
Audiovisual n-Back Training Alters the Neural Processes of Working Memory and Audiovisual Integration: Evidence of Changes in ERPs

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Participants
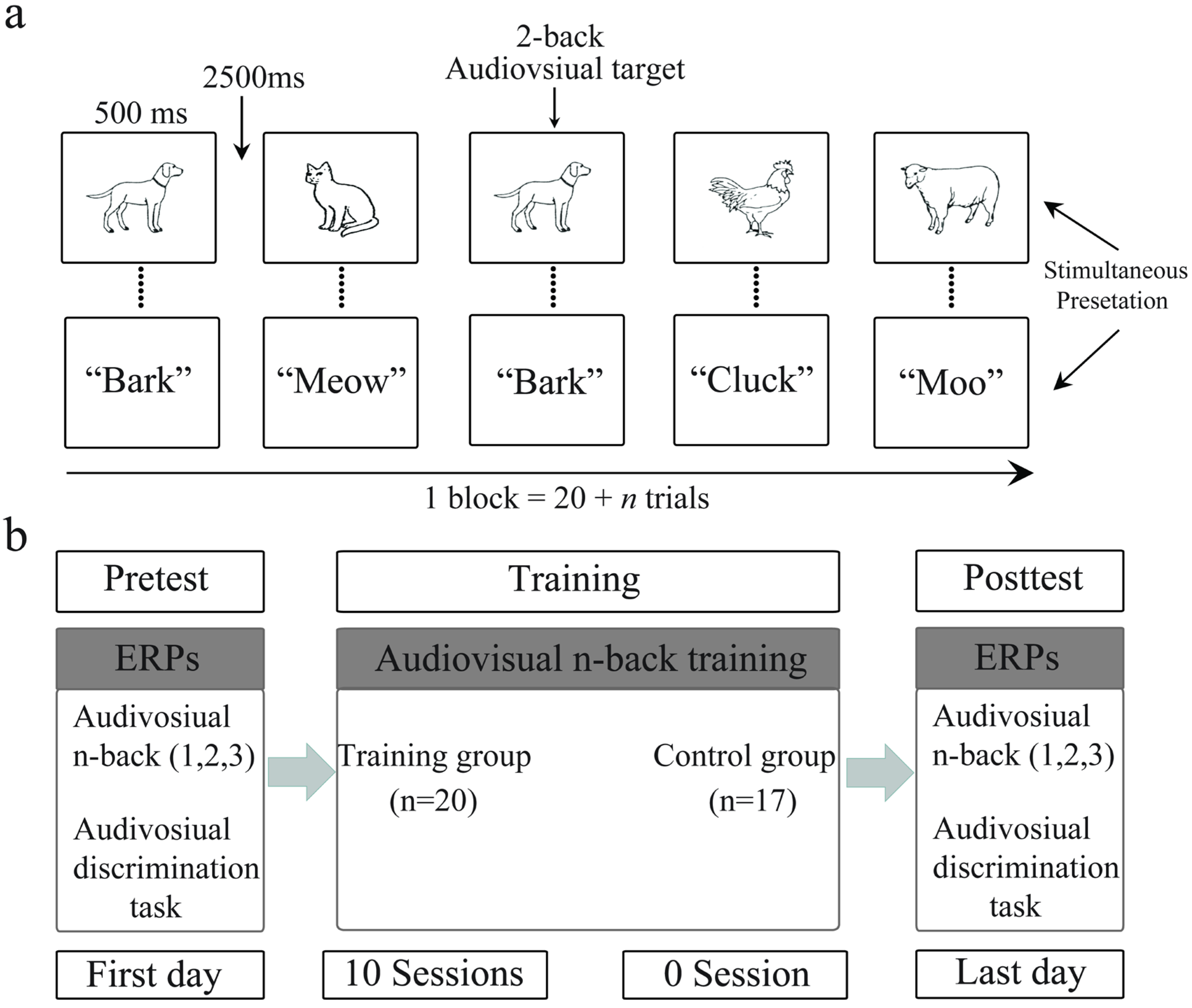
2.2. General Procedure
2.2.1. Training Task
2.2.2. Training and Transfer Outcomes
2.3. EEG Data Recording and Preprocessing
2.4. Data Analysis
3. Results
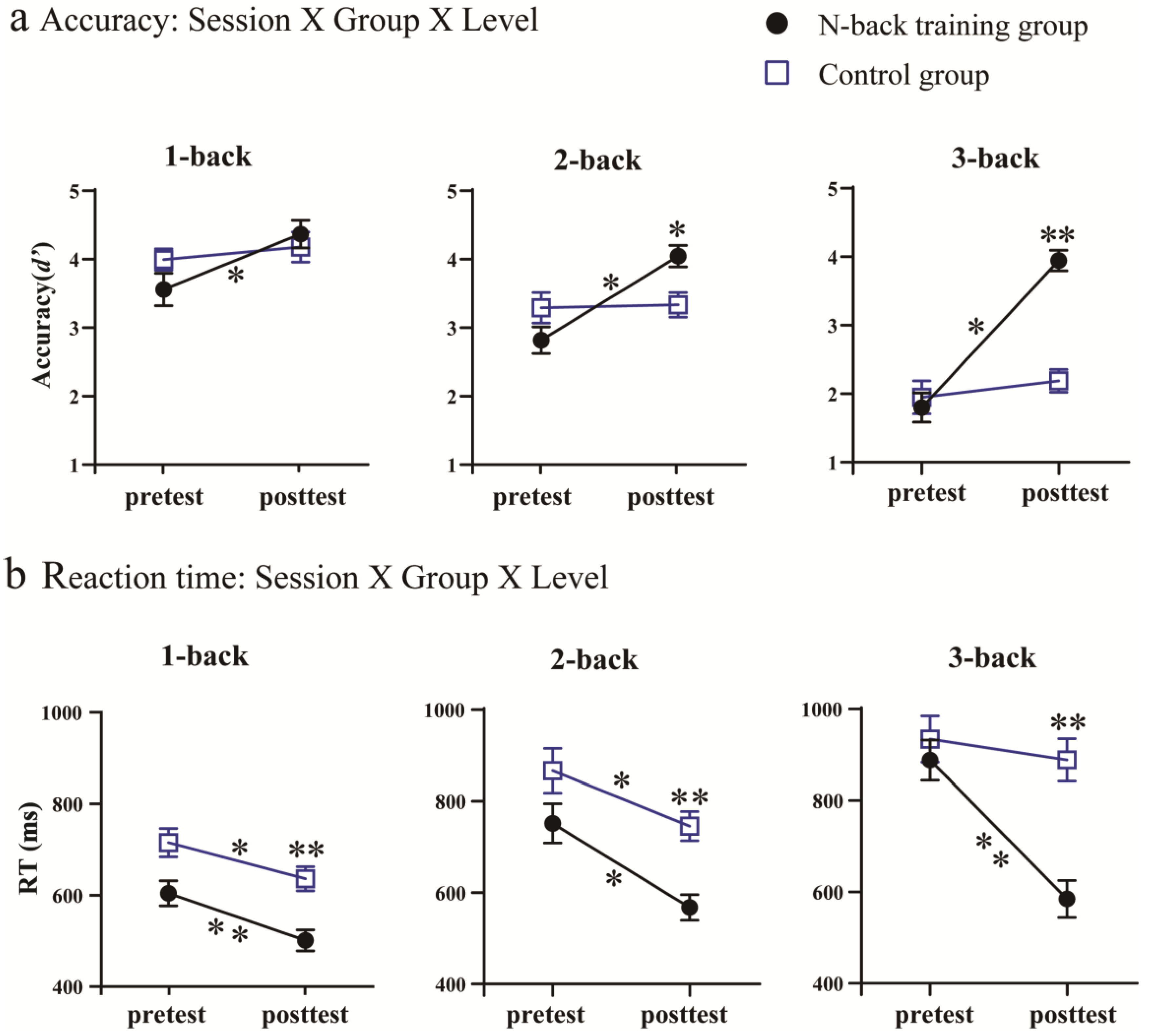
3.1. Behavioral Results
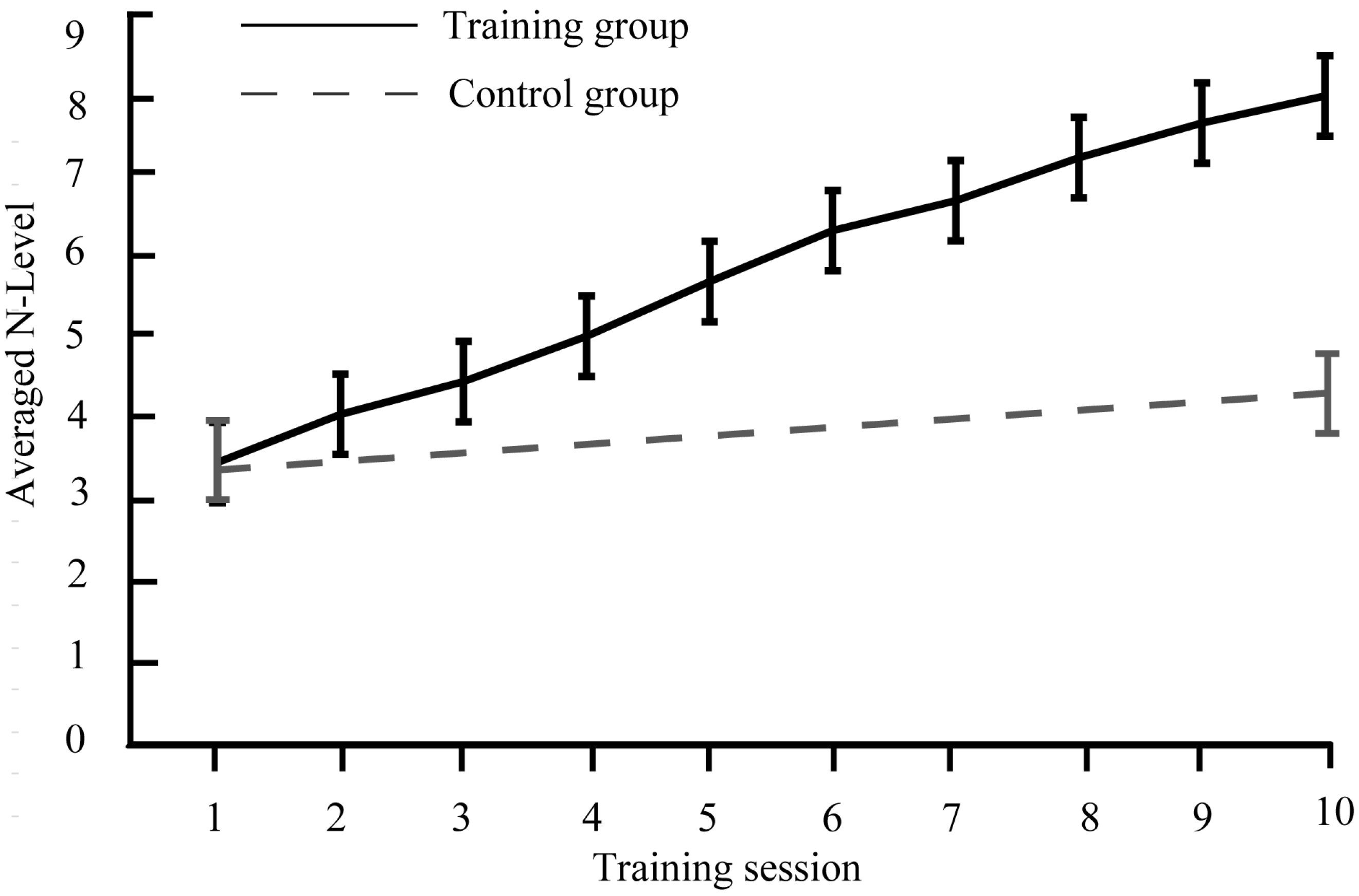
3.1.1. Training Outcomes
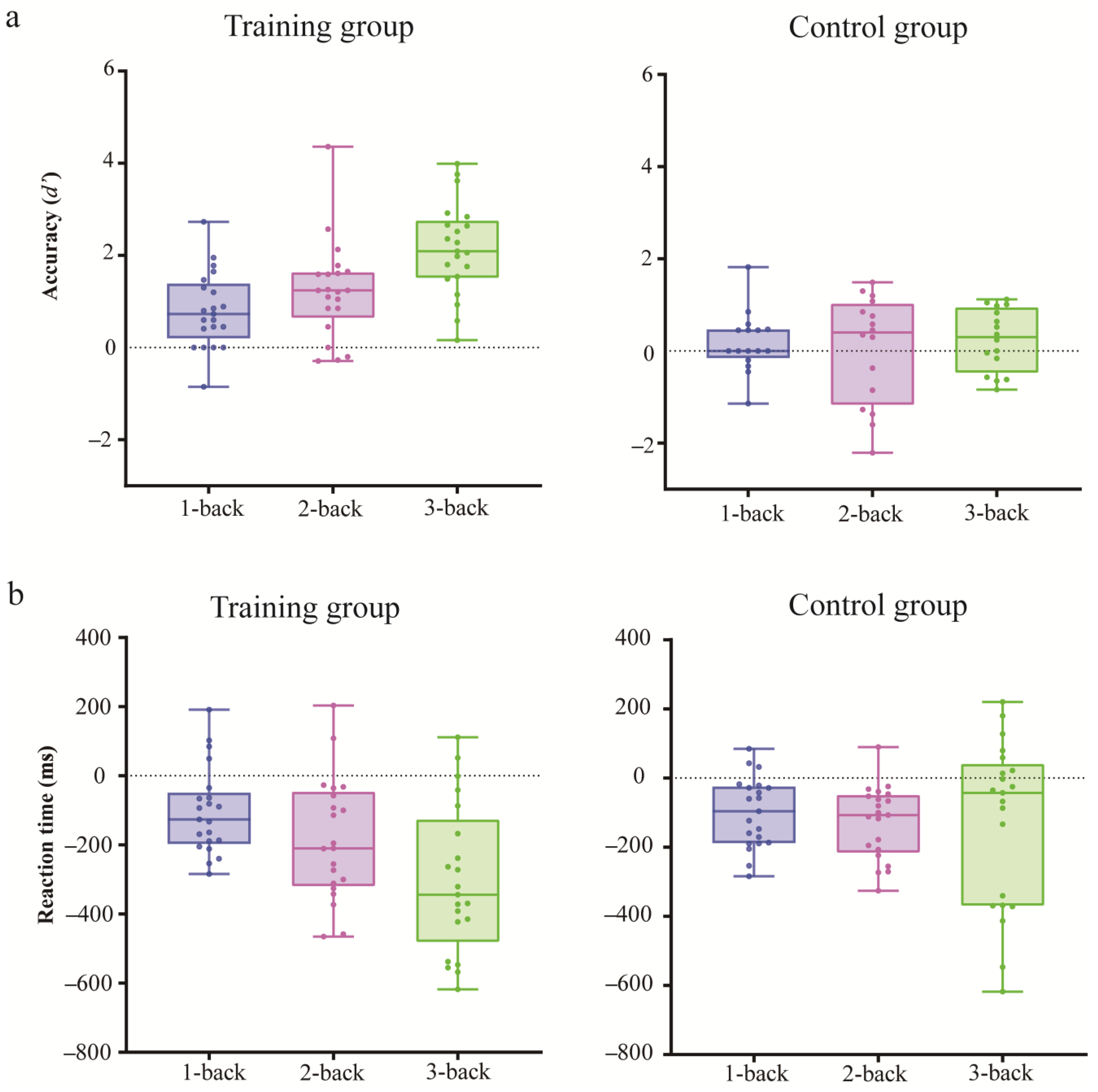
3.1.2. Training Gain
3.1.3. Transfer Outcomes
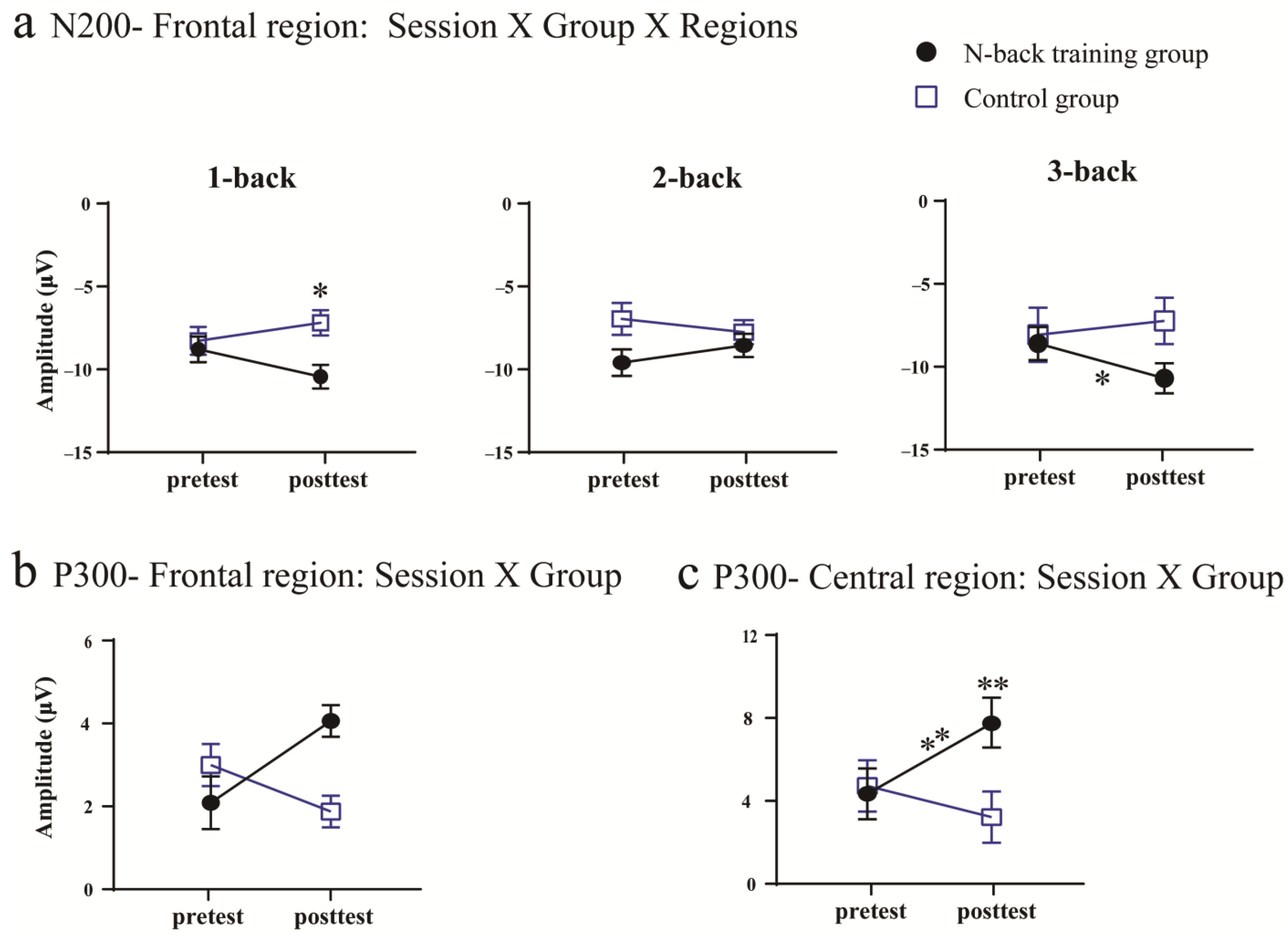
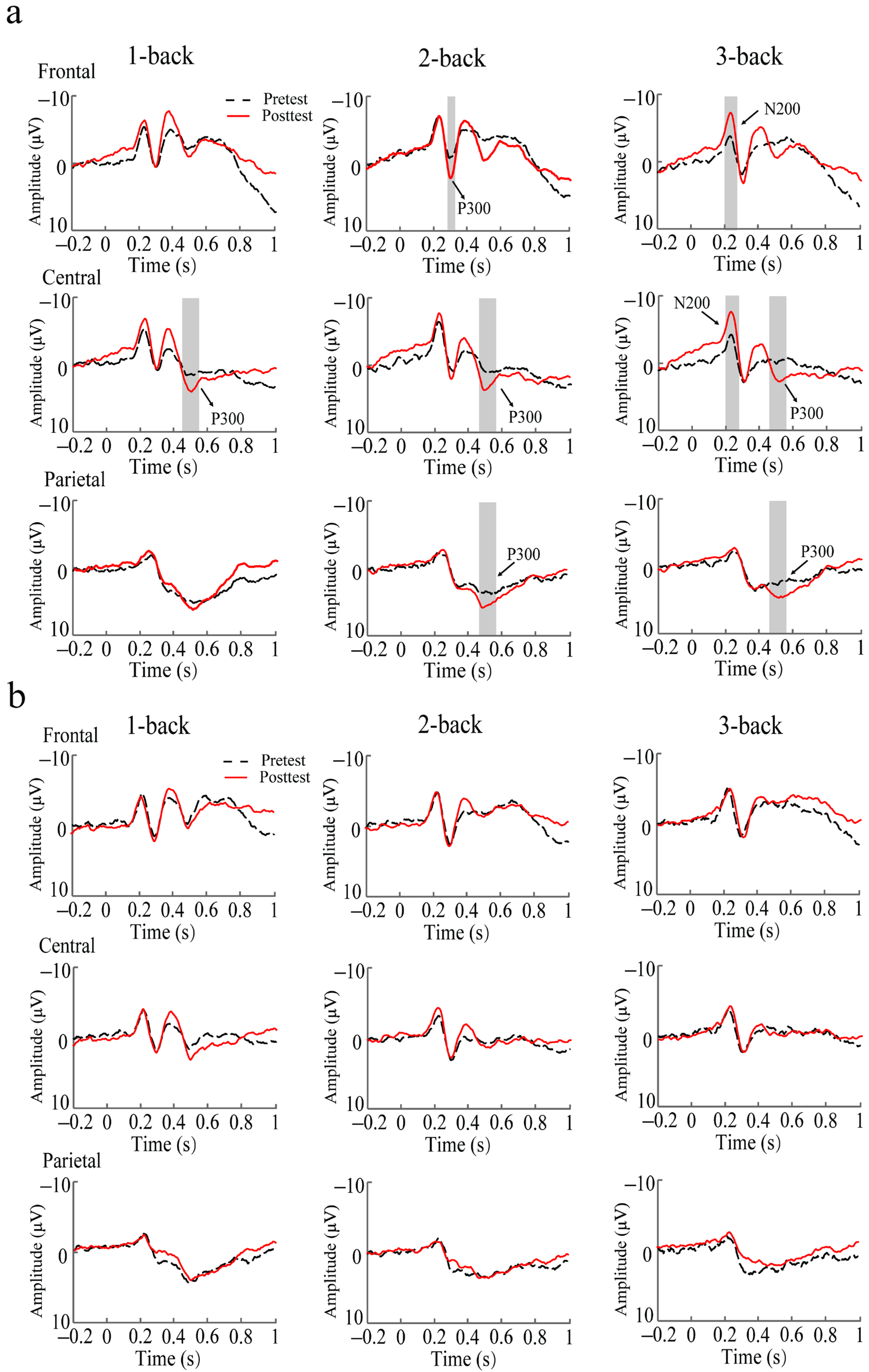
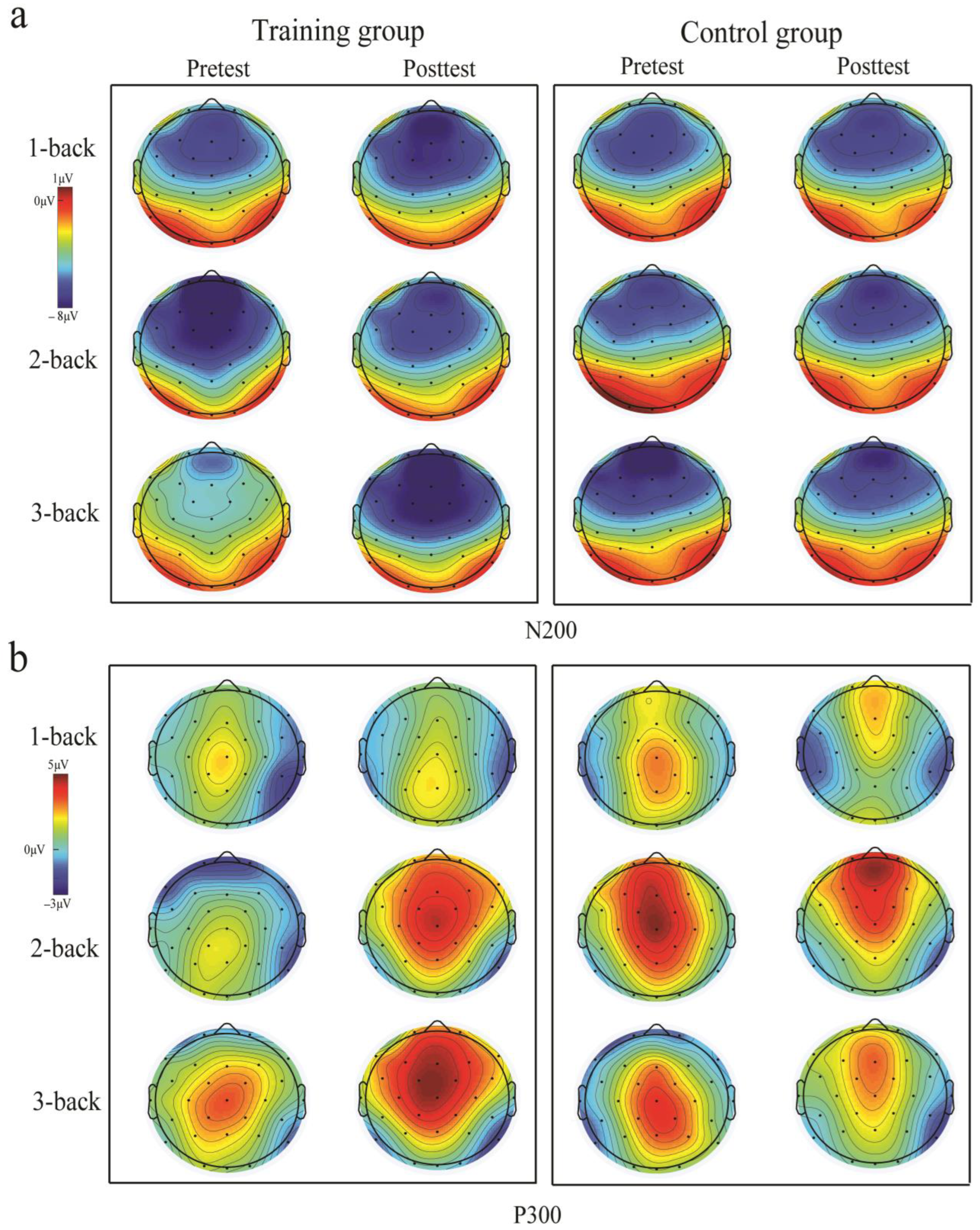
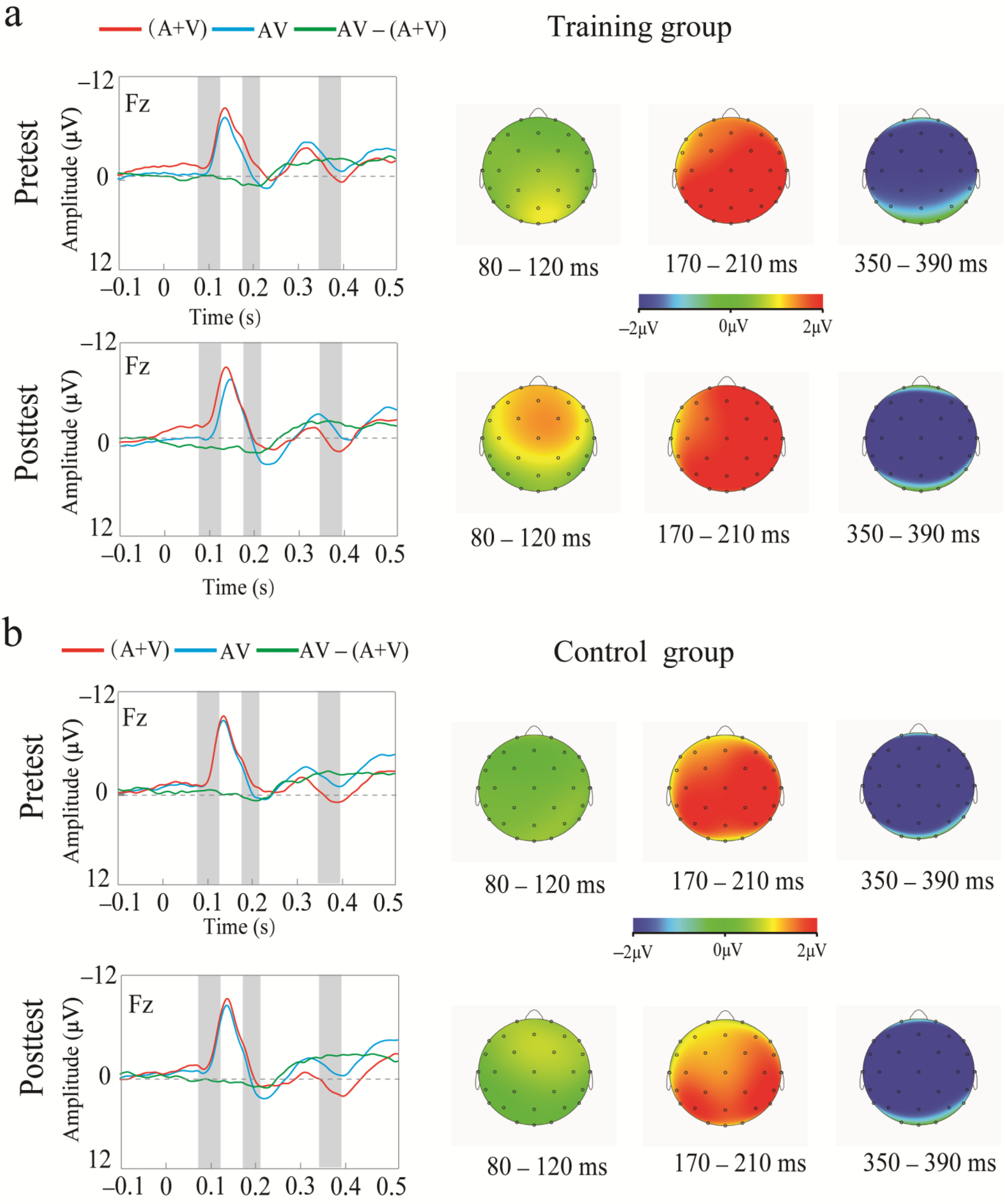
3.2. ERP Results
3.2.1. Training Outcomes
3.2.2. Transfer Outcome Measures
4. Discussion
4.1. Training Effect
4.1.1. Behavioral Performance
4.1.2. Neural Effects of Audiovisual n-Back Training
4.2. Transfer Effect
4.3. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baddeley, A. Working memory: Looking back and looking forward. Nat. Rev. Neurosci. 2003, 4, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Olesen, P.J.; Westerberg, H.; Klingberg, T. Increased prefrontal and parietal activity after training of working memory. Nat. Neurosci. 2004, 7, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Schneiders, J.A.; Opitz, B.; Tang, H.; Deng, Y.; Xie, C.; Li, H.; Mecklinger, A. The impact of auditory working memory training on the fronto-parietal working memory network. Front. Hum. Neurosci. 2012, 6, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneiders, J.A.; Opitz, B.; Krick, C.M.; Mecklinger, A. Separating intra-modal and across-modal training effects in visual working memory: An fMRI investigation. Cereb. Cortex 2011, 21, 2555–2564. [Google Scholar] [CrossRef]
- Frtusova, J.B.; Winneke, A.H.; Phillips, N.A. ERP evidence that auditory-visual speech facilitates working memory in younger and older adults. Psychol. Aging 2013, 28, 481–494. [Google Scholar] [CrossRef]
- Jaeggi, S.M.; Buschkuehl, M.; Jonides, J.; Perrig, W.J. Improving fluid intelligence with training on working memory. Proc. Natl. Acad. Sci. USA 2008, 105, 6829–6833. [Google Scholar] [CrossRef] [Green Version]
- Deveau, J.; Jaeggi, S.M.; Zordan, V.; Phung, C.; Seitz, A.R. How to build better memory training games. Front. Syst. Neurosci. 2015, 8, 243. [Google Scholar] [CrossRef]
- Jaeggi, S.M.; Studer-Luethi, B.; Buschkuehl, M.; Su, Y.-F.; Jonides, J.; Perrig, W.J. The relationship between n-back performance and matrix reasoning—Implications for training and transfer. Intelligence 2010, 38, 625–635. [Google Scholar] [CrossRef]
- Jaeggi, S.M.; Buschkuehl, M.; Shah, P.; Jonides, J. The role of individual differences in cognitive training and transfer. Mem. Cogn. 2014, 42, 464–480. [Google Scholar] [CrossRef]
- Laurienti, P.J.; Kraft, R.A.; Maldjian, J.A.; Burdette, J.H.; Wallace, M.T. Semantic congruence is a critical factor in multisensory behavioral performance. Exp. Brain Res. 2004, 158, 405–414. [Google Scholar] [CrossRef]
- Stein, B.E.; Stanford, T.R. Multisensory integration: Current issues from the perspective of the single neuron. Nat. Rev. Neurosci. 2008, 9, 255–266. [Google Scholar] [CrossRef]
- Giard, M.H.; Peronnet, F. Auditory-visual integration during multimodal object recognition in humans: A behavioral and electrophysiological study. J. Cogn. Neurosci. 1999, 11, 473–490. [Google Scholar] [CrossRef]
- Frtusova, J.B.; Phillips, N.A. The auditory-visual speech benefit on working memory in older adults with hearing impairment. Front. Psychol. 2016, 7, 490. [Google Scholar] [CrossRef] [Green Version]
- Leahy, W.; Sweller, J. Cognitive load theory, modality of presentation and the transient information effect. Appl. Cogn. Psychol. 2011, 25, 943–951. [Google Scholar] [CrossRef]
- Pahor, A.; Collins, C.; Smith-Peirce, R.N.; Moon, A.; Stavropoulos, T.; Silva, I.; Seitz, A.R. Multisensory Facilitation of Working Memory Training. J. Cogn. Enhanc. 2021, 5, 386–395. [Google Scholar] [CrossRef]
- Constantinidis, C.; Klingberg, T. The neuroscience of working memory capacity and training. Nat. Rev. Neurosci. 2016, 17, 438–449. [Google Scholar] [CrossRef]
- Xu, J.; Deng, M.; Nan, W.; Cai, D. The effects of working memory training in children revealed by behavioral responses and ERP. Brain Behav. 2021, 11, e2310. [Google Scholar] [CrossRef]
- Covey, T.J.; Shucard, J.L.; Shucard, D.W. Working memory training and perceptual discrimination training impact overlapping and distinct neurocognitive processes: Evidence from event-related potentials and transfer of training gains. Cognition 2019, 182, 50–72. [Google Scholar] [CrossRef]
- Folstein, J.R.; Van Petten, C. Influence of cognitive control and mismatch on the N2 component of the ERP: A review. Psychophysiology 2008, 45, 152–170. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Zhou, R.; Fu, L. Working memory updating function training influenced brain activity. PLoS ONE 2013, 8, e71063. [Google Scholar] [CrossRef]
- Cohen, J.R.; Sreenivasan, K.K.; D’Esposito, M. Correspondence between stimulus encoding-and maintenance-related neural processes underlies successful working memory. Cereb. Cortex 2014, 24, 593–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitada, R.; Sasaki, A.T.; Okamoto, Y.; Kochiyama, T.; Sadato, N. Role of the precuneus in the detection of incongruency between tactile and visual texture information: A functional MRI study. Neuropsychologia 2014, 64, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Polich, J. Updating P300: An integrative theory of P3a and P3b. Clin. Neurophysiol. 2007, 118, 2128–2148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, B.A.; Pichora-Fuller, M.K. Implications of perceptual deterioration for cognitive aging research. In Handbook of Aging and Cognition; Lawrence Erlbaum Associates: Mahwah, NJ, USA, 2000; pp. 155–219. [Google Scholar]
- Salminen, T.; Frensch, P.; Strobach, T.; Schubert, T. Age-specific differences of dual n-back training. Aging Neuropsychol. Cogn. 2016, 23, 18–39. [Google Scholar] [CrossRef] [PubMed]
- Au, J.; Sheehan, E.; Tsai, N.; Duncan, G.J.; Buschkuehl, M.; Jaeggi, S.M. Improving fluid intelligence with training on working memory: A meta-analysis. Psychon. Bull. Rev. 2015, 22, 366–377. [Google Scholar] [CrossRef] [Green Version]
- Bogg, T.; Lasecki, L. Reliable gains? Evidence for substantially underpowered designs in studies of working memory training transfer to fluid intelligence. Front. Psychol. 2015, 5, 1589. [Google Scholar] [CrossRef] [Green Version]
- Dahlin Neely, A.S.; Larsson, A.; Backman, L.; Nyberg, L. Transfer of learning after updating training mediated by the striatum. Science 2008, 320, 1510–1512. [Google Scholar] [CrossRef] [Green Version]
- Cowan, N.; Li, D.; Moffitt, A.; Becker, T.M.; Martin, E.A.; Saults, J.S.; Christ, S.E. A neural region of abstract working memory. J. Cogn. Neurosci. 2011, 23, 2852–2863. [Google Scholar] [CrossRef] [Green Version]
- von Bastian, C.C.; Oberauer, K. Effects and mechanisms of working memory training: A review. Psychol. Res. 2014, 78, 803–820. [Google Scholar] [CrossRef]
- Chen, Y.C.; Yeh, S.L.; Tang, P.F. Age-related changes in audiovisual simultaneity perception and their relationship with working memory. J. Gerontol. Ser. B 2021, 76, 1095–1103. [Google Scholar] [CrossRef]
- Baum, S.H.; Stevenson, R.A. Shifts in audiovisual processing in healthy aging. Curr. Behav. Neurosci. Rep. 2017, 4, 198–208. [Google Scholar] [CrossRef]
- Snodgrass, J.G.; Vanderwart, M. A standardized set of 260 pictures: Norms for name agreement, image agreement, familiarity, and visual complexity. J. Exp. Psychol. Hum. Learn. Mem. 1980, 6, 174. [Google Scholar] [CrossRef]
- Shalchy, M.A.; Pergher, V.; Pahor, A.; Van Hulle, M.M.; Seitz, A.R. N-Back Related ERPs Depend on Stimulus Type, Task Structure, Pre-processing, and Lab Factors. Front. Hum. Neurosci. 2020, 14, 549966. [Google Scholar] [CrossRef]
- Peng, Z.; Dai, C.; Ba, Y.; Zhang, L.; Shao, Y.; Tian, J. Effect of sleep deprivation on the working memory-related N2-P3 components of the event-related potential waveform. Front. Neurosci. 2020, 14, 469. [Google Scholar] [CrossRef]
- Guthrie, D.; Buchwald, J.S. Significance testing of difference potentials. Psychophysiology 1991, 28, 240–244. [Google Scholar] [CrossRef]
- Lawlor-Savage, L.; Goghari, V.M. Dual N-back working memory training in healthy adults: A randomized comparison to processing speed training. PLoS ONE 2016, 11, e151817. [Google Scholar] [CrossRef]
- Küper, K.; Karbach, J. Increased training complexity reduces the effectiveness of brief working memory training: Evidence from short-term single and dual n-back training interventions. J. Cogn. Psychol. 2016, 28, 199–208. [Google Scholar] [CrossRef]
- Regenbogen, C.; Seubert, J.; Johansson, E.; Finkelmeyer, A.; Andersson, P.; Lundström, J.N. The intraparietal sulcus governs multisensory integration of audiovisual information based on task difficulty. Hum. Brain Mapp. 2018, 39, 1313–1326. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Guo, Z.; Wang, X.; Sun, K.; Lin, X.; Wang, X.; Zhang, J. Effects of Audiovisual Interactions on Working Memory Task Performance—Interference or Facilitation. Brain Sci. 2022, 12, 886. [Google Scholar] [CrossRef]
- Pergher, V.; Wittevrongel, B.; Tournoy, J.; Schoenmakers, B.; Van Hulle, M.M. N-back training and transfer effects revealed by behavioral responses and EEG. Brain Behav. 2018, 8, e01136. [Google Scholar] [CrossRef] [Green Version]
- Collette, F.; Van der Linden, M. Brain imaging of the central executive component of working memory. Neurosci. Biobehav. Rev. 2002, 26, 105–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen, A.M.; McMillan, K.M.; Laird, A.R.; Bullmore, E. N-back working memory paradigm: A meta-analysis of normative functional neuroimaging studies. Hum. Brain Mapp. 2005, 25, 46–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, O.; Lisman, J.E. Hippocampal sequence-encoding driven by a cortical multi-item working memory buffer. Trends Neurosci. 2005, 28, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Moran, Z.D.; Bachman, P.; Pham, P.; Cho, S.H.; Cannon, T.D.; Shams, L. Multisensory encoding improves auditory recognition. Multisens. Res. 2013, 26, 581–592. [Google Scholar] [CrossRef] [Green Version]
- Oelhafen, S.; Nikolaidis, A.; Padovani, T.; Blaser, D.; Koenig, T.; Perrig, W.J. Increased parietal activity after training of interference control. Neuropsychologia 2013, 51, 2781–2790. [Google Scholar] [CrossRef]
- Schleepen, T.M.; Jonkman, L.M. The development of non-spatial working memory capacity during childhood and adolescence and the role of interference control: An N-Back task study. Dev. Neuropsychol. 2009, 35, 37–56. [Google Scholar] [CrossRef]
- Smith, E.E.; Jonides, J. Working memory: A view from neuroimaging. Cogn. Psychol. 1997, 33, 5–42. [Google Scholar] [CrossRef] [Green Version]
- Diamond, A. Normal development of prefrontal cortex from birth to young adulthood: Cognitive functions, anatomy, and biochemistry. Princ. Front. Lobe Funct. 2002, 466, 503. [Google Scholar]
- Talsma, D. Predictive coding and multisensory integration: An attentional account of the multisensory mind. Front. Integr. Neurosci. 2015, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Michail, G.; Senkowski, D.; Niedeggen, M.; Keil, J. Memory load alters perception-related neural oscillations during multisensory integration. J. Neurosci. 2021, 41, 1505–1515. [Google Scholar] [CrossRef]
- Baart, M.; Stekelenburg, J.J.; Vroomen, J. Electrophysiological evidence for speech-specific audiovisual integration. Neuropsychologia 2014, 53, 115–121. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Auditory | Visual | Audiovisual | ||||
|---|---|---|---|---|---|---|
| Pre-Test | Post-Test | Pre-Test | Post-Test | Pre-Test | Post-Test | |
| Training group | ||||||
| Accuracy (%) | 76 (20) | 87 (13) | 76 (19) | 83 (14) | 94 (6) | 95 (6) |
| RT (ms) | 420 (34) | 413 (28) | 416 (27) | 415 (25) | 382 (37) | 373 (29) |
| Control group | ||||||
| Accuracy (%) | 74 (25) | 77 (17) | 72 (20) | 73 (18) | 92 (9) | 95 (4) |
| RT (ms) | 424 (38) | 424 (33) | 422 (31) | 423 (24) | 394 (43) | 386 (39) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, A.; Yang, W.; Yang, X.; Lin, J.; Li, Z.; Ren, Y.; Yang, J.; Wu, J. Audiovisual n-Back Training Alters the Neural Processes of Working Memory and Audiovisual Integration: Evidence of Changes in ERPs. Brain Sci. 2023, 13, 992. https://doi.org/10.3390/brainsci13070992
Guo A, Yang W, Yang X, Lin J, Li Z, Ren Y, Yang J, Wu J. Audiovisual n-Back Training Alters the Neural Processes of Working Memory and Audiovisual Integration: Evidence of Changes in ERPs. Brain Sciences. 2023; 13(7):992. https://doi.org/10.3390/brainsci13070992
Chicago/Turabian StyleGuo, Ao, Weiping Yang, Xiangfu Yang, Jinfei Lin, Zimo Li, Yanna Ren, Jiajia Yang, and Jinglong Wu. 2023. "Audiovisual n-Back Training Alters the Neural Processes of Working Memory and Audiovisual Integration: Evidence of Changes in ERPs" Brain Sciences 13, no. 7: 992. https://doi.org/10.3390/brainsci13070992
APA StyleGuo, A., Yang, W., Yang, X., Lin, J., Li, Z., Ren, Y., Yang, J., & Wu, J. (2023). Audiovisual n-Back Training Alters the Neural Processes of Working Memory and Audiovisual Integration: Evidence of Changes in ERPs. Brain Sciences, 13(7), 992. https://doi.org/10.3390/brainsci13070992