1. Introduction
Sensory input and movement performance are intimately linked for both goal-directed reaching movements and multi-limb coordination tasks [
1,
2,
3,
4]. In the natural environment, both visual and proprioceptive feedback are available to plan and update goal-directed reaching movements [
5]. The multiple process model of limb control proposes that goal-directed aiming movements are comprised of two components, but multiple processes within these two components [
5,
6]. According to this model, the first component of the movement is defined by impulse control, which includes the pre-planned part of the movement where the limb covers most of the distance to reach the vicinity of the target. The second component is the limb-target control phase where the available sensory inputs are compared to internal representations. The multiple process model considers multiple factors, such as noise in the neural-motor system, force-related error, as well as efficiency of energy expenditure. It is suggested that the two components of the multiple process model are not independent, and that visual feedback is the main sensory driver of the impulse control phase. Limited visual input leads to decreased accuracy and precision. The availability of visual feedback results in the correction of motor planning errors during the latter phase of the movement trajectory [
7].
Proprioception in combination with visual inputs is another major source of information for limb-target regulation. Thus, effective limb control includes integration of expected and actual sensory inputs from multiple sensory sources. There is extensive research on the role of vision in controlling goal-directed movement of the upper limb due to the rich spatial information that vision can provide during movements [
8,
9,
10,
11,
12,
13]. Somatosensory information has a complementary role in the control of human movement by providing information about the location of the limb in the space as well as the relative location of limbs to each other [
1,
2,
3,
4,
7]. Thus, it is important to know how these two major sources of information are integrated for voluntary limb control.
Somatosensory input includes sensory input from the body and can be considered as a combination of both tactile and proprioceptive sensory feedback [
14,
15]. Tactile and proprioceptive inputs are integral for performing everyday tasks, such as grooming, dressing, and meal preparation. Independence may be reduced or lost if the ability to perform functional tasks is altered. For example, if individuals have decreased somatosensory input, they have difficulty reaching for or manipulating an object without having to look at their limb. Indeed, individuals with a variety of sensorimotor disorders that alter the central and/or peripheral nervous system (e.g., stroke, multiple sclerosis, Parkinson’s disease, Diabetes) experience reduced or disrupted somatosensory input. These individuals must then learn or re-learn voluntary goal-directed actions in the presence of altered somatosensory input [
16,
17].
Knowledge about the individual and integrative role of different sensory inputs is crucial for rehabilitation programs when either of these sensory inputs are deficient. Therefore, given the importance of somatosensory input for motor control, and the consequences of its loss, researchers have used a variety of methods to disrupt somatosensory feedback in a neurotypical population to better understand the contributions of somatosensory input for movement control. For example, the natural visual and proprioceptive relationship can be altered by rotating augmented visual feedback [
18,
19]. Proprioception can be targeted directly using muscle tendon vibration [
20,
21,
22]. While these methods have contributed to our understanding of the role of somatosensory feedback in motor control, the more common sensation of paresthesia experienced by individuals with sensorimotor disorders, and the impact on unconstrained goal-directed movements used in functional tasks, is not well understood [
23].
Another method to alter somatosensory feedback is to induce paresthesia using transcutaneous nerve stimulation to impair somatosensory inputs [
23,
24,
25]. Paresthesia can be induced by transcutaneous electrical stimulation applied over the estimated path of the chosen peripheral nerve [
26,
27]. This method does not eliminate the tactile and proprioception inputs, rather it causes a condition that resembles a radiating paresthesia pathology, or a feeling of tingling and numbness radiating along the course of the targeted peripheral nerve. The induced radiating paresthesia resembles the loss of sensory input in a neurologic injury or disease, which makes it a relevant and realistic model.
When performing a variety of motor tasks, including balance and goal-directed reaching tasks, multiple sensory modalities provide input about where and how our limbs are moving. It has been demonstrated that when the preferred modality becomes degraded then the appropriate or preferred modality will shift [
8,
9,
10]. This shift in the preferred sensory modality is consistent with what happens in neurologic injuries or diseases that affect sensory systems. For example, humans become more dependent on vision to maintain balance if the sensitivity of their somatosensory and/or vestibular systems decreases [
28,
29].
However, less is known about how humans compensate for altered or reduced somatosensory input during functional goal-directed movements. Elliott et al.’s multiple process model of goal-directed movement [
5] incorporates sensory input from vision and somatosensation. However, the systematic investigation of altered somatosensation, as observed in cases of peripheral nerve damage caused by disease or injury, has been lacking. Given that humans commonly experience changes in afferent inputs, gaining an understanding of the integration between vision, somatosensation, and altered somatosensory inputs is crucial. This knowledge will provide valuable insights into how individuals can adapt their movement performance strategies and the effects of modified somatosensory input.
The specific objectives of the current study were: (1) to determine, in an otherwise healthy nervous system, if the presence of induced paresthesia affects the speed and accuracy of goal-directed movements; (2) to determine if disrupted somatosensory feedback in the effector (index finger and hand) leads to an increased reliance on visual feedback of the target specifically. We hypothesized that individuals will be more reliant on visual feedback in the presence of induced paresthesia, and both the speed and accuracy of goal-directed reaching movements would be affected, and to a greater extent when vision of the target was unavailable [
30]. The premise for this manipulation is that when somatosensory feedback is disrupted then, as predicted by the multiple process model, the need for online visual feedback to be available to compare the location of the hand relative to the target location will increase. An additional objective was to explore how participants’ movement strategies may change with experience performing the aiming task in the presence of paresthesia. To carry this out, trials from early and late performance were compared to understand how individuals may adjust their movement control strategies in the context of disrupted somatosensory feedback. Specifically, we hypothesized that participants would update their movement control strategies to include more pre-planned movements that rely less on the need to make online corrections.
3. Results
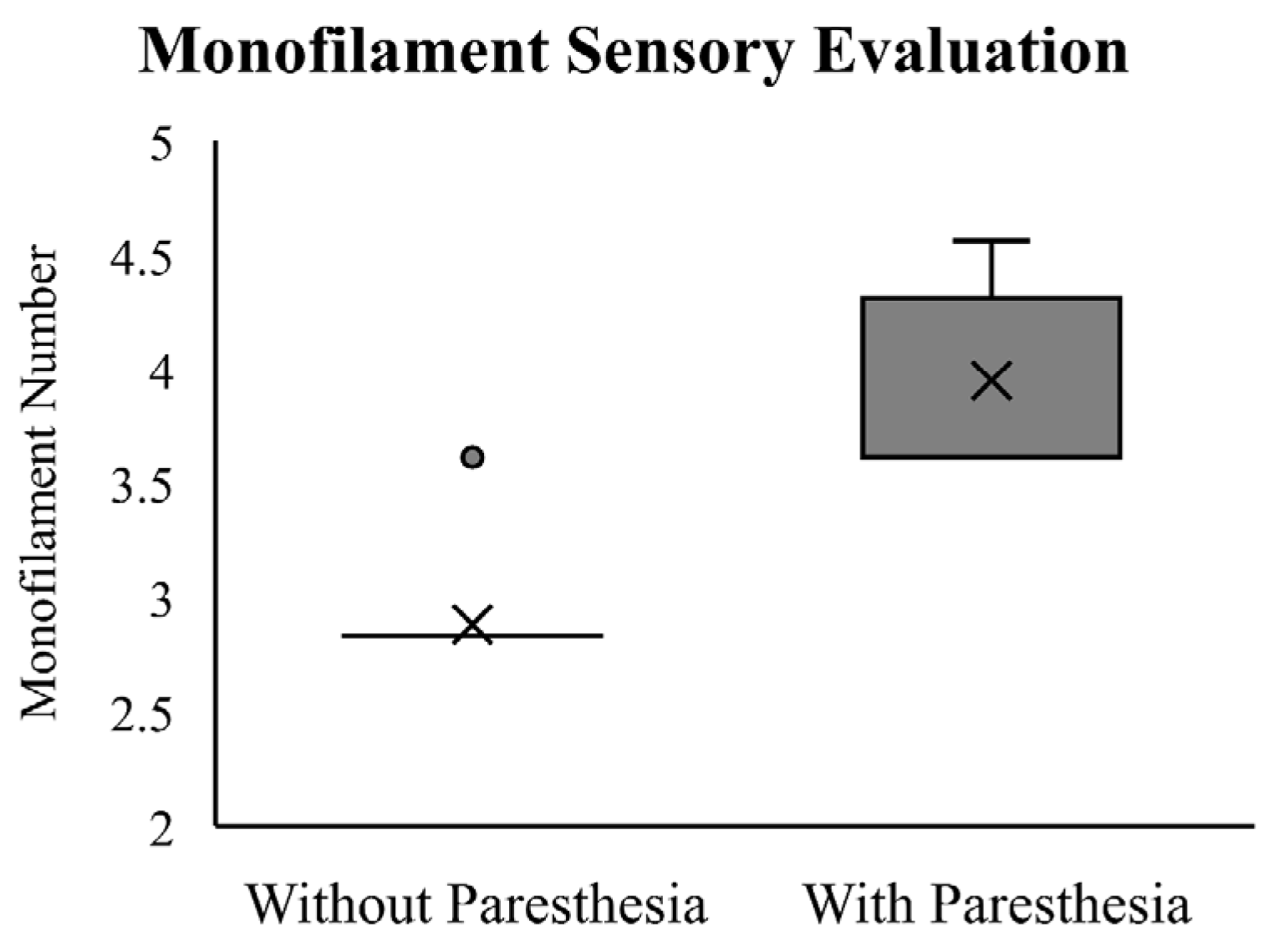
The Wilcoxon Signed-Rank Test showed that participants required a significantly thicker monofilament to detect skin deformation changes after paresthesia was induced (Z = −3.60,
p < 0.01). The results of the monofilament testing at baseline and with induced paresthesia are reported in
Figure 3. At baseline, all but one of the participants sensed the monofilament size of 2.83, which falls within the normal range based on Touch-Test
® thresholds. One participant detected the monofilament size of 3.61, suggesting a decrease in light touch sensitivity. Critical to the experimental manipulation, following induced paresthesia, all participants reported a larger monofilament size, ranging from 3.61 to 4.56 (median = 3.61).
Motion capture data for two participants were excluded from the statistical analysis because displacement data for more than 50% of the trials were missing due to IREDs becoming obscured during the reaching movement. Therefore, data analysis is based on the remaining 14 participants.
3.1. Temporal Measurments
For reaction time, no significant main effect or interaction was found for the factors of target vision, time, and paresthesia (results included in
Table 1).
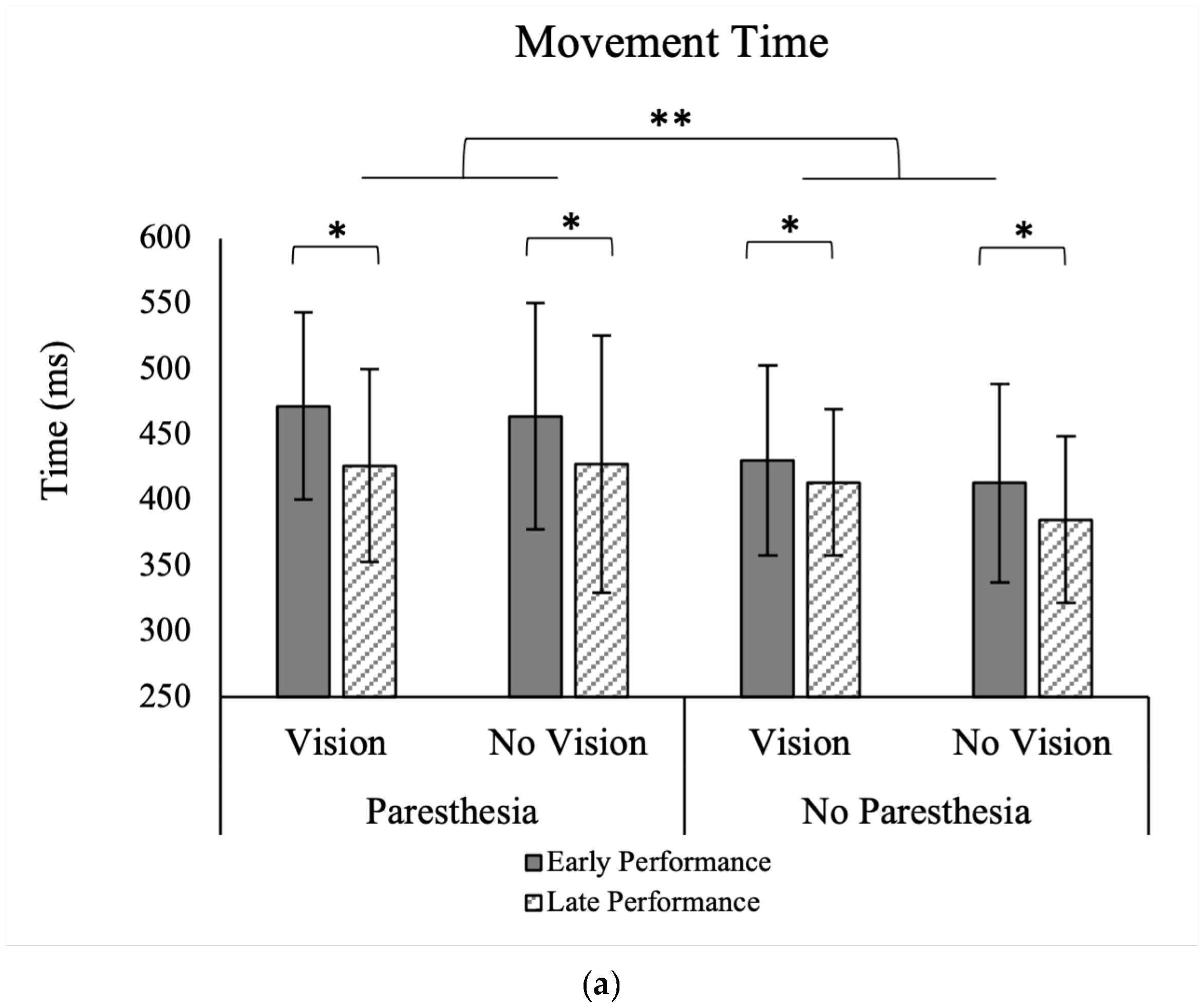
For MT, there were significant main effects for time, F (1, 13) = 15.48,
p = 0.002, ηp
2 = 0.54, and paresthesia, F (1, 13) = 8.50,
p = 0.012, ηp
2 = 0.39 (
Figure 4a). Movement time was shorter without paresthesia than with paresthesia, and during the late 20 trials than the early 20 trials (
Table 1). There were no significant interactions between the factors of target vision, time, and paresthesia.
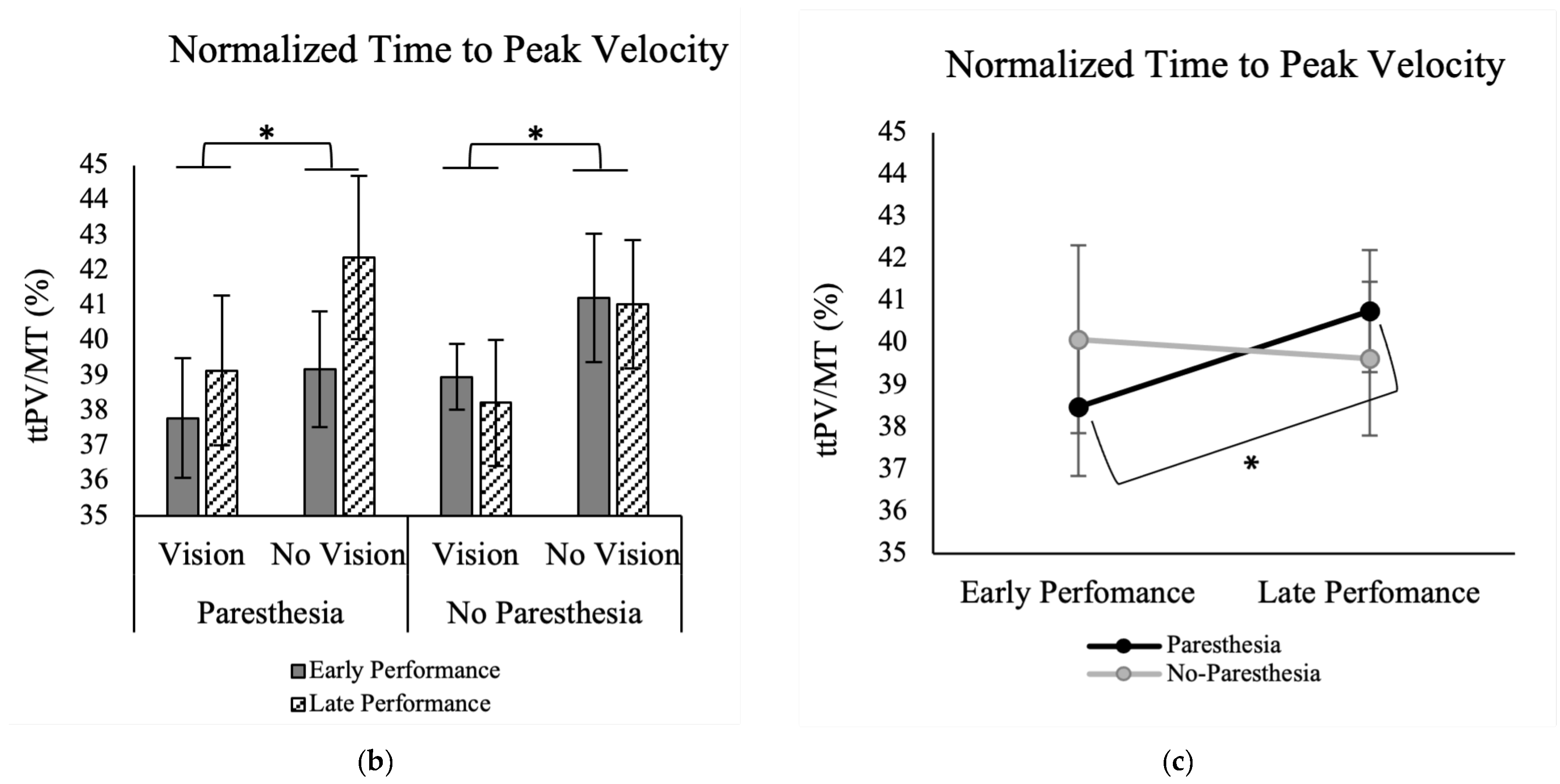
Results of normalized ttPV showed a significant main effect for the factor of Target Vision, F (1, 13) = 11.59,
p = 0.005, ηp
2 = 0.47. As shown in
Figure 4b, normalized ttPV was longer in without target vision (M = 40.9; SD = 7.1) than full vision conditions (M = 38.5; SD = 6.2). Also, there was a significant interaction between the factors of time and paresthesia, F (1, 13) = 7.25,
p = 0.018, ηp
2 = 0.36 (
Figure 4c). As illustrated in
Figure 4c, Tukey’s HSD test showed that while normalized ttPV for the early and late practice trials was not significantly different, when paresthesia was present, participants had longer ttPV for late trials (M = 40.2; SD = 7.6) versus the early trials (M = 39.3; SD = 5.8) (
Figure 4c).
3.2. Spatial Measurments
In the primary axis of movement, no significant main effects or interactions were found for the factors of target vision, time, or paresthesia for CE and VE (
Table 1).
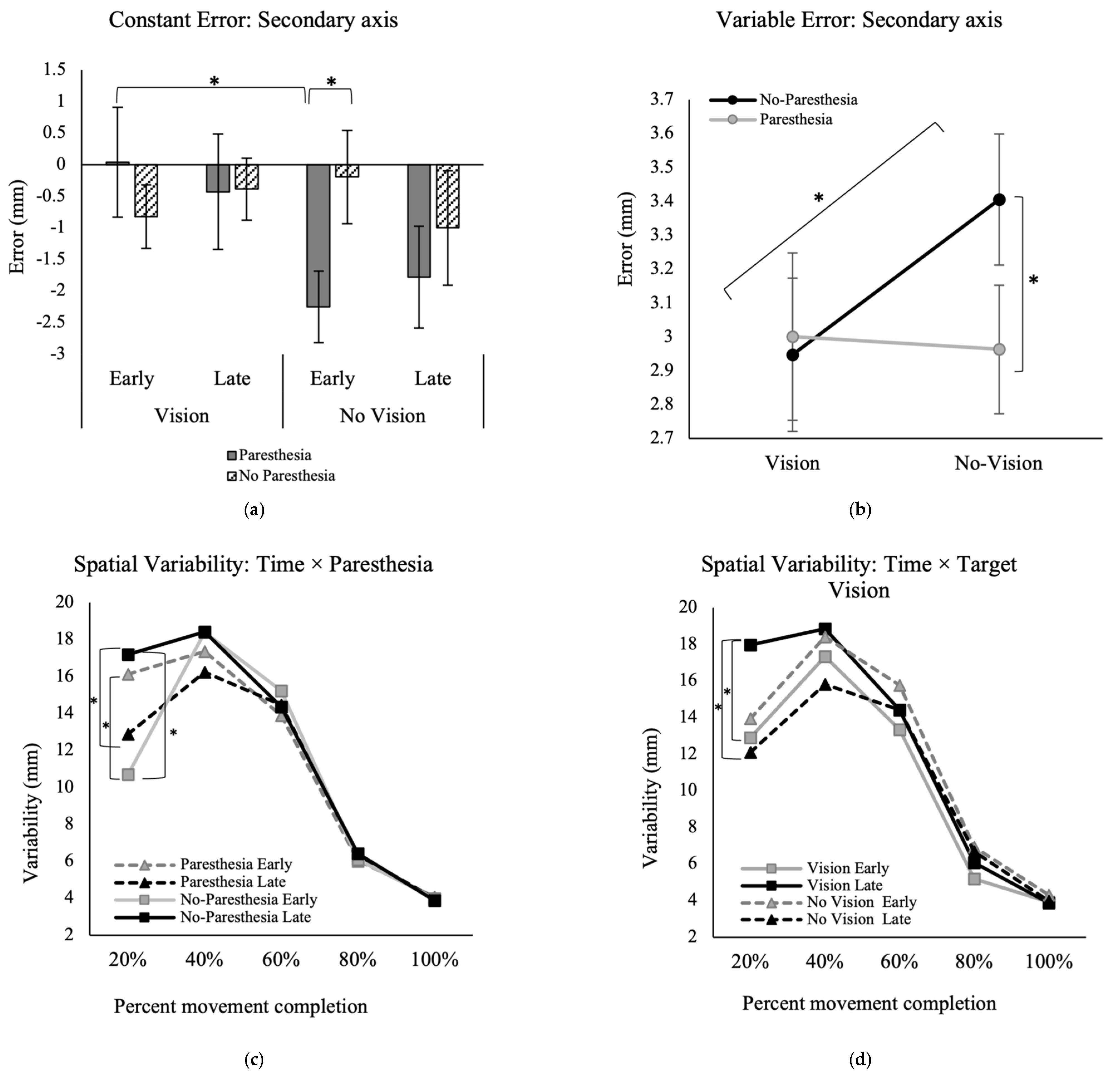
In the secondary movement axis, however, there was a significant interaction between the factors of time, paresthesia, and target vision for the outcome of CE, F (1, 13) = 6.03,
p = 0.029, ηp
2 = 0.32 (
Figure 5a). Post hoc analysis showed that when vision of the target was removed, the CE in early performance was significantly higher when paresthesia was induced (M = −2.2; SD = 2.1) versus without paresthesia (M = −0.2; SD = 2.8).
For VE in the secondary movement axis, there was a significant interaction between the factors of paresthesia and target vision, F (1, 13) = 6.48,
p = 0.02, ηp
2 = 0.33 (
Figure 5b). Post hoc analysis showed that in the no-paresthesia conditions, participants had significantly higher variability when vision of the target was removed (M = 3.4; SD = 0.9) versus when vision was available (M = 2.9; SD = 0.8). Also, when target vision was not available, participants had significantly higher variability without paresthesia (M = 3.4; SD = 0.9) compared to with paresthesia (M = 3.0; SD = 0.7).
3.3. Movement Trajectories
The repeated measures ANOVA for the variability of the movement trajectory in the primary movement axis showed a significant main effect for percent movement completion, F (4, 52) = 87.2,
p < 0.001, ηp
2 = 0.87. Also, there was a significant three-way interaction between factors of percent movement completion, time, and paresthesia, F (2.1, 27.3) = 10.74,
p < 0.001, ηp
2 = 0.45 (
Figure 5c), and another interaction between the factors of time and target vision, F (1, 13) = 13.25,
p = 0.003, ηp
2 = 0.50 (
Figure 5d). Post hoc analysis for the interaction of time, percent movement completion, and paresthesia showed that during early practice trials when paresthesia was induced, variability was significantly higher compared to the no-paresthesia condition at 20% of the movement; the mean difference of early practice with paresthesia versus no-paresthesia was 5.4 mm. However, this difference was reversed during late trials. Specifically, following more practice without paresthesia, late trials showed significantly higher variability at 20% movement time compared to the paresthesia condition; the mean difference of VE at 20% of the movement for the paresthesia versus no-paresthesia condition for late practice trials was 4.3 mm (
Figure 5c). Comparison of the late versus early trials when no paresthesia was induced also showed more variability at late performance (mean difference = 6.5 mm;
Figure 5c). Post hoc analysis for the interaction of target vision and time showed that when target vision was available, participants had significantly higher variability in the first 20% of movement time following more practice (late trials) versus early practice trials (
Figure 5d). Also, in late practice, the VE at 20% MT was significantly higher with target vision versus without target vision.
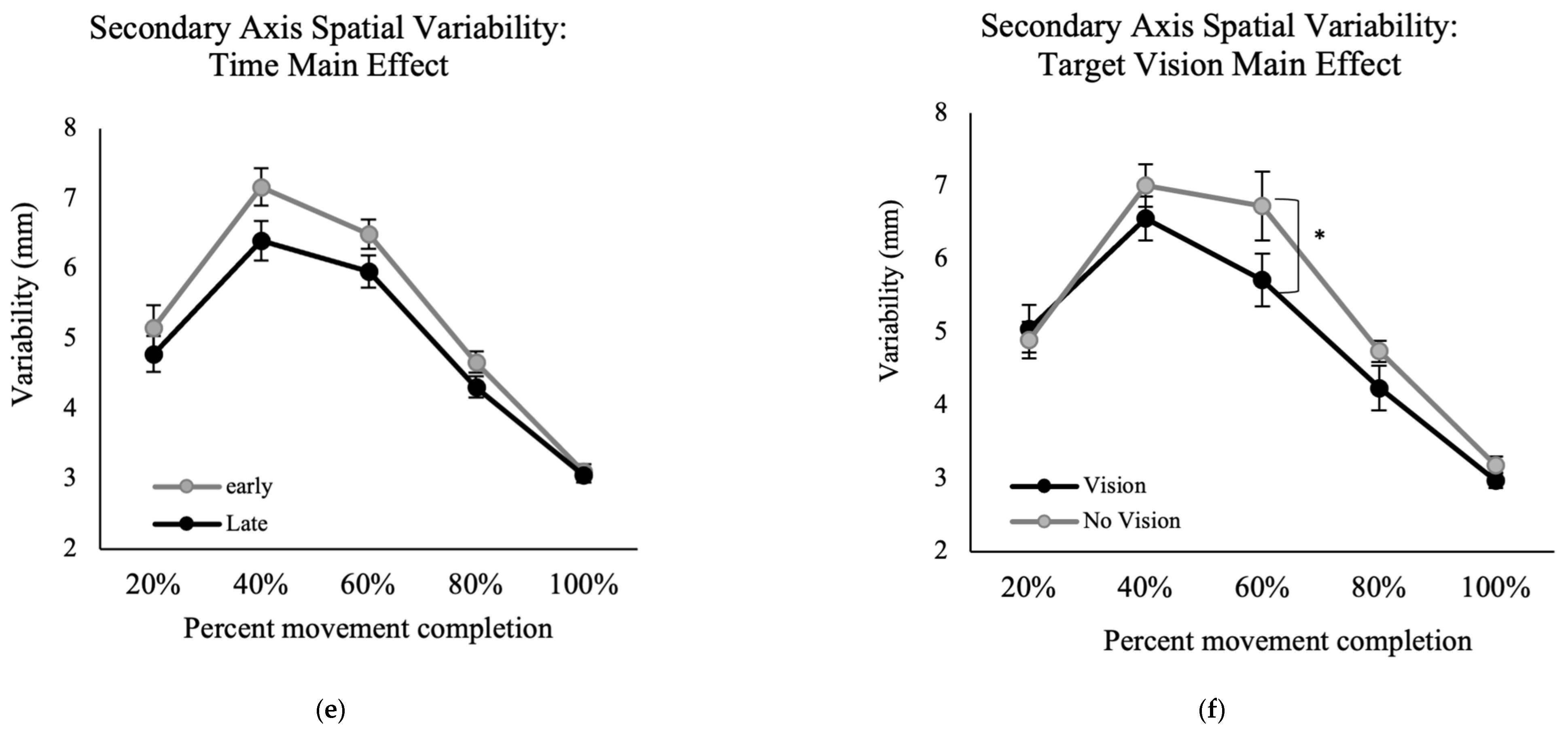
Results of the VE for movement trajectory in the secondary axis of movement showed a significant main effect for the factors of time, F (1, 13) = 9.22,
p = 0.01, ηp
2 = 0.41 (
Figure 5e) and percent movement time, F (2.0, 25.7) = 108.00,
p < 0.001, ηp
2 = 0.89 (
Figure 5f), indicating higher VE at late versus early performance. Also, there was a significant interaction between percent movement time and target vision, F (4, 52) = 3.01,
p = 0.026, ηp
2 = 0.19. Post hoc analysis for this interaction showed that at 60% of movement time, the with target vision conditions had significantly greater variability than without target vision conditions (
Figure 5e).
4. Discussion
In the current study, we explored if and how participants update their movement control strategies in the presence of induced paresthesia. Participants performed a functional goal-directed aiming task, with or without vision of the target, while paresthesia was induced in order to understand how motor control strategies change when trying to move quickly and accurately while experiencing paresthesia. Neurotypical individuals were recruited to participate in the current experiment and paresthesia was induced using constant current stimulation of the median nerve to assess the contribution of somatosensory input from the hand specifically.
We hypothesized that both speed and accuracy of goal-directed reaching movements would be affected by paresthesia, and to a greater extent when vision of the target was unavailable for limb-target control mechanisms [
8,
9,
10]. As expected, there was no significant difference between the conditions with and without induced paresthesia when vision of the target was available. However, in the absence of visual feedback, the presence of induced paresthesia had a greater impact on performance compared to the condition without paresthesia. It is well known that limb-target control heavily relies on visual feedback. Therefore, when both somatosensory input was interrupted and vision of the target was removed, the insufficient sensory inputs hindered the ability to make accurate limb-target corrections.
4.1. Effect of Sensory Manipulation on Endpoint Accuracy
We predicted that paresthesia would affect movements more robustly when vision of the target was not available. There were no differences in endpoint accuracy in the primary axis of movement. Only the accuracy measures in the secondary movement axis (i.e., mediolateral) were consistent with this prediction. In early performance trials, CE in the secondary axis became significantly larger with paresthesia compared to the no-paresthesia condition, but only when vision of the target was removed. The presence of paresthesia did not have an effect on endpoint accuracy when vison of the target was available.
Constant error early in performance, without target vision and with paresthesia, was significantly larger than early performance in the paresthesia condition with full vision (
Figure 5a). This pattern indicates that visual feedback was the dominant source of feedback for endpoint accuracy as predicted by the multiple process model. Paresthesia without target vision led to participants being consistently biased towards flexion, inferred from the consistent undershoot errors (
Figure 5a). One explanation for this bias could be the imbalance in the proprioception caused by stimulation of the median nerve only. Stimulation of the median nerve only could lead to a false sense of location and tendency for overcorrection towards the midline when proprioception was distorted [
41]. In a study by Goodman et al. [
21], simultaneous muscle tendon vibration of elbow flexors and extensors was used to manipulate somatosensory inputs in neurotypical adults [
21]. These authors applied muscle tendon vibration before trials of a horizontal goal-directed aiming task, while vision of the whole environment was manipulated using occlusion goggles. They found that vibration application resulted in increased CE in a goal-directed reaching task when vision was also removed. Their results for CE in the primary axis showed target undershoots, similar to the finding of the current study in the secondary axis. They also found target overshoots in the anterior–posterior axis (secondary axis in that study), similar to findings of the current study for CE in the anterior–posterior axis (which was the primary axis in the current study). It should be noted that the current study was performed in the anteroposterior direction, while the aiming task Goodman et al. used was performed mostly in the mediolateral direction. Although these two studies used different methods for disrupting proprioception, there was a tendency to perceive the position of the upper limb as closer to the midline. Given the location of the stimulations used in these two studies for interrupting proprioception, and the possible tendency to position the upper limb closer to the midline, the consistent findings reinforce the result that interrupting proprioception leads to target undershoots.
Endpoint variability (VE) in the secondary axis was also affected with paresthesia only when vision of the target was removed (
Figure 5b). However, in contrast to our expectation, in the no-paresthesia condition specifically, VE was significantly higher when vision of the target was removed compared to when vision of the target was available, namely, induced paresthesia did not increase endpoint variability in the secondary axis.
4.2. Effect of Sensory Manipulation on Movement Strategy
4.2.1. Time to Peak Velocity and Change in Movement Strategy
We found that MT was shorter after performing more trials, which aligns with previous findings [
32]. Movement time was also shorter when performing the task without induced paresthesia compared to the induced paresthesia condition. No significant interaction was found for the factors of target vision and paresthesia. Thus, overall MT was impacted by induced paresthesia and improved with practice in all four conditions.
Normalized ttPV was the temporal variable that supported our hypothesis, which predicted that individuals will be more reliant on visual feedback in the presence of a somatosensory disruption. In the current study, when vision of the target was available, regardless of paresthesia condition, ttPV was shorter (see
Figure 4b). The changes in ttPV can be interpreted using Elliott et al.’s multiple process model of limb control [
5,
42]. In this model, there are early and late online control processes: the impulse control and limb-target regulation phases. Most of the impulse control phase happens before the limb reaches peak velocity and is the distance covering a portion of the movement that determines the direction and velocity of the movement. Limb-target regulation happens after the peak velocity is reached and is performed by using the visual and somatosensory inputs to fine tune the landing of the limb on the target. By considering the ttPV finding in the current study, we can interpret that when vision of the target was available, participants reached the proximity of the target quickly and spent more time in limb-target phase, relying on the available visual information of the target location and the limb, whether or not somatosensory input was interrupted [
5,
40]. However, when vision of the target was removed, the time spent before PV (ttPV) was a larger portion of the movement time. Although statistical analysis did not show a significant effect of paresthesia on the ttPV in the without target vision conditions, analysis of the late versus early performance trials showed that when paresthesia was present, as more trials were performed (comparing first versus last twenty trials), the normalized ttPV increased, namely, a larger percentage of the movement time was spent in the impulse control phase, and less in limb-target regulation. One explanation for this finding could be that participants chose a different strategy for controlling their movement by pre-planning their movements. Pre-planning would be advantageous because they did not have somatosensory input or target vision available for feedback and current control. In other words, with practice, they adopted a strategy that included spending less time on limb-target regulation [
32,
43].
The present findings are consistent with the study conducted by Goodman et al. (mentioned above) [
21]. Goodman et al. also found that when vision was removed, or when vibration was applied to interrupt the somatosensory input, participants spent less time after peak velocity for online corrections to the limb movement.
4.2.2. Spatial Variability and Change in Movement Strategy
According to the findings of the variability of movement trajectories, participants appear to have selected different strategies when performing the task with and without paresthesia. Variability of the movement trajectory early in the movement (~50% of the movement) is known to be associated with motor planning and offline control [
38], while spatial variability late in the movement is thought to reflect online corrective processes. Looking at the results of the spatial variability in the primary axis, regardless of the available visual input, when there was no paresthesia, participants were more variable at movement initiation in late trials compared to both early trials without paresthesia and late trials with paresthesia (
Figure 5c). Movement initiation was also more variable with paresthesia than without paresthesia for early trials specifically. We propose that when there was no paresthesia, and regardless of the availability of the target, with more practice participants learned to use more open-loop control strategies, including pre-planned initial impulses at movement initiation. This strategy of more forceful initial impulses is expected to lead to higher impulse variability as detected by a larger position variability at 20% of movement time [
44].
Although participants were asked to move as fast as they could, the average movement time for the task was long enough for participants to perform limb-target corrections and reduce endpoint error and variability (i.e., overall mean of MT = 435 ms is longer than 200 ms, “see Schmidt, 1979 for more information”). On the other hand, when paresthesia was induced, participants appeared to use a more conservative movement strategy after more practice with the task. Specifically, we found lower VE at 20% of movement time during late trials with paresthesia versus early trials (
Figure 5c). One explanation for this finding could be that paresthesia may have increased neural-motor noise and affected force specification processes. Thus, participants chose a safe strategy with smaller impulses at movement initiation, leading to less variability at 20% of movement time. The movement time findings are also consistent with this explanation since movements became shorter with practice and longer with paresthesia.
The results of the movement trajectories in the mediolateral axis showed that with more practice, or when vision of the target was available, movements became less variable around 40% and 60% of movement time (
Figure 5e,f). This reduced variability of the movement indicates a more pre-planned movement and fewer online corrections when vision of the target was not available.
4.3. Effects of Practice
A secondary objective of the current study was to investigate how ongoing practice with the changed sensory inputs would affect movement strategy and performance. As expected, regardless of experimental condition, participants had shorter MTs with more practice (
Figure 4a). Also, normalized ttPV in the paresthesia condition became longer with more practice. These findings indicate that when paresthesia was induced, participants learned to spend more time during the pre-planned impulse control phase, and subsequently fewer online corrections. Although not statistically significant, the trends in movement strategies seemed to be larger when vision of the target was not available (
Figure 4b). In summary, the analysis of early versus late trials demonstrated that at least some of the changes in the temporal and spatial movement characteristics that resulted from manipulation of the sensory input were alleviated with practice.
4.4. Target Vision Availability and Movement Strategy
In the current study, visual input was manipulated by obscuring the vision of the target only because vison of the target is necessary for limb-target control processes that are used to acquire the target accurately [
45,
46,
47]. Lack of target vision (or its memory) is expected to lead performers to use more pre-planned movements as well as using kinesthetic or feedforward sources of information [
47]. In the current study, vision of the target was removed upon movement initiation; therefore, the memory of the target location was not decayed and was available for memory guided limb-target regulation [
46]. Hence, since target vision was removed in the current study, the effect of paresthesia on execution of a pre-planned and memory guided aiming task was assessed. As expected, the results of this study showed that participants’ aiming accuracy was significantly different in the two without target vision conditions with and without paresthesia (CE in the secondary axis,
Figure 5a). A possible mechanism considered for this finding is that the memory-guided movement required more mental effort for the participants and adding induced paresthesia could have overloaded attentional resources, which would interfere with visual attention towards dynamic limb location and online limb-target control processes [
48]. It is possible that although vision of the limb was present, and the target location memory should have been available, that participants did not engage online limb-target control processes and used a safe strategy where they pre-planned their movements. The findings of movement trajectories in the secondary axis also support a pre-planned movement strategy when target vision was removed. Removing target vision increased movement spatial variability at 60% of the movement time. Another explanation for the altered movement control strategy when target vision was removed could be that with memory guided movement control participants relied on their perception of the target location [
46], which itself was likely distorted or biased as a result of inducing paresthesia by only stimulating the median nerve.
4.5. Limitations and Future Directions
One limitation of the current study was the choice of the primary axis of the movement. As observed in the results, median nerve stimulation caused endpoint bias in the mediolateral direction. A task with a mediolateral direction as the primary movement would likely be more sensitive to changes in motor performance as a result of induced paresthesia caused by median nerve stimulation. Similarly, additional insights into the effects of the sensory perturbation on motor control could be assessed with the use of neurophysiological techniques such as transcranial magnetic stimulation (TMS). In the current study, monofilament pressure testing was utilized to assess gross sensory pressure changes as a result of induced paresthesia. However, incorporating a perceptual evaluation of illusory limb location induced by paresthesia, coupled with a measurement of sensory acuity, would have provided a more comprehensive understanding that included both subjective and objective dimensions of the effects of induced paresthesia. This assessment would facilitate a more refined interpretation of the movement accuracy outcomes and a more comprehensive evaluation of the intensity of the induced paresthesia. Future studies will also benefit from extending the current research to encompass neurodiverse populations, such as individuals experiencing typical aging, Down syndrome, or autistic traits.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}