
Propofol-Induced Neurotoxicity in the Fetal Animal Brain and Developments in Modifying These Effects—An Updated Review of Propofol Fetal Exposure in Laboratory Animal Studies

{kind=link}
Abstract
:1. Introduction
2. Current Understanding of Anesthetic Neurotoxicity
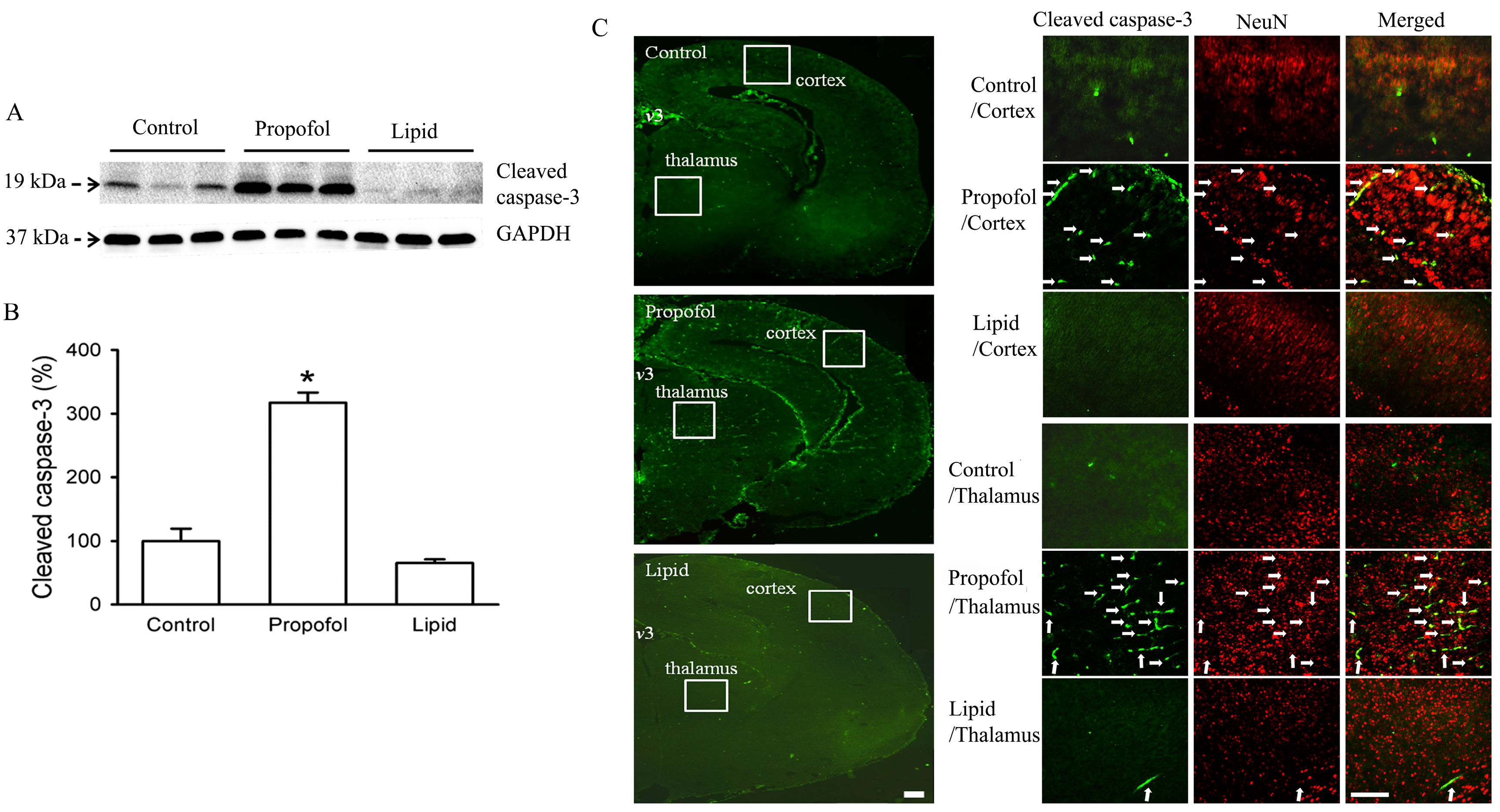
3. Effects of Propofol on Neonatal Animal Brain
4. Effects of Propofol in Fetal Animal Brain
5. Reversal Agents: Their Use in Understanding the Neurotoxic Effects of Propofol Exposure in a Fetal Brain
6. Conclusions
Conflicts of Interest
References
- Uemura, E.; Levin, E.D.; Bowman, R.E. Effects of halothane on synaptogenesis and learning behavior in rats. Exp. Neurol. 1985, 89, 520–529. [Google Scholar] [CrossRef]
- Chalon, J.; Tang, C.K.; Ramanathan, S.; Eisner, M.; Katz, R.; Turndorf, H. Exposure to halothane and enflurane affects learning function of murine progeny. Anesth. Analg. 1981, 60, 794–797. [Google Scholar] [CrossRef] [PubMed]
- Jevtovic-Todorovic, V.; Hartman, R.E.; Izumi, Y.; Benshoff, N.D.; Dikranian, K.; Zorumski, C.F.; Olney, J.W.; Wozniak, D.F. Early exposure to common anesthetic agents causes widespread neurodegeneration in the developing rat brain and persistent learning deficits. J. Neurosci. 2003, 23, 876–882. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Gao, J.; Karlsson, N.; Li, Q.; Zhang, Y.; Huang, Z.; Li, H.; Kuhn, H.G.; Blomgren, K. Isoflurane anesthesia induced persistent, progressive memory impairment, caused a loss of neural stem cells, and reduced neurogenesis in young, but not adult, rodents. J. Cereb. Blood Flow Metab. 2010, 30, 1017–1030. [Google Scholar] [CrossRef] [PubMed]
- Lunardi, N.; Ori, C.; Erisir, A.; Jevtovic-Todorovic, V. General anesthesia causes long-lasting disturbances in the ultrastructural properties of developing synapses in young rats. Neurotox. Res. 2010, 17, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Stratmann, G.; Sall, J.W.; May, L.D.; Bell, J.S.; Magnusson, K.R.; Rau, V.; Visrodia, K.H.; Alvi, R.S.; Ku, B.; Lee, M.T.; et al. Isoflurane differentially affects neurogenesis and long-term neurocognitive function in 60-day-old and 7-day-old rats. Anesthesiology 2009, 110, 834–848. [Google Scholar] [CrossRef] [PubMed]
- Satomoto, M.; Satoh, Y.; Terui, K.; Miyao, H.; Takishima, K.; Ito, M.; Imaki, J. Neonatal exposure to sevoflurane induces abnormal social behaviors and deficits in fear conditioning in mice. Anesthesiology 2009, 110, 628–637. [Google Scholar] [CrossRef] [PubMed]
- Slikker, W., Jr.; Zou, X.; Hotchkiss, C.E.; Divine, R.L.; Sadovova, N.; Twaddle, N.C.; Doerge, D.R.; Scallet, A.C.; Patterson, T.A.; Hanig, J.P.; et al. Ketamine-induced neuronal cell death in the perinatal rhesus monkey. Toxicol. Sci. 2007, 98, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Brambrink, A.M.; Evers, A.S.; Avidan, M.S.; Farber, N.B.; Smith, D.J.; Zhang, X.; Dissen, G.A.; Creeley, C.E.; Olney, J.W.; et al. Isoflurane-induced neuroapoptosis in the neonatal rhesus macaque brain. Anesthesiology 2010, 112, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Mazoit, J.X.; Roulleau, P.; Baujard, C. Isoflurane-induced neuroapoptosis in the neonatal rhesus macaque brain: Isoflurane or ischemia-reperfusion? Anesthesiology 2010, 113, 1245. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Dikkes, P.; Soriano, S.G. Repeated administration of ketamine may lead to neuronal degeneration in the developing rat brain. Paediatr. Anaesth. 2002, 12, 770–774. [Google Scholar] [CrossRef] [PubMed]
- Dobbing, J.; Sands, J. Comparative aspects of the brain growth spurt. Early Hum. Dev. 1979, 3, 79–83. [Google Scholar] [CrossRef]
- Bayer, S.A.; Altman, J.; Russo, R.J.; Zhang, X. Timetables of neurogenesis in the human brain based on experimentally determined patterns in the rat. Neurotoxicology 1993, 14, 83–144. [Google Scholar] [PubMed]
- Wang, H.; Luo, Z.; Xue, Z.G.; Cang, J. Propofol inhibits neuronal differentiation of mouse embryonic stem cells in vitro. Chin. Med. J. 2013, 126, 4186–4188. [Google Scholar] [PubMed]
- Al-Jahdari, W.S.; Saito, S.; Nakano, T.; Goto, F. Propofol induces growth cone collapse and neurite retractions in chick explant culture. Can. J. Anaesth. 2006, 53, 1078–1085. [Google Scholar] [CrossRef] [PubMed]
- Briner, A.; Nikonenko, I.; de Roo, M.; Dayer, A.; Muller, D.; Vutskits, L. Developmental stage-dependent persistent impact of propofol anesthesia on dendritic spines in the rat medial prefrontal cortex. Anesthesiology 2011, 115, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Pesic, V.; Milanovic, D.; Tanic, N.; Popic, J.; Kanazir, S.; Jevtovic-Todorovic, V.; Ruzdijić, S. Potential mechanism of cell death in the developing rat brain induced by propofol anesthesia. Int. J. Dev. Neurosci. 2009, 27, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Jiang, Y.; Gao, J.; Liu, B.; Chen, P. Repeated exposure to propofol potentiates neuroapoptosis and long-term behavioral deficits in neonatal rats. Neurosci. Lett. 2013, 534, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Fredriksson, A.; Ponten, E.; Gordh, T.; Eriksson, P. Neonatal exposure to a combination of N-methyl-d-aspartate and gamma-aminobutyric acid type A receptor anesthetic agents potentiates apoptotic neurodegeneration and persistent behavioral deficits. Anesthesiology 2007, 107, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Cattano, D.; Young, C.; Straiko, M.M.; Olney, J.W. Subanesthetic doses of propofol induce neuroapoptosis in the infant mouse brain. Anesth. Analg. 2008, 106, 1712–1714. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.M.; Qin, Y.; Liao, C.J.; Xie, Y.B.; Lan, Y.Y. Effects of propofol on expression of hippocampal survivin and Caspase-3 in newborn rats. Zhonghua Er Ke Za Zhi 2012, 50, 361–365. (In Chinese) [Google Scholar] [PubMed]
- Pearn, M.L.; Hu, Y.; Niesman, I.R.; Patel, H.H.; Drummond, J.C.; Roth, D.M.; Akassoglou, K.; Patel, P.M.; Head, B.P. Propofol neurotoxicity is mediated by p75 neurotrophin receptor activation. Anesthesiology 2012, 116, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Milanovic, D.; Pesic, V.; Popic, J.; Tanic, N.; Kanazir, S.; Jevtovic-Todorovic, V.; Ruždijić, S. Propofol anesthesia induces proapoptotic tumor necrosis factor-alpha and pro-nerve growth factor signaling and prosurvival Akt and XIAP expression in neonatal rat brain. J. Neurosci. Res. 2014, 92, 1362–1373. [Google Scholar] [CrossRef] [PubMed]
- Karen, T.; Schlager, G.W.; Bendix, I.; Sifringer, M.; Herrmann, R.; Pantazis, C.; Enot, D.; Keller, M.; Kerner, T.; Felderhoff-Mueser, U. Effect of propofol in the immature rat brain on short- and long-term neurodevelopmental outcome. PLoS ONE 2013, 8, e64480. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Liang, G.; Khojasteh, S.; Wu, Z.; Yang, W.; Joseph, D.; Wei, H. Comparison of neurodegeneration and cognitive impairment in neonatal mice exposed to propofol or isoflurane. PLoS ONE 2014, 9, e99171. [Google Scholar]
- Gao, J.; Peng, S.; Xiang, S.; Huang, J.; Chen, P. Repeated exposure to propofol impairs spatial learning, inhibits LTP and reduces CaMKIIalpha in young rats. Neurosci. Lett. 2014, 560, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Bercker, S.; Bert, B.; Bittigau, P.; Felderhoff-Muser, U.; Buhrer, C.; Ikonomidou, C.; Weise, M.; Kaisers, U.X.; Kerner, T. Neurodegeneration in newborn rats following propofol and sevoflurane anesthesia. Neurotox. Res. 2009, 16, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Creeley, C.; Dikranian, K.; Dissen, G.; Martin, L.; Olney, J.; Brambrink, A. Propofol-induced apoptosis of neurones and oligodendrocytes in fetal and neonatal rhesus macaque brain. Br. J. Anaesth. 2013, 110 (Suppl. 1), i29–i38. [Google Scholar] [CrossRef] [PubMed]
- Popic, J.; Pesic, V.; Milanovic, D.; Todorovic, S.; Kanazir, S.; Jevtovic-Todorovic, V.; Ruzdijic, S. Propofol-induced changes in neurotrophic signaling in the developing nervous system in vivo. PLoS ONE 2012, 7, e34396. [Google Scholar] [CrossRef] [PubMed]
- Kargaran, P.; Lenglet, S.; Montecucco, F.; Mach, F.; Copin, J.C.; Vutskits, L. Impact of propofol anaesthesia on cytokine expression profiles in the developing rat brain: An experimental study. Eur. J. Anaesthesiol. 2014, 31, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ikonomidou, C.; Bosch, F.; Miksa, M.; Bittigau, P.; Vockler, J.; Dikranian, K.; Tenkova, T.I.; Stefovska, V.; Turski, L.; Olney, J.W. Blockade of NMDA receptors and apoptotic neurodegeneration in the developing brain. Science 1999, 283, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.B.; Chen, G.; van den Pol, A.N. GABA-dependent firing of glutamate-evoked action potentials at AMPA/kainate receptors in developing hypothalamic neurons. J. Neurophysiol. 1998, 79, 716–726. [Google Scholar] [PubMed]
- Irifune, M.; Takarada, T.; Shimizu, Y.; Endo, C.; Katayama, S.; Dohi, T.; Kawahara, M. Propofol-induced anesthesia in mice is mediated by gamma-aminobutyric acid-A and excitatory amino acid receptors. Anesth. Analg. 2003, 97, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.; Rigo, J.M.; Rocher, V.; Belachew, S.; Malgrange, B.; Rogister, B.; Leprince, P.; Moonen, G. Neurotransmitters as early signals for central nervous system development. Cell Tissue Res. 2001, 305, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Herlenius, E.; Lagercrantz, H. Development of neurotransmitter systems during critical periods. Exp. Neurol. 2004, 190 (Suppl. 1), S8–S21. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xiong, M.; Nadavaluru, P.R.; Zuo, W.; Ye, J.H.; Eloy, J.D.; Bekker, A. Dexmedetomidine attenuates neurotoxicity induced by prenatal propofol exposure. J. Neurosurg. Anesthesiol. 2016, 28, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xiong, M.; Alhashem, H.M.; Zhang, Y.; Tilak, V.; Patel, A.; Siegel, A.; Ye, J.H.; Bekker, A. Effects of prenatal propofol exposure on postnatal development in rats. Neurotoxicol. Teratol. 2014, 43, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Pontén, E.; Fredriksson, A.; Gordh, T.; Eriksson, P.; Viberg, H. Neonatal exposure to propofol affects BDNF but not CaMKII, GAP-43, synaptophysin and tau in the neonatal brain and causes an altered behavioural response to diazepam in the adult mouse brain. Behav. Brain Res. 2011, 223, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.S.; Pontén, E.; Gordh, T.; Eriksson, P.; Fredriksson, A.; Sharma, A. Propofol promotes blood-brain barrier breakdown and heat shock protein (HSP 72 kd) activation in the developing mouse brain. CNS Neurol. Disord. Drug Targets 2014, 13, 1595–1603. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Yan, Y.; Canfield, S.; Muravyeva, M.Y.; Kikuchi, C.; Zaja, I.; Corbett, J.A.; Bosnjak, Z.J. Ketamine enhances human neural stem cell proliferation and induces neuronal apoptosis via reactive oxygen species-mediated mitochondrial pathway. Anesth. Analg. 2013, 116, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Dong, Y.; Xu, Z.; Crosby, G.; Culley, D.J.; Zhang, Y.; Xie, Z. Sevoflurane anesthesia in pregnant mice induces neurotoxicity in fetal and offspring mice. Anesthesiology 2013, 118, 516–526. [Google Scholar] [CrossRef] [PubMed]
- Schubert, H.; Eiselt, M.; Walter, B.; Fritz, H.; Brodhun, M.; Bauer, R. Isoflurane/nitrous oxide anesthesia and stress-induced procedures enhance neuroapoptosis in intrauterine growth-restricted piglets. Intensive Care Med. 2012, 38, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Xu, L.; He, D.; Zhang, X.; Lu, H. Effects of gestational isoflurane exposure on postnatal memory and learning in rats. Eur. J. Pharmacol. 2011, 670, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cheng, Y.; Liu, G.; Tian, X.; Tu, X.; Wang, J. Chronic exposure of gestation rat to sevoflurane impairs offspring brain development. Neurol. Sci. 2012, 33, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Alcaraz, A.; Quintana, M.B.; Laguarda, M. Placental transfer and neonatal effects of propofol in caesarean section. J. Clin. Pharm. Ther. 1998, 23, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Gin, T.; Gregory, M.A.; Chan, K.; Oh, T.E. Maternal and fetal levels of propofol at caesarean section. Anaesth. Intensive Care 1990, 18, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Xiong, M.; Li, J.; Alhashem, H.M.; Tilak, V.; Patel, A.; Pisklakov, S.; Siegel, A.; Ye, J.H.; Bekker, A. Propofol exposure in pregnant rats induces neurotoxicity and persistent learning deficit in the offspring. Brain Sci. 2014, 4, 356–375. [Google Scholar] [CrossRef] [PubMed]
- Straiko, M.M.; Young, C.; Cattano, D.; Creeley, C.E.; Wang, H.; Smith, D.J.; Johnson, S.A.; Li, E.S.; Olney, J.W. Lithium protects against anesthesia-induced developmental neuroapoptosis. Anesthesiology 2009, 110, 862–868. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, L.; Bennis, Y.; Velly, L.; Grandvuillemin, I.; Pisano, P.; Bruder, N.; Guillet, B. Erythropoietin protects newborn rat against sevoflurane-induced neurotoxicity. Paediatr. Anaesth. 2014, 24, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Degos, V.; Charpentier, T.L.; Chhor, V.; Brissaud, O.; Lebon, S.; Schwendimann, L.; Bednareck, N.; Passemard, S.; Mantz, J.; Gressens, P. Neuroprotective effects of dexmedetomidine against glutamate agonist-induced neuronal cell death are related to increased astrocyte brain-derived neurotrophic factor expression. Anesthesiology 2013, 118, 1123–1132. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.H.; Qiu, L.L.; Yang, J.J.; Zhang, H.; Sun, X.R.; Zhu, S.H.; Li, W.Y.; Yang, J.J. Pre-administration of curcumin prevents neonatal sevoflurane exposure-induced neurobehavioral abnormalities in mice. Neurotoxicology 2015, 46, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Man, Y.G.; Zhou, R.G.; Zhao, B. Efficacy of rutin in inhibiting neuronal apoptosis and cognitive disturbances in sevoflurane or propofol exposed neonatal mice. Int. J. Clin. Exp. Med. 2015, 8, 14397–14409. [Google Scholar] [PubMed]
- Paris, A.; Mantz, J.; Tonner, P.H.; Hein, L.; Brede, M.; Gressens, P. The effects of dexmedetomidine on perinatal excitotoxic brain injury are mediated by the alpha2A-adrenoceptor subtype. Anesth. Analg. 2006, 102, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Hossain, M.; Rajakumaraswamy, N.; Arshad, M.; Sanders, R.D.; Franks, N.P.; Maze, M. Dexmedetomidine produces its neuroprotective effect via the alpha 2A-adrenoceptor subtype. Eur. J. Pharmacol. 2004, 502, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Sanders, R.D.; Sun, P.; Patel, S.; Li, M.; Maze, M.; Ma, D. Dexmedetomidine provides cortical neuroprotection: Impact on anaesthetic-induced neuroapoptosis in the rat developing brain. Acta Anaesthesiol. Scand. 2010, 54, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Cao, D.; Han, X.; Liu, C.; Peng, J.; Zuo, Z.; Wang, F.; Li, Y. Both JNK and P38 MAPK pathways participate in the protection by dexmedetomidine against isoflurane-induced neuroapoptosis in the hippocampus of neonatal rats. Brain Res. Bull. 2014, 107, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.M.; Wang, C.C.; Chen, L.; Qian, L.B.; Ma, L.L.; Yu, J.; Zhu, M.H.; Wen, C.Y.; Yu, L.N.; Yan, M. Both PI3K/Akt and ERK1/2 pathways participate in the protection by dexmedetomidine against transient focal cerebral ischemia/reperfusion injury in rats. Brain Res. 2013, 1494, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zeng, M.; Chen, W.; Liu, C.; Wang, F.; Han, X.; Zuo, Z.; Peng, S. Dexmedetomidine reduces isoflurane-induced neuroapoptosis partly by preserving PI3K/Akt pathway in the hippocampus of neonatal rats. PLoS ONE 2014, 9, e93639. [Google Scholar]

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, M.; Zhang, L.; Li, J.; Eloy, J.; Ye, J.H.; Bekker, A. Propofol-Induced Neurotoxicity in the Fetal Animal Brain and Developments in Modifying These Effects—An Updated Review of Propofol Fetal Exposure in Laboratory Animal Studies. Brain Sci. 2016, 6, 11. https://doi.org/10.3390/brainsci6020011
Xiong M, Zhang L, Li J, Eloy J, Ye JH, Bekker A. Propofol-Induced Neurotoxicity in the Fetal Animal Brain and Developments in Modifying These Effects—An Updated Review of Propofol Fetal Exposure in Laboratory Animal Studies. Brain Sciences. 2016; 6(2):11. https://doi.org/10.3390/brainsci6020011
Chicago/Turabian StyleXiong, Ming, Li Zhang, Jing Li, Jean Eloy, Jiang Hong Ye, and Alex Bekker. 2016. "Propofol-Induced Neurotoxicity in the Fetal Animal Brain and Developments in Modifying These Effects—An Updated Review of Propofol Fetal Exposure in Laboratory Animal Studies" Brain Sciences 6, no. 2: 11. https://doi.org/10.3390/brainsci6020011
APA StyleXiong, M., Zhang, L., Li, J., Eloy, J., Ye, J. H., & Bekker, A. (2016). Propofol-Induced Neurotoxicity in the Fetal Animal Brain and Developments in Modifying These Effects—An Updated Review of Propofol Fetal Exposure in Laboratory Animal Studies. Brain Sciences, 6(2), 11. https://doi.org/10.3390/brainsci6020011