet. 2019, 28, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, T.; Rask-Andersen, M.; Pan, G.; Höglund, J.; Wadelius, C.; Ek, W.E.; Johansson, Å. Contribution of genetics to visceral adiposity and its relation to cardiovascular and metabolic disease. Nat. Med. 2019, 25, 1390–1395. [Google Scholar] [CrossRef] [PubMed]
- Grasby, K.L.; Jahanshad, N.; Painter, J.N.; Colodro-Conde, L.; Bralten, J.; Hibar, D.P.; Lind, P.A.; Pizzagalli, F.; Ching, C.R.K.; McMahon, M.A.B.; et al. The genetic architecture of the human cerebral cortex. Science 2020, 367, eaay6690. [Google Scholar] [CrossRef] [PubMed]
- Desikan, R.S.; Ségonne, F.; Fischl, B.; Quinn, B.T.; Dickerson, B.C.; Blacker, D.; Buckner, R.L.; Dale, A.M.; Maguire, R.P.; Hyman, B.T.; et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. NeuroImage 2006, 31, 968–980. [Google Scholar] [CrossRef]
- Burgess, S.; Thompson, S.G. Avoiding Bias from Weak Instruments in Mendelian Randomization Studies. Int. J. Epidemiol. 2011, 40, 755–764. [Google Scholar] [CrossRef]
- Pierce, B.L.; Ahsan, H.; Vanderweele, T.J. Power and Instrument Strength Requirements for Mendelian Randomization Studies Using Multiple Genetic Variants. Int. J. Epidemiol. 2011, 40, 740–752. [Google Scholar] [CrossRef]
- Burgess, S.; Davies, N.M.; Thompson, S.G. Bias Due to Participant Overlap in Two-Sample Mendelian Randomization. Genet. Epidemiol. 2016, 40, 597–608. [Google Scholar] [CrossRef]
- Verbanck, M.; Chen, C.Y.; Neale, B.; Do, R. Detection of Widespread Horizontal Pleiotropy in Causal Relationships Inferred from Mendelian Randomization between Complex Traits and Diseases. Nat. Genet. 2018, 50, 693–698. [Google Scholar] [CrossRef]
- Burgess, S.; Butterworth, A.; Thompson, S.G. Mendelian Randomization Analysis with Multiple Genetic Variants Using Summarized Data. Genet Epidemiol. 2013, 37, 658–665. [Google Scholar] [CrossRef]
- Del Greco, M.F.; Minelli, C.; Sheehan, N.A.; Thompson, J.R. Detecting pleiotropy in Mendelian randomisation studies with summary data and a continuous outcome. Stat. Med. 2015, 34, 2926–2940. [Google Scholar] [CrossRef]
- Bowden, J.; Davey Smith, G.; Burgess, S. Mendelian randomization with invalid instruments: Effect estimation and bias detection through Egger regression. Int. J. Epidemiol. 2015, 44, 512–525. [Google Scholar] [CrossRef] [PubMed]
- Hemani, G.; Tilling, K.; Smith, G.D. Orienting the Causal Relationship between Imprecisely Measured Traits Using Gwas Summary Data. PLoS Genet. 2017, 13, e1007081. [Google Scholar]
- Brion, M.-J.A.; Shakhbazov, K.; Visscher, P. Calculating statistical power in Mendelian randomization studies. Int. J. Epidemiol. 2013, 42, 1497–1501. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, E. Multivariable Mendelian Randomization and Mediation. Cold Spring Harb Perspect Med. 2021, 11, a038984. [Google Scholar] [CrossRef] [PubMed]
- Shen, E.H.; Overly, C.C.; Jones, A.R. The Allen Human Brain Atlas: Comprehensive Gene Expression Mapping of the Human Brain. Trends Neurosci. 2012, 35, 711–714. [Google Scholar] [CrossRef] [PubMed]
- Hawrylycz, M.J.; Lein, E.S.; Guillozet-Bongaarts, A.L.; Shen, E.H.; Ng, L.; Miller, J.A.; Van De Lagemaat, L.N.; Smith, K.A.; Ebbert, A.; Riley, Z.L.; et al. An Anatomically Comprehensive Atlas of the Adult Human Brain Transcriptome. Nature 2012, 489, 391–399. [Google Scholar] [CrossRef]
- Tomkins, J.E.; Manzoni, C. Advances in protein-protein interaction network analysis for Parkinson’s disease. Neurobiol. Dis. 2021, 155, 105395. [Google Scholar] [CrossRef]
- Gardener, H.; Caunca, M.; Dong, C.; Cheung, Y.K.; Rundek, T.; Elkind, M.S.V.; Wright, C.B.; Sacco, R.L. Obesity Measures in Relation to Cognition in the Northern Manhattan Study. J. Alzheimers Dis. 2020, 78, 1653–1660. [Google Scholar] [CrossRef]
- Frank, A.P.; Santos, R.d.S.; Palmer, B.F.; Clegg, D.J. Determinants of body fat distribution in humans may provide insight about obesity-related health risks. J. Lipid Res. 2019, 60, 1710–1719. [Google Scholar] [CrossRef] [PubMed]
- Chooi, Y.C.; Ding, C.; Magkos, F. The Epidemiology of Obesity. Metabolism 2019, 92, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Sangachin, M.G.; Cavuoto, L.A.; Wang, Y. Use of Various Obesity Measurement and Classification Methods in Occupational Safety and Health Research: A Systematic Review of the Literature. BMC Obes. 2018, 5, 28. [Google Scholar]
- Farruggia, M.C.; Small, D.M. Effects of Adiposity and Metabolic Dysfunction on Cognition: A Review. Physiol. Behav. 2019, 208, 112578. [Google Scholar] [CrossRef]
- Petry, N.M.; Barry, D.; Pietrzak, R.H.; Wagner, J.L. Overweight and Obesity Are Associated with Psychiatric Disorders: Results from the National Epidemiologic Survey on Alcohol and Related Conditions. Psychosom. Me. 2008, 70, 288–297. [Google Scholar] [CrossRef]
- Slomski, A. Obesity Is Now the Top Modifiable Dementia Risk Factor in the Us. JAMA 2022, 328, 10. [Google Scholar] [CrossRef] [PubMed]
- Weiner, M.W.; Veitch, D.P.; Aisen, P.S.; Beckett, A.L.; Cairns, J.N.; Green, R.C.; Harvey, D.; Jack, C.R.; Jagust, W.; Liu, E.; et al. The Alzheimer’s Disease Neuroimaging Initiative: A Review of Papers Published since Its Inception. Alzheimers Dement. 2013, 8, S1–S68. [Google Scholar] [CrossRef]
- Opel, N.; Thalamuthu, A.; Milaneschi, Y.; Grotegerd, D.; Flint, C.; Leenings, R.; Goltermann, J.; Richter, M.; Hahn, T.; Woditsch, G.; et al. Brain Structural Abnormalities in Obesity: Relation to Age, Genetic Risk, and Common Psychiatric Disorders: Evidence through Univariate and Multivariate Mega-Analysis Including 6420 Participants from the Enigma Mdd Working Group. Mol. Psychiatry 2021, 26, 4839–4852. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.; Cho, H.; Kim, J.; Lee, D.-W.; Kim, G.H.; Hong, Y.S.; Moon, S.; Park, S.; Lee, S.; Lee, S.; et al. Brain changes in overweight/obese and normal-weight adults with type 2 diabetes mellitus. Diabetologia 2017, 60, 1207–1217. [Google Scholar] [CrossRef]
- Ottino-González, J.; Jurado, M.A.; García-García, I.; Segura, B.; Marqués-Iturria, I.; Sender-Palacios, M.J.; Tor, E.; Prats-Soteras, X.; Caldú, X.; Junqué, C.; et al. Allostatic Load Is Linked to Cortical Thickness Changes Depending on Body-Weight Status. Front. Hum. Neurosci. 2017, 11, 639. [Google Scholar] [CrossRef]
- O’Brien, P.D.; Hinder, L.M.; Callaghan, B.C.; Feldman, E.L. Neurological consequences of obesity. Lancet Neurol. 2017, 16, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Apo, E.; García-Sierra, A.; Silva-Pereyra, J.; Soto-Abraham, V.; Mondragón-Maya, A.; Velasco-Vales, V.; Pescatello, L.S. A Postmortem Study of Frontal and Temporal Gyri Thickness and Cell Number in Human Obesity. Obesity 2018, 26, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Apo, E.; Mondragón-Maya, A.; Ferrari-Díaz, M.; Silva-Pereyra, J. Structural Brain Changes Associated with Overweight and Obesity. J. Obes. 2021, 2021, 6613385. [Google Scholar] [CrossRef]
- Igelström, K.M.; Graziano, M.S. The inferior parietal lobule and temporoparietal junction: A network perspective. Neuropsychologia 2017, 105, 70–83. [Google Scholar] [CrossRef] [PubMed]
- Thomas, K.R.; Osuna, J.R.; Weigand, A.J.; Edmonds, E.C.; Clark, A.L.; Holmqvist, S.; Cota, I.H.; Wierenga, C.E.; Bondi, M.W.; Bangen, K.J.; et al. Regional hyperperfusion in older adults with objectively-defined subtle cognitive decline. J. Cereb. Blood Flow. Metab. 2021, 41, 1001–1012. [Google Scholar] [CrossRef]
- Bokde, A.L.W.; Lopez-Bayo, P.; Meindl, T.; Pechler, S.; Born, C.; Faltraco, F.; Teipel, S.J.; Möller, H.-J.; Hampel, H. Functional connectivity of the fusiform gyrus during a face-matching task in subjects with mild cognitive impairment. Brain 2006, 129, 1113–1124. [Google Scholar] [CrossRef]
- Karagiannides, I.; Abdou, R.; Tzortzopoulou, A.; Voshol, P.J.; Kypreos, K.E. Apolipoprotein E Predisposes to Obesity and Related Metabolic Dys-functions in Mice. FEBS J. 2008, 275, 4796–4809. [Google Scholar] [CrossRef]
- Martínez-Martínez, A.B.; Torres-Perez, E.; Devanney, N.; Del Moral, R.; Johnson, L.A.; Arbones-Mainar, J.M. Beyond the CNS: The many peripheral roles of APOE. Neurobiol. Dis. 2020, 138, 104809. [Google Scholar] [CrossRef]
- Serrano-Pozo, A.; Das, S.; Hyman, B.T. Apoe and Alzheimer’s Disease: Advances in Genetics, Pathophysiology, and Therapeutic Approaches. Lancet Neurol. 2021, 20, 68–80. [Google Scholar] [CrossRef]
- Liu, L.; Chan, C. The role of inflammasome in Alzheimer’s disease. Ageing Res. Rev. 2014, 15, 6–15. [Google Scholar] [CrossRef]
- Ghanbari, M.; Momen Maragheh, S.; Aghazadeh, A.; Mehrjuyan, S.R.; Hussen, B.M.; Abdoli Shadbad, M.; Dastmalchi, N.; Safaralizadeh, R. Interleukin-1 in Obesity-Related Low-Grade Inflammation: From Molecular Mechanisms to Therapeutic Strategies. Int. Immunopharmacol. 2021, 96, 107765. [Google Scholar] [CrossRef] [PubMed]
- Magliozzi, R.; Pitteri, M.; Ziccardi, S.; Pisani, A.I.; Montibeller, L.; Marastoni, D.; Rossi, S.; Mazziotti, V.; Guandalini, M.; Dapor, C.; et al. CSF parvalbumin levels reflect interneuron loss linked with cortical pathology in multiple sclerosis. Ann. Clin. Transl. Neurol. 2021, 8, 534–547. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Gan, J.; Jonas, P. Interneurons. Fast-Spiking, Parvalbumin⁺ Gabaergic Interneurons: From Cellular Design to Microcircuit Function. Science 2014, 345, 1255263. [Google Scholar] [CrossRef]
- Lin, S.; Zhang, A.; Yuan, L.; Wang, Y.; Zhang, C.; Jiang, J.; Xu, H.; Yuan, H.; Yao, H.; Zhang, Q.; et al. Targeting parvalbumin promotes M2 macrophage polarization and energy expenditure in mice. Nat. Commun. 2022, 13, 3301. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Brouwers, B.; Liu, H.; Liu, H.; Lawler, K.; de Oliveira, E.M.; Lee, D.-K.; Yang, Y.; Cox, A.R.; Keogh, J.M.; et al. Human loss-of-function variants in the serotonin 2C receptor associated with obesity and maladaptive behavior. Nat. Med. 2022, 28, 2537–2546. [Google Scholar] [CrossRef]
- Li, X.; Chen, W.; Pan, K.; Li, H.; Pang, P.; Guo, Y.; Shu, S.; Cai, Y.; Pei, L.; Liu, D.; et al. Serotonin receptor 2c-expressing cells in the ventral CA1 control attention via innervation of the Edinger–Westphal nucleus. Nat. Neurosci. 2018, 21, 1239–1250. [Google Scholar] [CrossRef]
- Liu, S.; Seidlitz, J.; Blumenthal, J.D.; Clasen, L.S.; Raznahan, A. Integrative structural, functional, and transcriptomic analyses of sex-biased brain organization in humans. Proc. Natl. Acad. Sci. USA 2020, 117, 18788–18798. [Google Scholar] [CrossRef]
- Mayoral, L.P.C.; Andrade, G.M.; Mayoral, E.P.; Huerta, T.H.; Canseco, S.P.; Canales, F.J.R.; Cabrera-Fuentes, H.A.; Cruz, M.M.; Santiago, A.D.P.; Alpuche, J.J.; et al. Obesity Subtypes, Related Biomarkers & Heterogeneity. Indian J. Med. Res. 2020, 151, 11–21. [Google Scholar]
- Deng, Y.-T.; Li, Y.-Z.; Huang, S.-Y.; Ou, Y.-N.; Zhang, W.; Chen, S.-D.; Zhang, Y.-R.; Yang, L.; Dong, Q.; Feng, J.-F.; et al. Association of life course adiposity with risk of incident dementia: A prospective cohort study of 322,336 participants. Mol. Psychiatry 2022, 27, 3385–3395. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
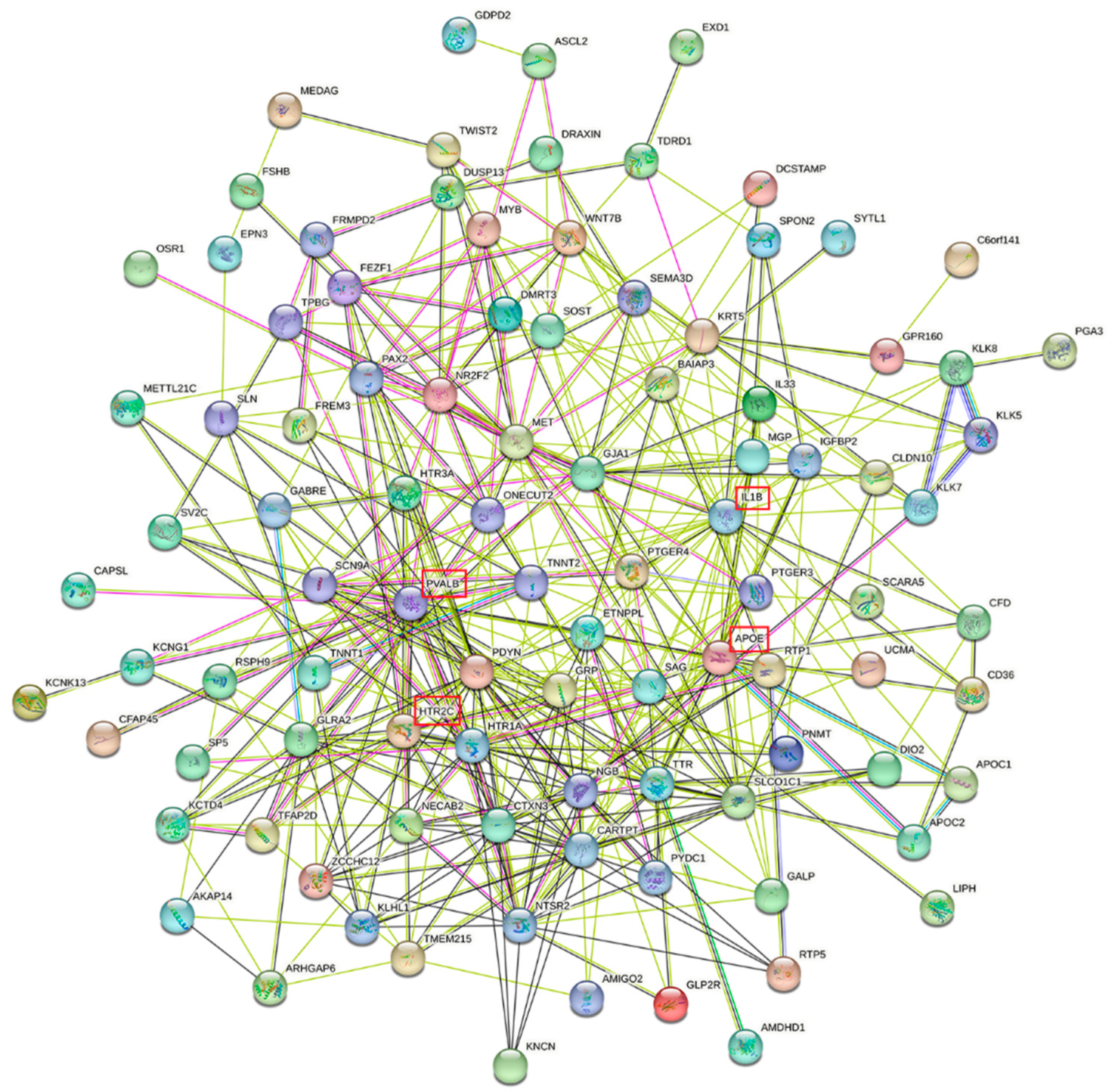{kind=link}
| Exposure | Unit | Consortium or Study | Sex | Sample Size | Population | SNP Number | F-Value |
|---|---|---|---|---|---|---|---|
| BMI | SD (kg/m2) | GIANT+UKB | Males and females | 681,275 | European | 513 | 74.036 |
| WHR | SD | GIANT+UKB | Males and females | 697,734 | European | 72 | 71.708 |
| BFP | SD (%) | UKB | Males and females | 492,787 | European | 300 | 42.851 |
| VAT | SD (kg) | UKB | Males and females | 325,153 | European | 220 | 57.661 |
| Exposure | NSNP | Beta | SE | p | Exposure | NSNP | Beta | SE | p |
|---|---|---|---|---|---|---|---|---|---|
| CT−global | SA_inferior parietal | ||||||||
| BMI | 20 | −0.026 | 0.052 | 0.623 | BMI | 20 | 268.238 | 241.654 | 0.267 |
| BFP | 20 | 0.006 | 0.033 | 0.860 | BFP | 20 | 13.166 | 156.641 | 0.933 |
| VAT | 21 | 0.015 | 0.055 | 0.787 | VAT | 21 | −315.274 | 255.332 | 0.217 |
| CT_entorhinal | SA_inferior temporal | ||||||||
| BMI | 20 | 0.084 | 0.163 | 0.607 | BMI | 20 | −167.396 | 139.979 | 0.232 |
| BFP | 20 | −0.032 | 0.106 | 0.762 | BFP | 20 | −148.798 | 90.549 | 0.100 |
| VAT | 21 | −0.023 | 0.173 | 0.894 | VAT | 21 | 214.826 | 147.914 | 0.146 |
| CT_fusiform | SA_isthmus cingulate | ||||||||
| BMI | 20 | −0.007 | 0.046 | 0.875 | BMI | 20 | −25.197 | 62.640 | 0.688 |
| BFP | 20 | −0.066 | 0.030 | 0.029 * | BFP | 20 | −43.801 | 40.681 | 0.282 |
| VAT | 21 | 0.031 | 0.048 | 0.519 | VAT | 21 | 73.874 | 66.186 | 0.264 |
| CT_inferior parietal | SA_precentral | ||||||||
| BMI | 20 | −0.016 | 0.038 | 0.676 | BMI | 20 | 23.006 | 152.876 | 0.880 |
| BFP | 20 | 0.020 | 0.024 | 0.399 | BFP | 20 | −71.641 | 98.818 | 0.468 |
| VAT | 21 | 0.007 | 0.040 | 0.852 | VAT | 21 | −10.743 | 161.507 | 0.947 |
| CT_inferior temporal | SA_precuneus | ||||||||
| BMI | 20 | 0.058 | 0.047 | 0.215 | BMI | 20 | −92.171 | 166.077 | 0.579 |
| BFP | 20 | 0.036 | 0.030 | 0.238 | BFP | 20 | −226.395 | 107.496 | 0.035* |
| VAT | 21 | −0.083 | 0.049 | 0.091 | VAT | 21 | 240.129 | 175.493 | 0.171 |
| CT_pars orbitalis | SA_transverse temporal | ||||||||
| BMI | 20 | −0.118 | 0.067 | 0.079 | BMI | 20 | 18.932 | 27.781 | 0.496 |
| BFP | 20 | 0.005 | 0.044 | 0.913 | BFP | 20 | 14.190 | 18.018 | 0.431 |
| VAT | 21 | 0.105 | 0.071 | 0.141 | VAT | 21 | −11.132 | 29.354 | 0.705 |
| CT_pars triangularis | |||||||||
| BMI | 20 | −0.059 | 0.068 | 0.383 | |||||
| BFP | 20 | −0.060 | 0.043 | 0.165 | |||||
| VAT | 21 | 0.079 | 0.071 | 0.268 | |||||
| CT_superiorparietal | |||||||||
| BMI | 20 | −0.090 | 0.044 | 0.038 * | |||||
| BFP | 20 | 0.026 | 0.028 | 0.351 | |||||
| VAT | 21 | 0.077 | 0.046 | 0.095 | |||||
| CT_supramarginal | |||||||||
| BMI | 20 | −0.014 | 0.046 | 0.764 | |||||
| BFP | 20 | 0.016 | 0.030 | 0.590 | |||||
| VAT | 21 | −0.003 | 0.048 | 0.953 | |||||
| CT_temporalpole | |||||||||
| BMI | 20 | 0.026 | 0.147 | 0.860 | |||||
| BFP | 20 | −0.063 | 0.095 | 0.509 | |||||
| VAT | 21 | −0.003 | 0.155 | 0.983 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Zhao, S.; Wang, Y.; Niu, X.; Zhang, B.; Li, X.; Peng, D. Genetic Insights into Obesity and Brain: Combine Mendelian Randomization Study and Gene Expression Analysis. Brain Sci. 2023, 13, 892. https://doi.org/10.3390/brainsci13060892
Chen L, Zhao S, Wang Y, Niu X, Zhang B, Li X, Peng D. Genetic Insights into Obesity and Brain: Combine Mendelian Randomization Study and Gene Expression Analysis. Brain Sciences. 2023; 13(6):892. https://doi.org/10.3390/brainsci13060892
Chicago/Turabian StyleChen, Leian, Shaokun Zhao, Yuye Wang, Xiaoqian Niu, Bin Zhang, Xin Li, and Dantao Peng. 2023. "Genetic Insights into Obesity and Brain: Combine Mendelian Randomization Study and Gene Expression Analysis" Brain Sciences 13, no. 6: 892. https://doi.org/10.3390/brainsci13060892
APA StyleChen, L., Zhao, S., Wang, Y., Niu, X., Zhang, B., Li, X., & Peng, D. (2023). Genetic Insights into Obesity and Brain: Combine Mendelian Randomization Study and Gene Expression Analysis. Brain Sciences, 13(6), 892. https://doi.org/10.3390/brainsci13060892