

Food Finding Test without Deprivation: A Sensorial Paradigm Sensitive to Sex, Genotype, and Isolation Shows Signatures of Derangements in Old Mice with Alzheimer’s Disease Pathology and Normal Aging



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Social Conditions
2.3. Physical Status and Behavioral Assessment
2.3.1. Physical Status
2.3.2. The Food Finding Test of Olfactory Ability without Food Deprivation
2.4. Statistical Analysis
3. Results
3.1. Physical Status
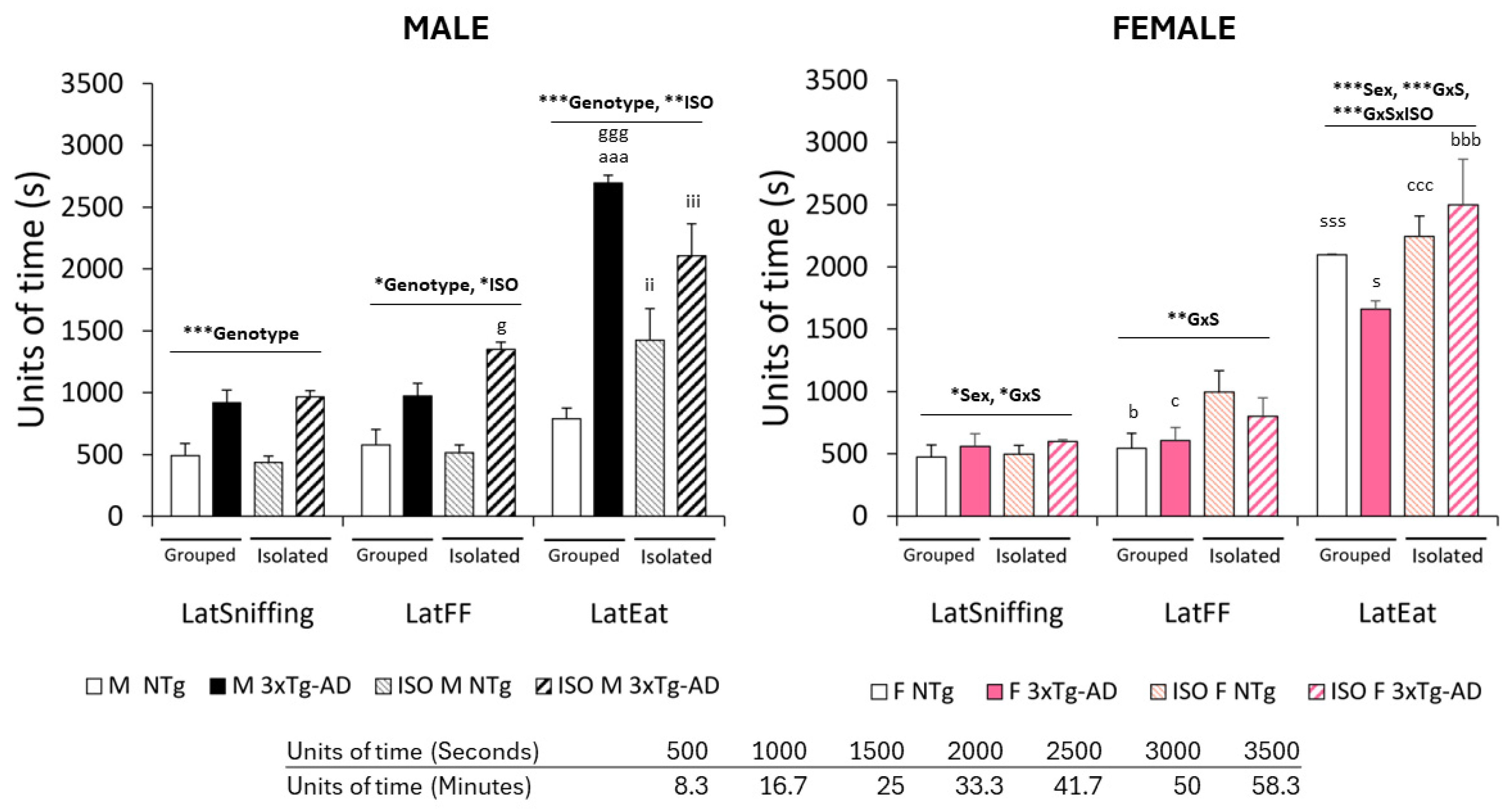
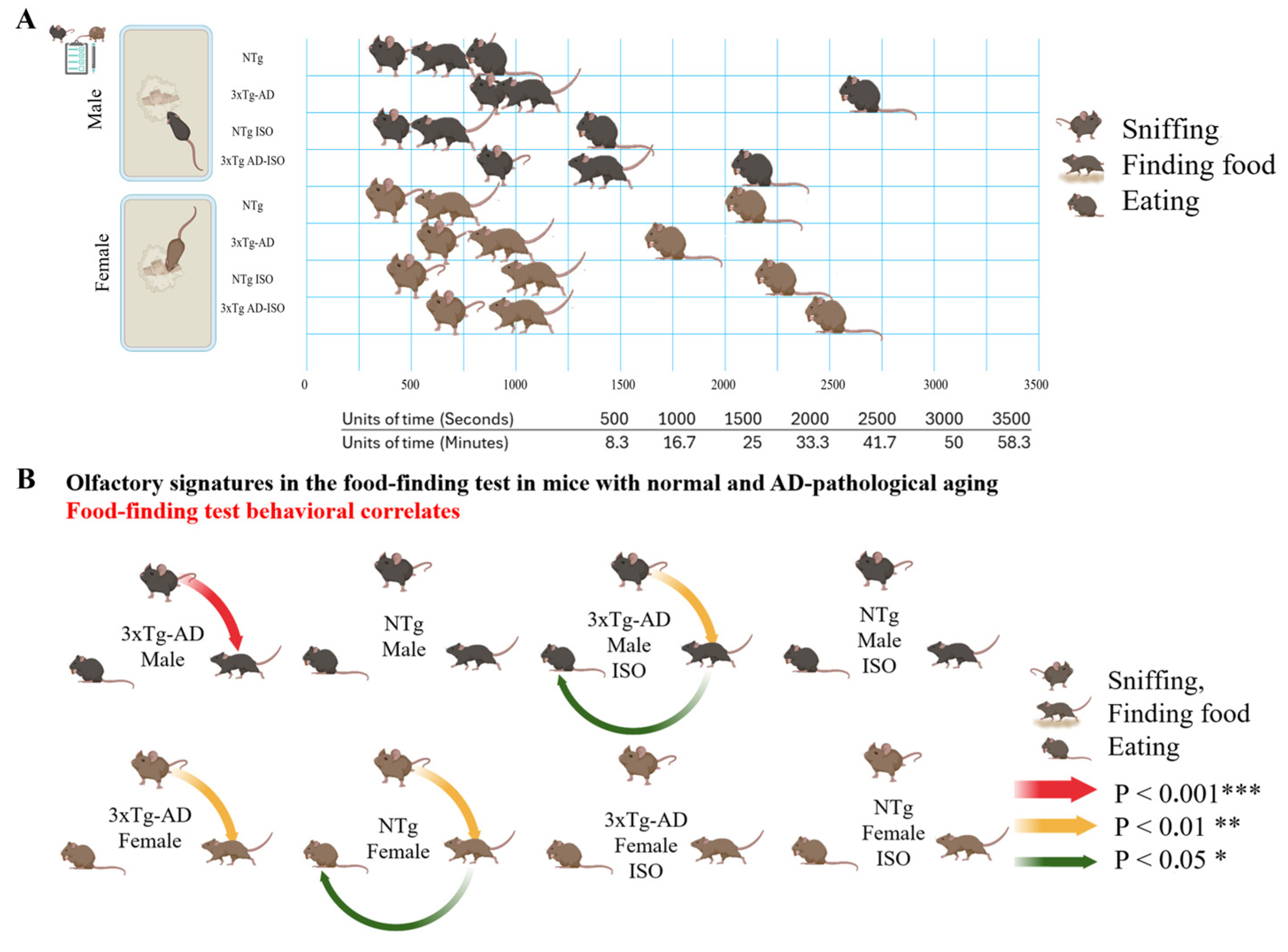
3.2. Elicitation of Olfactory Signatures despite the Lack of Overnight Food Deprivation
3.3. Time Delays and FFT Behavioral Correlates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Volkers, K.M.; Scherder, E.J.A. Impoverished Environment, Cognition, Aging and Dementia. Revneuro 2011, 22, 259–266. [Google Scholar] [CrossRef]
- Dziechciaż, M.; Filip, R. Biological Psychological and Social Determinants of Old Age: Bio-Psycho-Social Aspects of Human Aging. Ann. Agric. Environ. Med. 2014, 21, 835–838. [Google Scholar] [CrossRef]
- Murphy, C. Olfactory and Other Sensory Impairments in Alzheimer Disease. Nat. Rev. Neurol. 2019, 15, 11–24. [Google Scholar] [CrossRef]
- Devanand, D.P.; Lee, S.; Manly, J.; Andrews, H.; Schupf, N.; Doty, R.L.; Stern, Y.; Zahodne, L.B.; Louis, E.D.; Mayeux, R. Olfactory Deficits Predict Cognitive Decline and Alzheimer Dementia in an Urban Community. Neurology 2015, 84, 182–189. [Google Scholar] [CrossRef]
- Kondo, K.; Kikuta, S.; Ueha, R.; Suzukawa, K.; Yamasoba, T. Age-Related Olfactory Dysfunction: Epidemiology, Pathophysiology, and Clinical Management. Front. Aging Neurosci. 2020, 12, 208. [Google Scholar] [CrossRef]
- Van Regemorter, V.; Hummel, T.; Rosenzweig, F.; Mouraux, A.; Rombaux, P.; Huart, C. Mechanisms Linking Olfactory Impairment and Risk of Mortality. Front. Neurosci. 2020, 14, 140. [Google Scholar] [CrossRef]
- Wilson, R.S.; Arnold, S.E.; Schneider, J.A.; Boyle, P.A.; Buchman, A.S.; Bennett, D.A. Olfactory Impairment in Presymptomatic Alzheimer’s Disease. Ann. N. Y. Acad. Sci. 2009, 1170, 730–735. [Google Scholar] [CrossRef]
- Conti, M.Z.; Vicini-Chilovi, B.; Riva, M.; Zanetti, M.; Liberini, P.; Padovani, A.; Rozzini, L. Odor Identification Deficit Predicts Clinical Conversion from Mild Cognitive Impairment to Dementia Due to Alzheimer’s Disease. Arch. Clin. Neuropsychol. 2013, 28, 391–399. [Google Scholar] [CrossRef]
- Invitto, S.; Piraino, G.; Ciccarese, V.; Carmillo, L.; Caggiula, M.; Trianni, G.; Nicolardi, G.; Di Nuovo, S.; Balconi, M. Potential Role of OERP as Early Marker of Mild Cognitive Impairment. Front. Aging Neurosci. 2018, 10, 272. [Google Scholar] [CrossRef]
- Zou, Y.; Lu, D.; Liu, L.-P.; Zhang, H.; Zhou, Y. Olfactory Dysfunction in Alzheimer’s Disease. Neuropsychiatr. Dis. Treat. 2016, 12, 869–875. [Google Scholar] [CrossRef]
- Brai, E.; Hummel, T.; Alberi, L. Smell, an Underrated Early Biomarker for Brain Aging. Front. Neurosci. 2020, 14, 559918. [Google Scholar] [CrossRef]
- Oddo, S.; Caccamo, A.; Shepherd, J.D.; Murphy, M.P.; Golde, T.E.; Kayed, R.; Metherate, R.; Mattson, M.P.; Akbari, Y.; LaFerla, F.M. Triple-Transgenic Model of Alzheimer’s Disease with Plaques and Tangles. Neuron 2003, 39, 409–421. [Google Scholar] [CrossRef]
- Giménez-Llort, L.; Blázquez, G.; Cañete, T.; Johansson, B.; Oddo, S.; Tobeña, A.; LaFerla, F.M.; Fernández-Teruel, A. Modeling Behavioral and Neuronal Symptoms of Alzheimer’s Disease in Mice: A Role for Intraneuronal Amyloid. Neurosci. Biobehav. Rev. 2007, 31, 125–147. [Google Scholar] [CrossRef]
- Wersinger, S.R.; Caldwell, H.K.; Martinez, L.; Gold, P.; Hu, S.-B.; Young, W.S., 3rd. Vasopressin 1a Receptor Knockout Mice Have a Subtle Olfactory Deficit but Normal Aggression. Genes. Brain Behav. 2007, 6, 540–551. [Google Scholar] [CrossRef]
- Deacon, R.M.J.; Koros, E.; Bornemann, K.D.; Rawlins, J.N.P. Aged Tg2576 Mice Are Impaired on Social Memory and Open Field Habituation Tests. Behav. Brain Res. 2009, 197, 466–468. [Google Scholar] [CrossRef]
- Marín-Pardo, D.; Giménez-Llort, L. Olfactory Signatures in the Food Finding Test in Mice With Normal and Alzheimer’s Disease-Pathological Aging With Special Concerns on the Effects of Social Isolation. Front. Neurosci. 2021, 15, 733984. [Google Scholar] [CrossRef]
- Welker, W.I. Analysis of Sniffing of the Albino Rat 1. Behaviour 1964, 22, 223–244. [Google Scholar] [CrossRef]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving Bioscience Research Reporting: The ARRIVE Guidelines for Reporting Animal Research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef]
- Coronas-Sámano, G.; Portillo, W.; Beltrán Campos, V.; Medina-Aguirre, G.I.; Paredes, R.G.; Diaz-Cintra, S. Deficits in Odor-Guided Behaviors in the Transgenic 3xTg-AD Female Mouse Model of Alzheimer’s Disease. Brain Res. 2014, 1572, 18–25. [Google Scholar] [CrossRef]
- Slotnick, B.; Coppola, D.M. Odor-Cued Taste Avoidance: A Simple and Robust Test of Mouse Olfaction. Chem. Senses 2015, 40, 269–278. [Google Scholar] [CrossRef]
- Dan, X.; Wechter, N.; Gray, S.; Mohanty, J.G.; Croteau, D.L.; Bohr, V.A. Olfactory Dysfunction in Aging and Neurodegenerative Diseases. Ageing Res. Rev. 2021, 70, 101416. [Google Scholar] [CrossRef]
- Belfiore, R.; Rodin, A.; Ferreira, E.; Velazquez, R.; Branca, C.; Caccamo, A.; Oddo, S. Temporal and Regional Progression of Alzheimer’s Disease-like Pathology in 3xTg-AD Mice. Aging Cell 2019, 18, e12873. [Google Scholar] [CrossRef]
- Muntsant, A.; Castillo-Ruiz, M.d.M.; Giménez-Llort, L. Survival Bias, Non-Lineal Behavioral and Cortico-Limbic Neuropathological Signatures in 3xTg-AD Mice for Alzheimer’s Disease from Premorbid to Advanced Stages and Compared to Normal Aging. Int. J. Mol. Sci. 2023, 24, 13796. [Google Scholar] [CrossRef]
- Giménez-Llort, L.; Marin-Pardo, D.; Marazuela, P.; Hernández-Guillamón, M. Survival Bias and Crosstalk between Chronological and Behavioral Age: Age-and Genotype-Sensitivity Tests Define Behavioral Signatures in Middle-Aged, Old, and Long-Lived Mice with Normal and AD-Associated Aging. Biomedicines 2021, 9, 636. [Google Scholar] [CrossRef]
- Guayerbas, N.; Sánchez, A.I.; Gamallo, A.; De la Fuente, M. Mouse Performance in an Exploratory Activity Test as a Longevity Biomarker. Anim. Res. Welf. A Partnersh. 2000, 159–161. [Google Scholar]
- Slotnick, B.; Cockerham, R.; Pickett, E. Olfaction in Olfactory Bulbectomized Rats. J. Neurosci. 2004, 24, 9195–9200. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| NTg | ISO NTg | 3xTg-AD | ISO 3xTg-AD | ||
|---|---|---|---|---|---|
| Mean ± SEM | Mean ± SEM | Mean ± SEM | Mean ± SEM | ||
| Body Weight | Males | 46.90 ± 2.18 | 44.53 ± 1.52 | 33.60 ± 1.08 *** | 30.97 ± 0.50 *** |
| Factor: G**, S* | Females | 30.23 ± 1.77 | 30.41 ± 1.24 | 27.89 ± 0.97 | 24.49 ± 0.76 ** |
| Sniffing—Finding | Males | 130.2 ± 83.5 | 54.3 ± 34.0 | 80.4 ± 63.4 | 382.2 ± 192.4 |
| Factor: **ISO | Females | 70.0 ± 35.2 | 498.8 ± 183.4 | 48.7 ± 42.2 | 200.4 ± 152.7 |
| Finding—Eating | Males | 552.7 ± 112.4 | 1720.0 ± 142.0 | 906.6 ± 274.5 | 757.4 ± 240.5 |
| Factor: **S | Females | 1556.6 ± 289.6 | 1253.1 ± 255.2 | 1055.3 ± 233.9 | 1697.0 ± 319.9 |
| Sniffing—Eating | Males | 422.5 ± 111.0 | 1774.3 ± 133.9 | 987.0 ± 264.0 | 1139.6 ± 256.0 |
| Factor: **G × S | Females | 1626.6 ± 292.5 | 1751.9 ± 198.2 | 1104.0 ± 241.1 | 1897.4 ± 358.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marín-Pardo, D.; Giménez-Llort, L. Food Finding Test without Deprivation: A Sensorial Paradigm Sensitive to Sex, Genotype, and Isolation Shows Signatures of Derangements in Old Mice with Alzheimer’s Disease Pathology and Normal Aging. Brain Sci. 2024, 14, 288. https://doi.org/10.3390/brainsci14030288
Marín-Pardo D, Giménez-Llort L. Food Finding Test without Deprivation: A Sensorial Paradigm Sensitive to Sex, Genotype, and Isolation Shows Signatures of Derangements in Old Mice with Alzheimer’s Disease Pathology and Normal Aging. Brain Sciences. 2024; 14(3):288. https://doi.org/10.3390/brainsci14030288
Chicago/Turabian StyleMarín-Pardo, Daniela, and Lydia Giménez-Llort. 2024. "Food Finding Test without Deprivation: A Sensorial Paradigm Sensitive to Sex, Genotype, and Isolation Shows Signatures of Derangements in Old Mice with Alzheimer’s Disease Pathology and Normal Aging" Brain Sciences 14, no. 3: 288. https://doi.org/10.3390/brainsci14030288
APA StyleMarín-Pardo, D., & Giménez-Llort, L. (2024). Food Finding Test without Deprivation: A Sensorial Paradigm Sensitive to Sex, Genotype, and Isolation Shows Signatures of Derangements in Old Mice with Alzheimer’s Disease Pathology and Normal Aging. Brain Sciences, 14(3), 288. https://doi.org/10.3390/brainsci14030288