
Alzheimer’s Disease: Insights from Large-Scale Brain Dynamics Models



Abstract
:1. Introduction
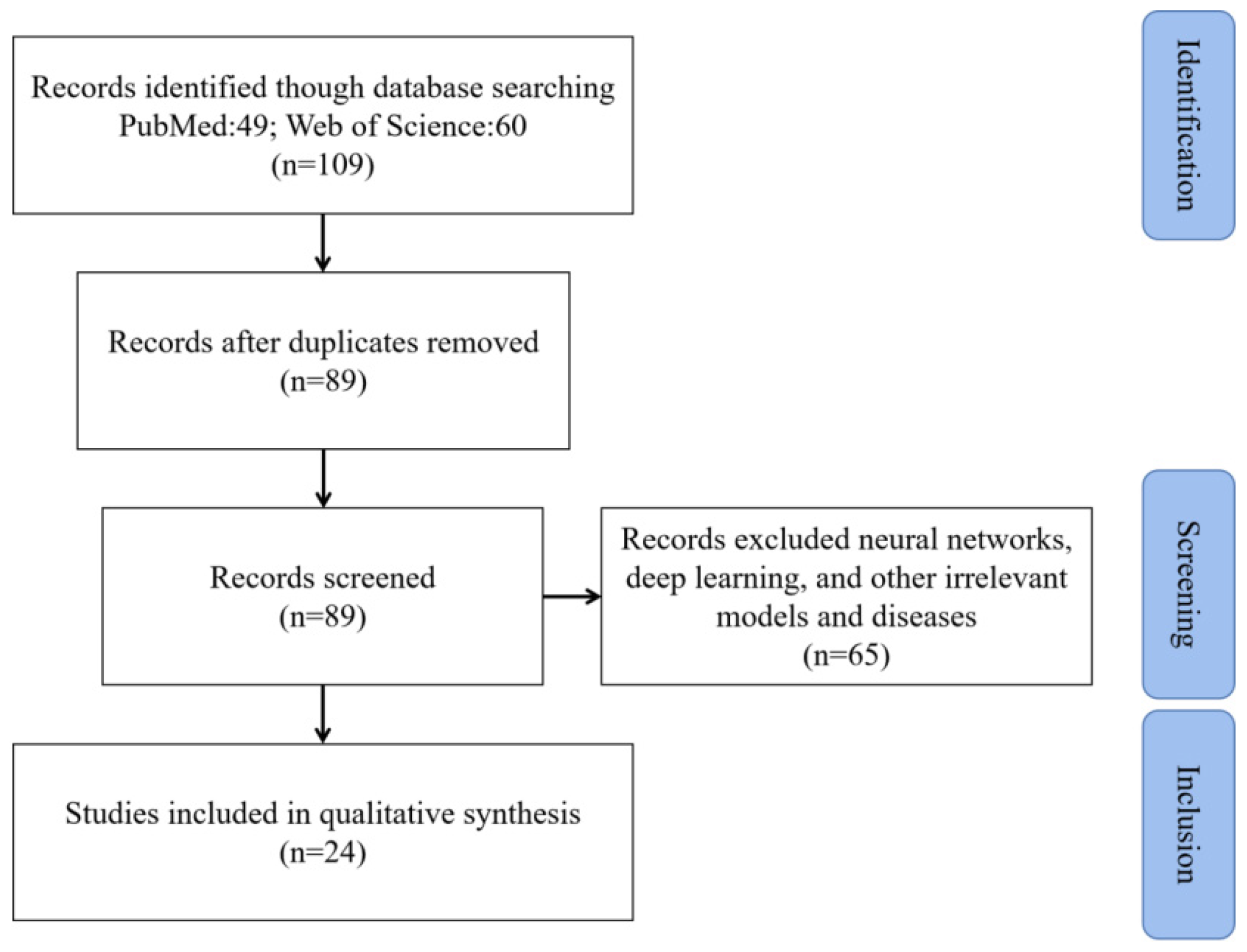
2. Materials and Methods
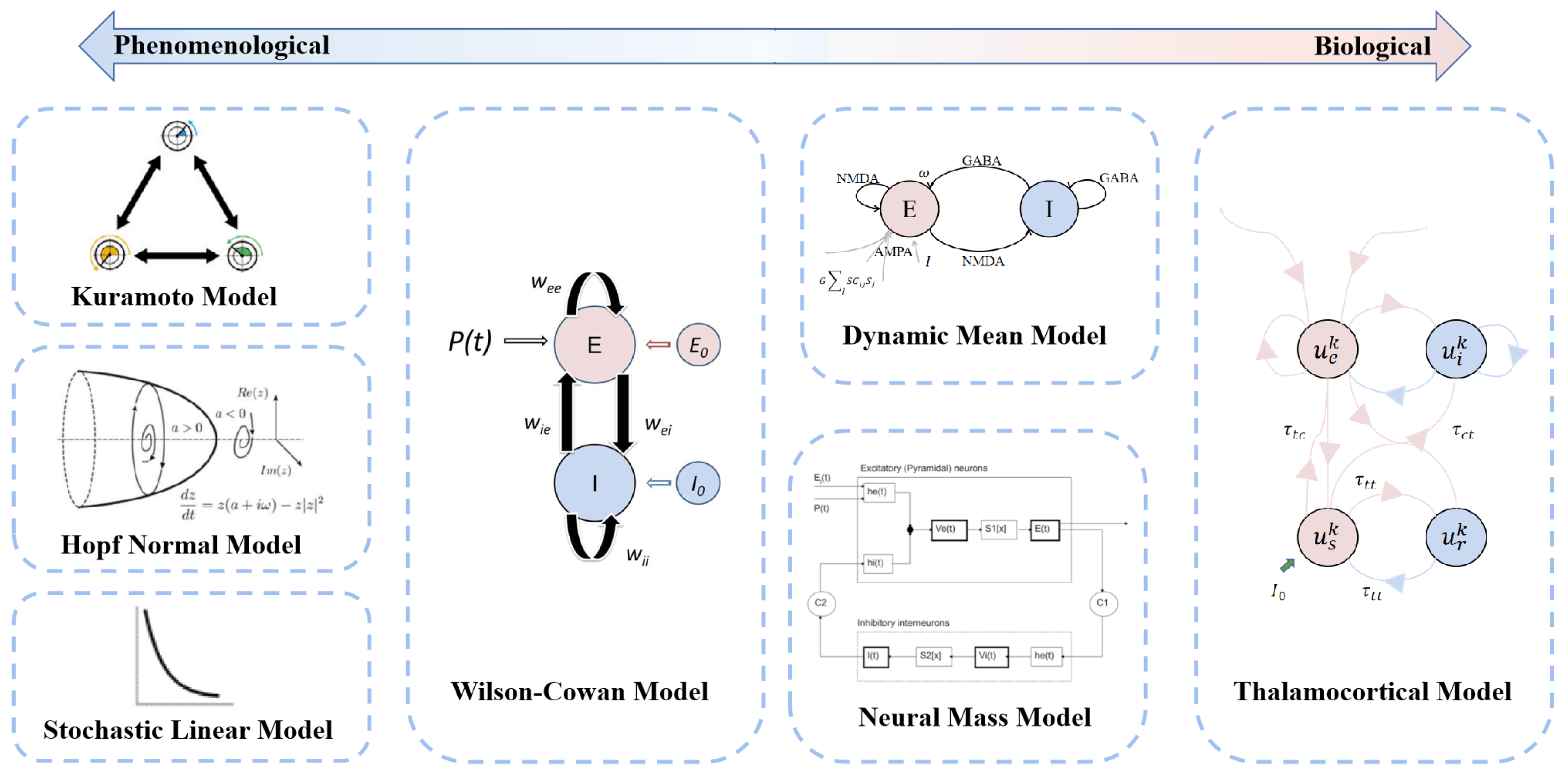
3. Large-Scale Brain Dynamics Models
3.1. Neural Dynamics Models
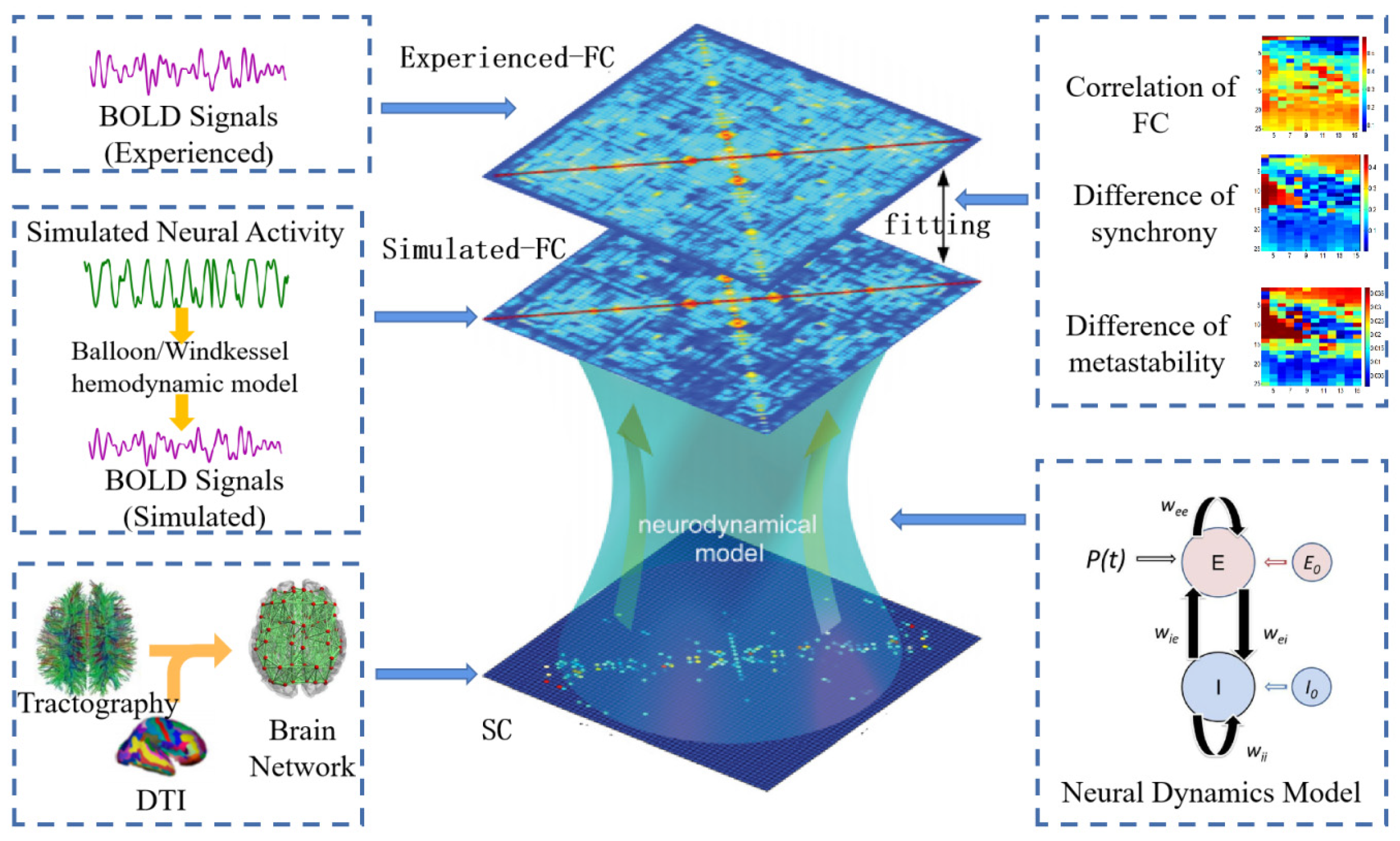
3.2. Large-Scale Dynamics Model Building
4. Applications of Large-Scale Brain Dynamics Models to Alzheimer’s Disease
4.1. Slowing of Alpha Rhythm: A Biomarker for AD
4.2. Altering Neuronal Excitability: A Therapeutic Strategy for AD
4.3. Abnormal Regions of the Brain: Potential Therapeutic Target Areas
5. Future Directions
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ackleh, A.S.; Elaydi, S.; Livadiotis, G.; Veprauskas, A. A continuous-time mathematical model and discrete approximations for the aggregation of beta-Amyloid. J. Biol. Dyn. 2021, 15, 109–136. [Google Scholar] [CrossRef] [PubMed]
- Jack, C.J.; Knopman, D.S.; Jagust, W.J.; Petersen, R.C.; Weiner, M.W.; Aisen, P.S.; Shaw, L.M.; Vemuri, P.; Wiste, H.J.; Weigand, S.D.; et al. Tracking pathophysiological processes in Alzheimer’s disease: An updated hypothetical model of dynamic biomarkers. Lancet. Neurol. 2013, 12, 207–216. [Google Scholar] [CrossRef] [Green Version]
- D’Amelio, M.; Rossini, P.M. Brain excitability and connectivity of neuronal assemblies in Alzheimer’s disease: From animal models to human findings. Prog. Neurobiol. 2012, 99, 42–60. [Google Scholar] [CrossRef] [PubMed]
- Pons, V.; Rivest, S. Targeting Systemic Innate Immune Cells as a Therapeutic Avenue for Alzheimer Disease. Pharm. Rev. 2022, 74, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Hampel, H.; Toschi, N.; Babiloni, C.; Baldacci, F.; Black, K.L.; Bokde, A.; Bun, R.S.; Cacciola, F.; Cavedo, E.; Chiesa, P.A.; et al. Revolution of Alzheimer Precision Neurology. Passageway of Systems Biology and Neurophysiology. J. Alzheimers Dis. 2018, 64, S47–S105. [Google Scholar] [CrossRef]
- Bachmann, C.; Tetzlaff, T.; Duarte, R.; Morrison, A. Firing rate homeostasis counteracts changes in stability of recurrent neural networks caused by synapse loss in Alzheimer’s disease. PLoS Comput. Biol. 2020, 16, e1007790. [Google Scholar] [CrossRef]
- Tampellini, D. Synaptic activity and Alzheimer’s disease: A critical update. Front. Neurosci. 2015, 9, 423. [Google Scholar] [CrossRef] [Green Version]
- Melgosa-Ecenarro, L.; Doostdar, N.; Radulescu, C.I.; Jackson, J.S.; Barnes, S.J. Pinpointing the locus of GABAergic vulnerability in Alzheimer’s disease. Semin. Cell Dev. Biol. 2023, 139, 35–54. [Google Scholar] [CrossRef]
- Serrano, M.E.; Kim, E.; Petrinovic, M.M.; Turkheimer, F.; Cash, D. Imaging Synaptic Density: The Next Holy Grail of Neuroscience? Front. Neurosci. 2022, 16, 796129. [Google Scholar] [CrossRef]
- Mandal, P.K.; Banerjee, A.; Tripathi, M.; Sharma, A. A Comprehensive Review of Magnetoencephalography (MEG) Studies for Brain Functionality in Healthy Aging and Alzheimer’s Disease (AD). Front. Comput. Neurosci. 2018, 12, 60. [Google Scholar] [CrossRef]
- Cassani, R.; Estarellas, M.; San-Martin, R.; Fraga, F.J.; Falk, T.H. Systematic Review on Resting-State EEG for Alzheimer’s Disease Diagnosis and Progression Assessment. Dis. Markers 2018, 2018, 5174815. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Kong, M.; Zhang, C.; Guan, L.; Ba, M. The structural MRI markers and cognitive decline in prodromal Alzheimer’s disease: A 2-year longitudinal study. Quant Imaging Med. Surg. 2018, 8, 1004–1019. [Google Scholar] [CrossRef] [PubMed]
- Chandra, A.; Dervenoulas, G.; Politis, M. Magnetic resonance imaging in Alzheimer’s disease and mild cognitive impairment. J. Neurol. 2019, 266, 1293–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talwar, P.; Kushwaha, S.; Chaturvedi, M.; Mahajan, V. Systematic Review of Different Neuroimaging Correlates in Mild Cognitive Impairment and Alzheimer’s Disease. Clin. Neuroradiol. 2021, 31, 953–967. [Google Scholar] [CrossRef] [PubMed]
- Maestu, F.; Cuesta, P.; Hasan, O.; Fernandez, A.; Funke, M.; Schulz, P.E. The Importance of the Validation of M/EEG With Current Biomarkers in Alzheimer’s Disease. Front. Hum. Neurosci. 2019, 13, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, A.; Szucs, A.; Hidasi, Z.; Csukly, G.; Barcs, G.; Kamondi, A. Prevalence, Semiology, and Risk Factors of Epilepsy in Alzheimer’s Disease: An Ambulatory EEG Study. J. Alzheimers Dis. 2018, 63, 1045–1054. [Google Scholar] [CrossRef]
- Li, H.J.; Hou, X.H.; Liu, H.H.; Yue, C.L.; He, Y.; Zuo, X.N. Toward systems neuroscience in mild cognitive impairment and Alzheimer’s disease: A meta-analysis of 75 fMRI studies. Hum. Brain Mapp. 2015, 36, 1217–1232. [Google Scholar] [CrossRef]
- Maestu, F.; de Haan, W.; Busche, M.A.; DeFelipe, J. Neuronal excitation/inhibition imbalance: Core element of a translational perspective on Alzheimer pathophysiology. Ageing Res. Rev. 2021, 69, 101372. [Google Scholar] [CrossRef]
- Breakspear, M. Dynamic models of large-scale brain activity. Nat. Neurosci. 2017, 20, 340–352. [Google Scholar] [CrossRef]
- Popovych, O.V.; Manos, T.; Hoffstaedter, F.; Eickhoff, S.B. What Can Computational Models Contribute to Neuroimaging Data Analytics? Front. Syst. Neurosci. 2018, 12, 68. [Google Scholar] [CrossRef] [Green Version]
- Bansal, K.; Nakuci, J.; Muldoon, S.F. Personalized brain network models for assessing structure-function relationships. Curr. Opin. Neurobiol. 2018, 52, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Honey, C.J.; Sporns, O.; Cammoun, L.; Gigandet, X.; Thiran, J.P.; Meuli, R.; Hagmann, P. Predicting human resting-state functional connectivity from structural connectivity. Proc. Natl. Acad. Sci. USA 2009, 106, 2035–2040. [Google Scholar] [CrossRef] [PubMed]
- Deco, G.; Tononi, G.; Boly, M.; Kringelbach, M.L. Rethinking segregation and integration: Contributions of whole-brain modelling. Nat. Rev. Neurosci. 2015, 16, 430–439. [Google Scholar] [PubMed]
- Cabral, J.; Vidaurre, D.; Marques, P.; Magalhaes, R.; Silva, M.P.; Miguel, S.J.; Deco, G.; Sousa, N.; Kringelbach, M.L. Cognitive performance in healthy older adults relates to spontaneous switching between states of functional connectivity during rest. Sci. Rep. 2017, 7, 5135. [Google Scholar] [PubMed] [Green Version]
- Lee, H.; Golkowski, D.; Jordan, D.; Berger, S.; Ilg, R.; Lee, J.; Mashour, G.A.; Lee, U. Relationship of critical dynamics, functional connectivity, and states of consciousness in large-scale human brain networks. Neuroimage 2019, 188, 228–238. [Google Scholar] [CrossRef]
- Deco, G.; Kringelbach, M.L. Great expectations: Using whole-brain computational connectomics for understanding neuropsychiatric disorders. Neuron 2014, 84, 892–905. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, G.C.; Tewarie, P.; Vidaurre, D.; Liuzzi, L.; Woolrich, M.W.; Brookes, M.J. Dynamics of large-scale electrophysiological networks: A technical review. Neuroimage 2018, 180, 559–576. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, T.T.; Jirsa, V.K.; Spiegler, A.; McIntosh, A.R.; Deco, G. Bottom up modeling of the connectome: Linking structure and function in the resting brain and their changes in aging. Neuroimage 2013, 80, 318–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naik, S.; Banerjee, A.; Bapi, R.S.; Deco, G.; Roy, D. Metastability in Senescence. Trends Cogn. Sci. 2017, 21, 509–521. [Google Scholar]
- Sase, T.; Kitajo, K. The metastable brain associated with autistic-like traits of typically developing individuals. PLoS Comput. Biol. 2021, 17, e1008929. [Google Scholar]
- Alderson, T.H.; Bokde, A.; Kelso, J.; Maguire, L.; Coyle, D. Metastable neural dynamics in Alzheimer’s disease are disrupted by lesions to the structural connectome. Neuroimage 2018, 183, 438–455. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yang, X.; Sun, Z. Alpha rhythm slowing in a modified thalamo-cortico-thalamic model related with Alzheimer’s disease. PLoS ONE 2020, 15, e229950. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Gomez, S.J.; Hornero, R.; Poza, J.; Maturana-Candelas, A.; Pinto, N.; Gomez, C. Computational modeling of the effects of EEG volume conduction on functional connectivity metrics. Application to Alzheimer’s disease continuum. J. Neural. Eng. 2019, 16, 66019. [Google Scholar] [CrossRef] [PubMed]
- Stefanovski, L.; Triebkorn, P.; Spiegler, A.; Diaz-Cortes, M.A.; Solodkin, A.; Jirsa, V.; McIntosh, A.R.; Ritter, P. Linking Molecular Pathways and Large-Scale Computational Modeling to Assess Candidate Disease Mechanisms and Pharmacodynamics in Alzheimer’s Disease. Front. Comput. Neurosci. 2019, 13, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demirtas, M.; Falcon, C.; Tucholka, A.; Gispert, J.D.; Molinuevo, J.L.; Deco, G. A whole-brain computational modeling approach to explain the alterations in resting-state functional connectivity during progression of Alzheimer’s disease. Neuroimage Clin. 2017, 16, 343–354. [Google Scholar] [CrossRef]
- Das, S.; Puthankattil, S.D. Functional Connectivity and Complexity in the Phenomenological Model of Mild Cognitive-Impaired Alzheimer’s Disease. Front. Comput. Neurosci. 2022, 16, 877912. [Google Scholar] [CrossRef]
- de Haan, W.; Mott, K.; van Straaten, E.C.; Scheltens, P.; Stam, C.J. Activity dependent degeneration explains hub vulnerability in Alzheimer’s disease. PLoS Comput. Biol. 2012, 8, e1002582. [Google Scholar] [CrossRef] [Green Version]
- Ghorbanian, P.; Ramakrishnan, S.; Ashrafiuon, H. Stochastic coupled oscillator model of EEG for Alzheimer’s disease. In Proceedings of the 2014 36th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Chicago, IL, USA, 26–30 August 2014; pp. 706–709. [Google Scholar]
- de Haan, W.; van Straaten, E.; Gouw, A.A.; Stam, C.J. Altering neuronal excitability to preserve network connectivity in a computational model of Alzheimer’s disease. PLoS Comput. Biol. 2017, 13, e1005707. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, J.; Perry, A.; Breakspear, M.; Schirner, M.; Sachdev, P.; Wen, W.; Kochan, N.A.; Mapstone, M.; Ritter, P.; McIntosh, A.R.; et al. Differentiation of Alzheimer’s disease based on local and global parameters in personalized Virtual Brain models. Neuroimage Clin. 2018, 19, 240–251. [Google Scholar] [CrossRef]
- Cakir, Y. Hybrid modeling of alpha rhythm and the amplitude of low-frequency fluctuations abnormalities in the thalamocortical region and basal ganglia in Alzheimer’s disease. Eur. J. Neurosci. 2020, 52, 2944–2961. [Google Scholar] [CrossRef]
- Arbabyazd, L.; Shen, K.; Wang, Z.; Hofmann-Apitius, M.; Ritter, P.; McIntosh, A.R.; Battaglia, D.; Jirsa, V. Virtual Connectomic Datasets in Alzheimer’s Disease and Aging Using Whole-Brain Network Dynamics Modelling. Eneuro 2021, 8. [Google Scholar] [CrossRef]
- Triebkorn, P.; Stefanovski, L.; Dhindsa, K.; Diaz-Cortes, M.A.; Bey, P.; Bulau, K.; Pai, R.; Spiegler, A.; Solodkin, A.; Jirsa, V.; et al. Brain simulation augments machine-learning-based classification of dementia. Alzheimers Dement 2022, 8, e12303. [Google Scholar] [CrossRef] [PubMed]
- van Nifterick, A.M.; Gouw, A.A.; van Kesteren, R.E.; Scheltens, P.; Stam, C.J.; de Haan, W. A multiscale brain network model links Alzheimer’s disease-mediated neuronal hyperactivity to large-scale oscillatory slowing. Alzheimers Res. 2022, 14, 101. [Google Scholar] [CrossRef] [PubMed]
- Patow, G.; Stefanovski, L.; Ritter, P.; Deco, G.; Kobeleva, X. Whole-brain modeling of the differential influences of Amyloid-Beta and Tau in Alzheimer’s Disease. bioRxiv 2023, XXI, 1–5. [Google Scholar]
- Salimi-Nezhad, N.; Missault, S.; Notario-Reinoso, A.; Hassani, A.; Amiri, M.; Keliris, G.A. The impact of selective and non-selective medial septum stimulation on hippocampal neuronal oscillations: A study based on modeling and experiments. Neurobiol. Dis. 2023, 180, 106052. [Google Scholar] [CrossRef]
- Sanz, P.Y.; Fittipaldi, S.; Gonzalez, C.C.; Moguilner, S.; Cruzat, J.; Fraile-Vazquez, M.E.; Herzog, R.; Kringelbach, M.L.; Deco, G.; Prado, P.; et al. Model-based whole-brain perturbational landscape of neurodegenerative diseases. Elife 2023, 12. [Google Scholar]
- Alexandersen, C.G.; de Haan, W.; Bick, C.; Goriely, A. A multi-scale model explains oscillatory slowing and neuronal hyperactivity in Alzheimer’s disease. J. R. Soc. Interface 2023, 20, 20220607. [Google Scholar] [CrossRef]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gotzsche, P.C.; Ioannidis, J.P.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate healthcare interventions: Explanation and elaboration. BMJ 2009, 339, b2700. [Google Scholar] [CrossRef] [Green Version]
- HODGKINA, L.; HUXLEYA, F. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 1952, 117, 500–544. [Google Scholar] [CrossRef] [PubMed]
- Wilson, H.R.; Cowan, J.D. Excitatory and inhibitory interactions in localized populations of model neurons. Biophys. J. 1972, 12, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Pathak, A.; Roy, D.; Banerjee, A. Whole-Brain Network Models: From Physics to Bedside. Front. Comput. Neurosci. 2022, 16, 866517. [Google Scholar] [CrossRef] [PubMed]
- Amari, S. Dynamics of pattern formation in lateral-inhibition type neural fields. Biol. Cybern. 1977, 27, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Deco, G.; Jirsa, V.; McIntosh, A.R.; Sporns, O.; Kotter, R. Key role of coupling, delay, and noise in resting brain fluctuations. Proc Natl. Acad. Sci. USA 2009, 106, 10302–10307. [Google Scholar] [CrossRef] [PubMed]
- Cofre, R.; Herzog, R.; Mediano, P.; Piccinini, J.; Rosas, F.E.; Sanz, P.Y.; Tagliazucchi, E. Whole-Brain Models to Explore Altered States of Consciousness from the Bottom Up. Brain Sci. 2020, 10, 626. [Google Scholar] [CrossRef]
- Cabral, J.; Hugues, E.; Sporns, O.; Deco, G. Role of local network oscillations in resting-state functional connectivity. Neuroimage 2011, 57, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Breakspear, M.; Heitmann, S.; Daffertshofer, A. Generative models of cortical oscillations: Neurobiological implications of the kuramoto model. Front. Hum. Neurosci. 2010, 4, 190. [Google Scholar] [CrossRef] [Green Version]
- Lord, L.D.; Stevner, A.B.; Deco, G.; Kringelbach, M.L. Understanding principles of integration and segregation using whole-brain computational connectomics: Implications for neuropsychiatric disorders. Philos. Trans. A Math. Phys. Eng. Sci. 2017, 375. [Google Scholar]
- Saggio, M.L.; Ritter, P.; Jirsa, V.K. Analytical Operations Relate Structural and Functional Connectivity in the Brain. PLoS ONE 2016, 11, e157292. [Google Scholar]
- Deco, G.; Ponce-Alvarez, A.; Hagmann, P.; Romani, G.L.; Mantini, D.; Corbetta, M. How local excitation-inhibition ratio impacts the whole brain dynamics. J. Neurosci. 2014, 34, 7886–7898. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, J.D.; McIntosh, A.R.; Lefebvre, J. A Connectome-Based, Corticothalamic Model of State- and Stimulation-Dependent Modulation of Rhythmic Neural Activity and Connectivity. Front. Comput. Neurosci. 2020, 14, 575143. [Google Scholar] [CrossRef]
- Cabral, J.; Kringelbach, M.L.; Deco, G. Functional connectivity dynamically evolves on multiple time-scales over a static structural connectome: Models and mechanisms. Neuroimage 2017, 160, 84–96. [Google Scholar] [CrossRef]
- Venkadesh, S.; Van Horn, J.D. Integrative Models of Brain Structure and Dynamics: Concepts, Challenges, and Methods. Front. Neurosci. 2021, 15, 752332. [Google Scholar] [CrossRef] [PubMed]
- Deco, G.; Jirsa, V.K.; McIntosh, A.R. Emerging concepts for the dynamical organization of resting-state activity in the brain. Nat. Rev. Neurosci. 2011, 12, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.D.; Demirtas, M.; Anticevic, A. Biophysical Modeling of Large-Scale Brain Dynamics and Applications for Computational Psychiatry. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2018, 3, 777–787. [Google Scholar] [PubMed]
- Deco, G.; McIntosh, A.R.; Shen, K.; Hutchison, R.M.; Menon, R.S.; Everling, S.; Hagmann, P.; Jirsa, V.K. Identification of optimal structural connectivity using functional connectivity and neural modeling. J. Neurosci. 2014, 34, 7910–7916. [Google Scholar] [PubMed] [Green Version]
- Vasa, F.; Shanahan, M.; Hellyer, P.J.; Scott, G.; Cabral, J.; Leech, R. Effects of lesions on synchrony and metastability in cortical networks. Neuroimage 2015, 118, 456–467. [Google Scholar] [CrossRef] [Green Version]
- Kringelbach, M.L.; Cruzat, J.; Cabral, J.; Knudsen, G.M.; Carhart-Harris, R.; Whybrow, P.C.; Logothetis, N.K.; Deco, G. Dynamic coupling of whole-brain neuronal and neurotransmitter systems. Proc. Natl. Acad. Sci. USA 2020, 117, 9566–9576. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Cui, Y.; Zhang, Y.; Cao, H.; Zhou, G.; Shu, H.; Yao, D.; Xia, Y.; Chen, K.; Guo, D. Computational exploration of dynamic mechanisms of steady state visual evoked potentials at the whole brain level. Neuroimage 2021, 237, 118166. [Google Scholar] [CrossRef]
- Jafari, Z.; Kolb, B.E.; Mohajerani, M.H. Neural oscillations and brain stimulation in Alzheimer’s disease. Prog. Neurobiol. 2020, 194, 101878. [Google Scholar] [CrossRef]
- Pons, A.J.; Cantero, J.L.; Atienza, M.; Garcia-Ojalvo, J. Relating structural and functional anomalous connectivity in the aging brain via neural mass modeling. Neuroimage 2010, 52, 848–861. [Google Scholar] [CrossRef]
- Filippi, M.; Agosta, F. Structural and functional network connectivity breakdown in Alzheimer’s disease studied with magnetic resonance imaging techniques. J. Alzheimers Dis. 2011, 24, 455–474. [Google Scholar] [PubMed]
- Dickerson, B.C.; Salat, D.H.; Bates, J.F.; Atiya, M.; Killiany, R.J.; Greve, D.N.; Dale, A.M.; Stern, C.E.; Blacker, D.; Albert, M.S.; et al. Medial temporal lobe function and structure in mild cognitive impairment. Ann. Neurol. 2004, 56, 27–35. [Google Scholar] [PubMed] [Green Version]
- Sanchez-Rodriguez, L.M.; Iturria-Medina, Y.; Baines, E.A.; Mallo, S.C.; Dousty, M.; Sotero, R.C. Design of optimal nonlinear network controllers for Alzheimer’s disease. PLoS Comput. Biol. 2018, 14, e1006136. [Google Scholar]
- Francis, P.T.; Ramirez, M.J.; Lai, M.K. Neurochemical basis for symptomatic treatment of Alzheimer’s disease. Neuropharmacology 2010, 59, 221–229. [Google Scholar] [PubMed]
- Khan, A.F.; Adewale, Q.; Baumeister, T.R.; Carbonell, F.; Zilles, K.; Palomero-Gallagher, N.; Iturria-Medina, Y. Personalized brain models identify neurotransmitter receptor changes in Alzheimer’s disease. Brain 2022, 145, 1785–1804. [Google Scholar] [CrossRef] [PubMed]
- Coito, A.; Michel, C.M.; Vulliemoz, S.; Plomp, G. Directed functional connections underlying spontaneous brain activity. Hum. Brain Mapp. 2019, 40, 879–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, J.; Guo, H.; Gao, Y.; Wang, X.; Cui, H.; Chen, Z.; Wang, B.; Xiang, J. Altered Directed Functional Connectivity of the Hippocampus in Mild Cognitive Impairment and Alzheimer’s Disease: A Resting-State fMRI Study. Front. Aging Neurosci. 2019, 11, 326. [Google Scholar] [PubMed] [Green Version]
- Khazaee, A.; Ebrahimzadeh, A.; Babajani-Feremi, A. Classification of patients with MCI and AD from healthy controls using directed graph measures of resting-state fMRI. Behav. Brain Res. 2017, 322, 339–350. [Google Scholar] [CrossRef] [Green Version]
- Tijms, B.M.; Ten, K.M.; Gouw, A.A.; Borta, A.; Verfaillie, S.; Teunissen, C.E.; Scheltens, P.; Barkhof, F.; van der Flier, W.M. Gray matter networks and clinical progression in subjects with predementia Alzheimer’s disease. Neurobiol. Aging 2018, 61, 75–81. [Google Scholar] [CrossRef]
- Xu, D.; Cheng, W.; Luo, D.; Gu, Y.; Liu, X.; Ni, J.; Zong, B.; Chen, H.; Zhang, X. Adaptive Neural Network for Node Classification in Dynamic Networks. In Proceedings of the 2019 IEEE International Conference on Data Mining (ICDM), Beijing, China, 8–11 November 2019. [Google Scholar]
- Kundu, S.; Lukemire, J.; Wang, Y.; Guo, Y. A Novel Joint Brain Network Analysis Using Longitudinal Alzheimer’s Disease Data. Sci. Rep. 2019, 9, 19589. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| References | Dynamics Model | Subject | Imaging Modality | Means of Simulating AD | Main Conclusion |
|---|---|---|---|---|---|
| (de Haan et al. 2012) [37] | Neural Mass Model | NC\MCI\AD | EEG | Lowering synaptic strength | Excessive neuronal activity and hub vulnerability. |
| (Ghorbanian et al. 2014) [38] | Coupled Duffing–van der Pol Oscillator Model | NC\AD | EEG | Simulation based on empirical data | α dominant for CTL subject and θ dominant for AD patients. |
| (de Haan et al. 2017) [39] | Neural Mass Model | NC\AD | EEG | Changing neuronal excitability levels due to varied threshold potential (Vd) settings | AD-like network degeneration can be countered by global stimulation of excitatory neurons. |
| (Demirtas et al. 2017) [35] | Hopf Normal Model | NC\PAD\MCI\AD | fMRI | Manipulating the bifurcation parameter | Synchronization ↓, FC strength ↓, and the significant EC differences in AD were located in left temporal lobe. |
| (Zimmermann et al. 2018) [40] | Reduced Wong–Wang Model | NC\MCI\AD | fMRI | Simulation based on empirical data | The model parameters correlated with cognition and the predictive capacity ↑. |
| (Alderson et al. 2018) [31] | Kuramoto Model | NC\MCI\AD | fMRI | Lesioned the structural connections of healthy subjects | Metastability ↓; damage was centered around highly connected nodes; abnormal network topology; a link between metastable neural dynamics, cognition, and the structural integrity of the human brain. |
| (Stefanovski et al. 2019) [34] | Jansen–Rit Model | NC\MCI\AD | fMRI | Based on individual PET-derived distributions of Abeta | Spatial heterogeneous Abeta distribution, impaired inhibitory function, and neural frequencies ↓. |
| (Ruiz-Gomez et al. 2019) [33] | Kuramoto Model | NC\MCI\AD | EEG | Simulation based on empirical data | Θ band ↑ and α band ↓. |
| (Cakir 2020) [41] | Izhikevich Model and Neural Mass Model | NC\AD | fMRI | Simulation based on empirical data | The alpha rhythms ↓ in the thalamic, and fALFF of slow-4 band ↓ in the striatum. |
| (Li et al. 2020) [32] | Thalamo-cortico-thalamic (TCT) Circuitry Model | Simulated AD | EEG | Decreasing synaptic connectivity parameters | Synapse loss and alpha rhythm ↓. |
| (Bachmann et al. 2020) [6] | Leaky Integrate-and-Fire (LIF) Model | Simulated AD | EEG | Adjusting the weight of excitatory synapses | The loss of excitatory synapses on excitatory neurons. |
| (Arbabyazd et al. 2021) [42] | Stochastic Linear Model (SLM) and Mean-Field Model (MFM) | NC\MCI\AD | fMRI | Simulation based on empirical data | Realistic data can be generated by whole-brain modeling. |
| (Triebkorn et al. 2022) [43] | Jansen–Rit Neural Mass Model | NC\MCI\AD | fMRI | Adjusting the global scaling factor G and linked local Aβ concentrations | Local hyperexcitation caused by Aβ can classify AD. |
| (van Nifterick et al. 2022) [44] | Neural Mass Model | SCD\MCI\AD | MEG | Adjusting single relevant parameters | Oscillatory ↓; hyperexcitation. |
| (Das and Puthankattil 2022) [36] | Kuramoto Model | NC\MCI-AD | EEG | Edges originating from one specific region of the cortex are set to the lowest value | Functional connectivity ↓ and complexity ↓ in anterior and central regions. |
| (Patow et al. 2023) [45] | Balanced Excitation–Inhibition Model | NC\MCI\AD | fMRI | Adjusting the inhibitory bias and scaling parameters | The neuronal activity of Aβ over tau in MCI, while tau dominates over Aβ in AD. |
| (Salimi-Nezhad et al. 2023) [46] | Pinsky–Rinzel Neuron Model | Healthy rats | LFP | Eliminated 75% of MS cholinergic neurons | Selectively stimulating the remaining healthy cholinergic neurons was sufficient for network recovery. |
| (Sanz et al. 2023) [47] | Hopf Normal Model | NC\AD\bvFTD | fMRI | Simulation based on empirical data | Key nodes to transition from AD towards the healthy state included the hippocampus as well as temporo-posterior regions. |
| (Alexandersen et al. 2023) [48] | Neural Mass Model | NC | E/MEG | Adjusting the Aβ and τP damage variables | Excitatory neuronal activity↓; oscillatory↓ independently of structural changes due to axonal damage. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Lu, J.; Li, D.; Xiang, J.; Yan, T.; Sun, J.; Wang, B. Alzheimer’s Disease: Insights from Large-Scale Brain Dynamics Models. Brain Sci. 2023, 13, 1133. https://doi.org/10.3390/brainsci13081133
Yang L, Lu J, Li D, Xiang J, Yan T, Sun J, Wang B. Alzheimer’s Disease: Insights from Large-Scale Brain Dynamics Models. Brain Sciences. 2023; 13(8):1133. https://doi.org/10.3390/brainsci13081133
Chicago/Turabian StyleYang, Lan, Jiayu Lu, Dandan Li, Jie Xiang, Ting Yan, Jie Sun, and Bin Wang. 2023. "Alzheimer’s Disease: Insights from Large-Scale Brain Dynamics Models" Brain Sciences 13, no. 8: 1133. https://doi.org/10.3390/brainsci13081133
APA StyleYang, L., Lu, J., Li, D., Xiang, J., Yan, T., Sun, J., & Wang, B. (2023). Alzheimer’s Disease: Insights from Large-Scale Brain Dynamics Models. Brain Sciences, 13(8), 1133. https://doi.org/10.3390/brainsci13081133