
The Hypothermic Influence on CHOP and Ero1-α in an Endoplasmic Reticulum Stress Model of Cerebral Ischemia

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Animals and Cell Lines
2.2. Global Cerebral Ischemia
2.3. Thapsigargin and Hypoxia Stressed PC12 Cells
2.4. RNA Extraction and Complementary DNA (cDNA) Synthesis
2.5. Quantitative-PCR
2.6. Statistical Analysis
3. Results
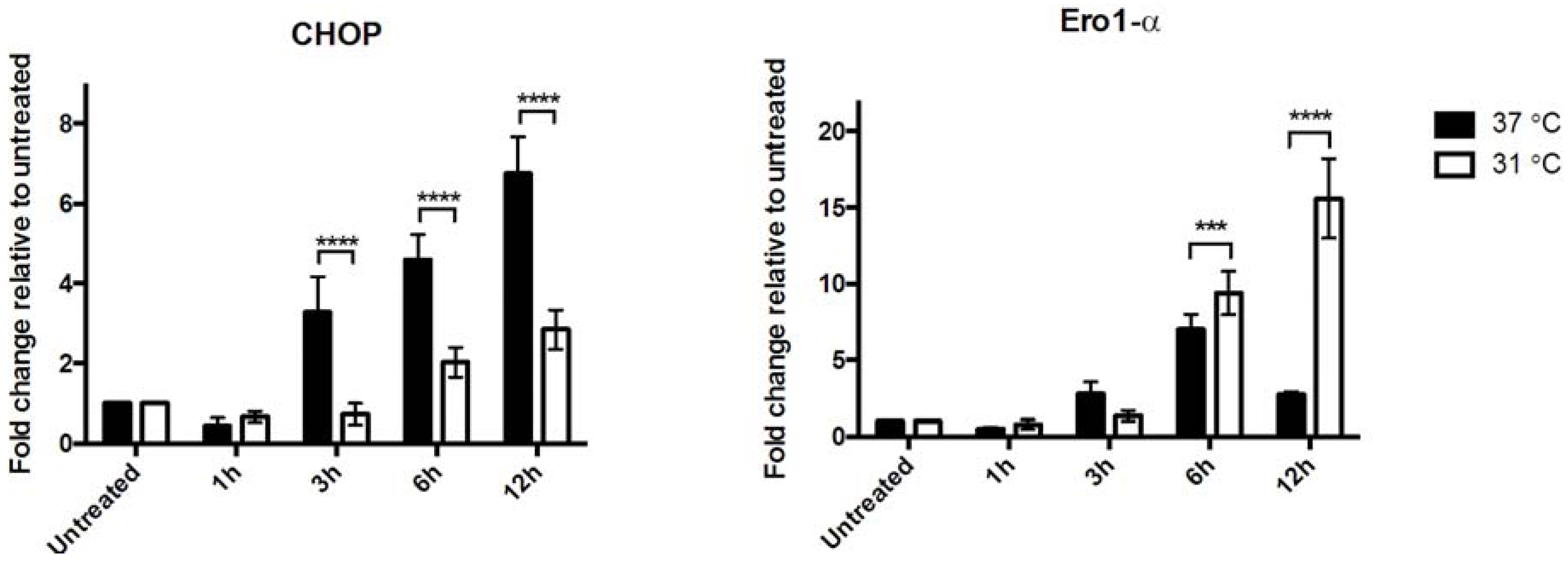
3.1. Global Cerebral Ischemia Induces Up-regulation of CHOP and Ero1-α in Rat Hippocampi

3.2. Thapsigargin and Verification of PC12 Cell Suitability

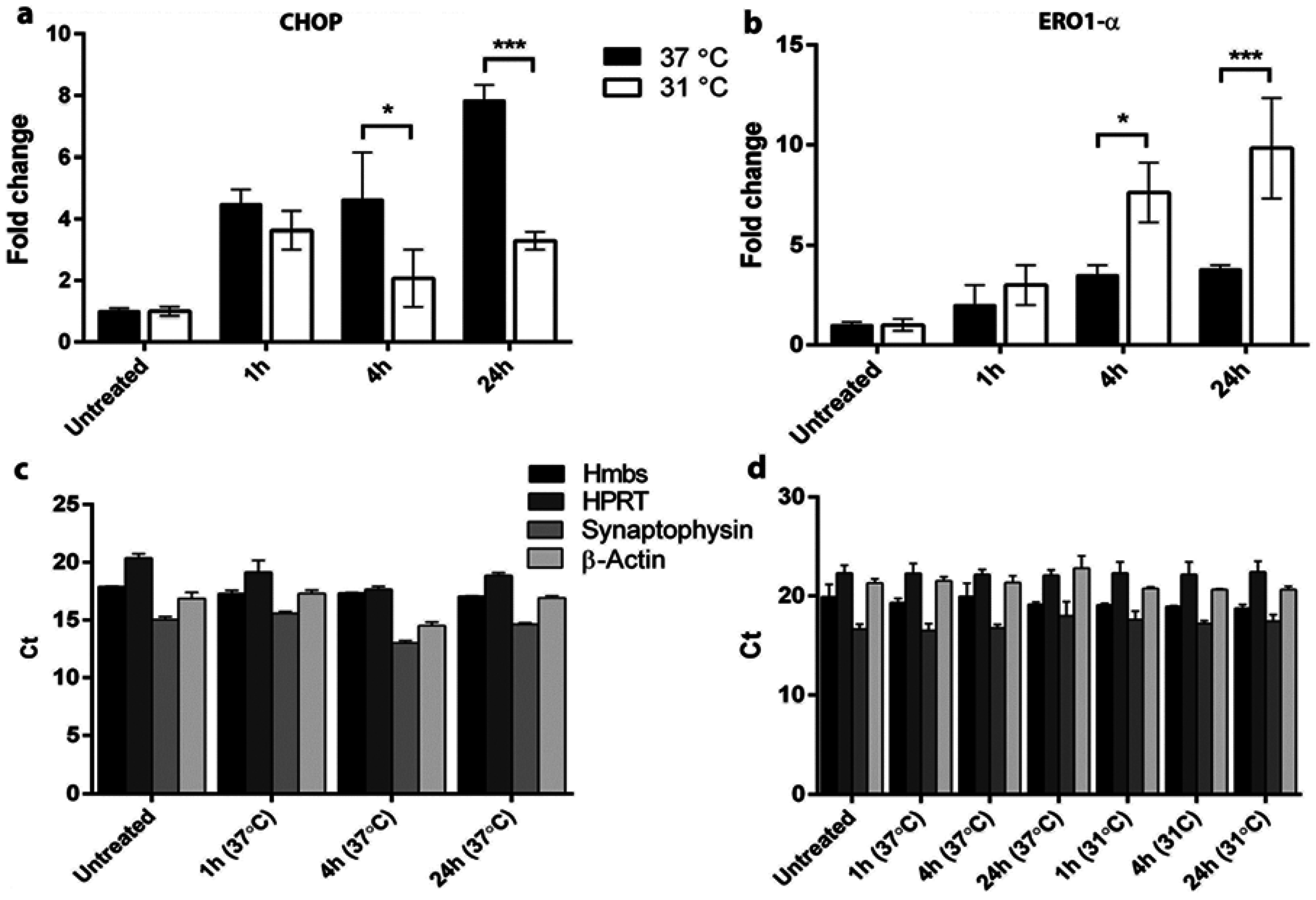
3.3. Hypothermia Attenuates CHOP and Augments Ero1-α Expression in Hypoxic Cells in Vitro

4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jiang, J.Y.; Xu, W.; Li, W.P.; Gao, G.Y.; Bao, Y.H.; Liang, Y.M.; Luo, Q.Z. Effect of long-term mild hypothermia or short-term mild hypothermia on outcome of patients with severe traumatic brain injury. J. Cereb. Blood Flow Metab. 2006, 26, 771–776. [Google Scholar] [CrossRef]
- Mottillo, S.; Sharma, K.; Eisenberg, M.J. Therapeutic hypothermia in acute myocardial infarction: A systematic review. Can. J. Cardiol. 2011, 27, 555–561. [Google Scholar] [CrossRef]
- England, T.N. Mild therapeutic hypothermia to improve the neurologic outcome after cardiac arrest. N. Engl. J. Med. 2002, 346, 549–556. [Google Scholar] [CrossRef]
- Hong, J.M.; Lee, J.S.; Song, H.J.; Jeong, H.; Choi, H.A.; Lee, K. Therapeutic hypothermia after recanalization in patients with acute ischemic stroke. Stroke 2014, 45, 134–140. [Google Scholar] [CrossRef]
- Krieger, D.W.; Yenari, M.A. Therapeutic hypothermia for acute ischemic stroke: What do laboratory studies teach us? Stroke 2004, 35, 1482–1489. [Google Scholar] [CrossRef]
- Ohta, H.; Terao, Y.; Shintani, Y.; Kiyota, Y. Therapeutic time window of post-ischemic mild hypothermia and the gene expression associated with the neuroprotection in rat focal cerebral ischemia. Neurosci. Res. 2007, 57, 424–433. [Google Scholar] [CrossRef]
- Wang, J.; Chambers, G.; Cottrell, J.E.; Kass, I.S. Differential Fall in ATP Accounts for Effects of Temperature on Hypoxic Damage in Rat Hippocampal Slices. J. Neurophysiol. 2000, 83, 3462–3472. [Google Scholar] [PubMed]
- Zhao, H.; Steinberg, G.K.; Sapolsky, R.M. General versus specific actions of mild-moderate hypothermia in attenuating cerebral ischemic damage. J. Cereb. Blood Flow Metab. 2007, 27, 1879–1894. [Google Scholar] [CrossRef]
- Liu, L.; Kim, J.Y.; Koike, M.A.; Yoon, Y.J.; Tang, X.N.; Ma, H.; Lee, H.; Steinberg, G.K.; Lee, J.E.; Yenari, M.A. FasL shedding is reduced by hypothermia in experimental stroke. J. Neurochem. 2008, 106, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Tajiri, S.; Oyadomari, S.; Yano, S.; Morioka, M.; Gotoh, T.; Hamada, J.I.; Ushio, Y.; Mori, M. Ischemia-induced neuronal cell death is mediated by the endoplasmic reticulum stress pathway involving CHOP. Cell Death Differ. 2004, 11, 403–415. [Google Scholar] [CrossRef]
- Szegezdi, E.; Logue, S.E.; Gorman, A.M.; Samali, A. Mediators of endoplasmic reticulum stress-induced apoptosis. EMBO Rep. 2006, 7, 880–885. [Google Scholar] [CrossRef] [PubMed]
- Harding, H.P.; Zhang, Y.; Zeng, H.; Novoa, I.; Lu, P.D.; Calfon, M.; Sadri, N.; Yun, C.; Popko, B.; Paules, R.; et al. An integrated stress response regulates amino acid metabolism and resistance to oxidative stress. Mol. Cell 2003, 11, 619–633. [Google Scholar]
- Ni, M.; Lee, A.S. ER chaperones in mammalian development and human diseases. FEBS Lett. 2007, 581, 3641–3651. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.R.; Martone, M.E.; Jones, Y.Z.; Liu, C.L. Protein aggregation after transient cerebral ischemia. J. Neurosci. 2000, 20, 3191–3199. [Google Scholar] [PubMed]
- Marciniak, S.J.; Yun, C.Y.; Oyadomari, S.; Novoa, I.; Zhang, Y.; Jungreis, R.; Nagata, K.; Harding, H.P.; Ron, D. CHOP induces death by promoting protein synthesis and oxidation in the stressed endoplasmic reticulum. Genes Dev. 2004, 18, 3066–3077. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Mongillo, M.; Chin, K.T.; Harding, H.; Ron, D.; Marks, A.R.; Tabas, I. Role of ERO1-alpha-mediated stimulation of inositol 1,4,5-triphosphate receptor activity in endoplasmic reticulum stress-induced apoptosis. J. Cell Biol. 2009, 186, 783–792. [Google Scholar] [CrossRef]
- Kalliokoski, O.; Abelson, K.S.; Koch, J.; Boschian, A.; Thormose, S.F.; Fauerby, N.; Rasmussen, R.S.; Johansen, F.F.; Hau, J. The effect of voluntarily ingested buprenorphine on rats subjected to surgically induced global cerebral ischaemia. In Vivo 2010, 24, 641–646. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Liu, X.; Wang, M.; Chen, H.; Guo, Y.; Ma, F.; Shi, F.; Bi, Y.; Li, Y. Hypothermia protects the brain from transient global ischemia/reperfusion by attenuating endoplasmic reticulum response-induced apoptosis through CHOP. PLoS ONE 2013, 8, e53431. [Google Scholar] [CrossRef] [PubMed]
- Aoki, M.; Tamatani, M.; Taniguchi, M.; Yamaguchi, A.; Bando, Y.; Kasai, K.; Miyoshi, Y.; Nakamura, Y.; Vitek, M.P.; Tohyama, M.; et al. Hypothermic treatment restores glucose regulated protein 78 (GRP78) expression in ischemic brain. Mol. Brain Res. 2001, 95, 117–128. [Google Scholar]
- Xu, S.Y.; Hu, Y.F.; Li, W.P.; Wu, Y.M.; Ji, Z.; Wang, S.N.; Li, K.; Pan, S.Y. Intermittent Hypothermia Is Neuroprotective in an in vitro Model of Ischemic Stroke. Int. J. Biol. Sci. 2014, 10, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Gess, B.; Hofbauer, K.H.; Wenger, R.H.; Lohaus, C.; Meyer, H.E.; Kurtz, A. The cellular oxygen tension regulates expression of the endoplasmic oxidoreductase ERO1-Lα. Eur. J. Biochem. 2003, 2235, 2228–2235. [Google Scholar] [CrossRef] [Green Version]
- May, D.; Itin, A.; Gal, O.; Kalinski, H.; Feinstein, E.; Keshet, E. Ero1-L alpha plays a key role in a HIF-1-mediated pathway to improve disulfide bond formation and VEGF secretion under hypoxia: Implication for cancer. Oncogene 2005, 24, 1011–1020. [Google Scholar] [CrossRef]
- Raghubir, R.; Nakka, V.P.; Mehta, S.L. Endoplasmic reticulum stress in brain damage. Methods Enzymol. 2011, 489, 259–275. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poone, G.K.; Hasseldam, H.; Munkholm, N.; Rasmussen, R.S.; Grønberg, N.V.; Johansen, F.F. The Hypothermic Influence on CHOP and Ero1-α in an Endoplasmic Reticulum Stress Model of Cerebral Ischemia. Brain Sci. 2015, 5, 178-187. https://doi.org/10.3390/brainsci5020178
Poone GK, Hasseldam H, Munkholm N, Rasmussen RS, Grønberg NV, Johansen FF. The Hypothermic Influence on CHOP and Ero1-α in an Endoplasmic Reticulum Stress Model of Cerebral Ischemia. Brain Sciences. 2015; 5(2):178-187. https://doi.org/10.3390/brainsci5020178
Chicago/Turabian StylePoone, Gagandip K., Henrik Hasseldam, Nina Munkholm, Rune S. Rasmussen, Nina V. Grønberg, and Flemming F. Johansen. 2015. "The Hypothermic Influence on CHOP and Ero1-α in an Endoplasmic Reticulum Stress Model of Cerebral Ischemia" Brain Sciences 5, no. 2: 178-187. https://doi.org/10.3390/brainsci5020178
APA StylePoone, G. K., Hasseldam, H., Munkholm, N., Rasmussen, R. S., Grønberg, N. V., & Johansen, F. F. (2015). The Hypothermic Influence on CHOP and Ero1-α in an Endoplasmic Reticulum Stress Model of Cerebral Ischemia. Brain Sciences, 5(2), 178-187. https://doi.org/10.3390/brainsci5020178