
Prenatal Alcohol Exposure Affects Progenitor Cell Numbers in Olfactory Bulbs and Dentate Gyrus of Vervet Monkeys




Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Histology
2.3. Immunohistochemistry
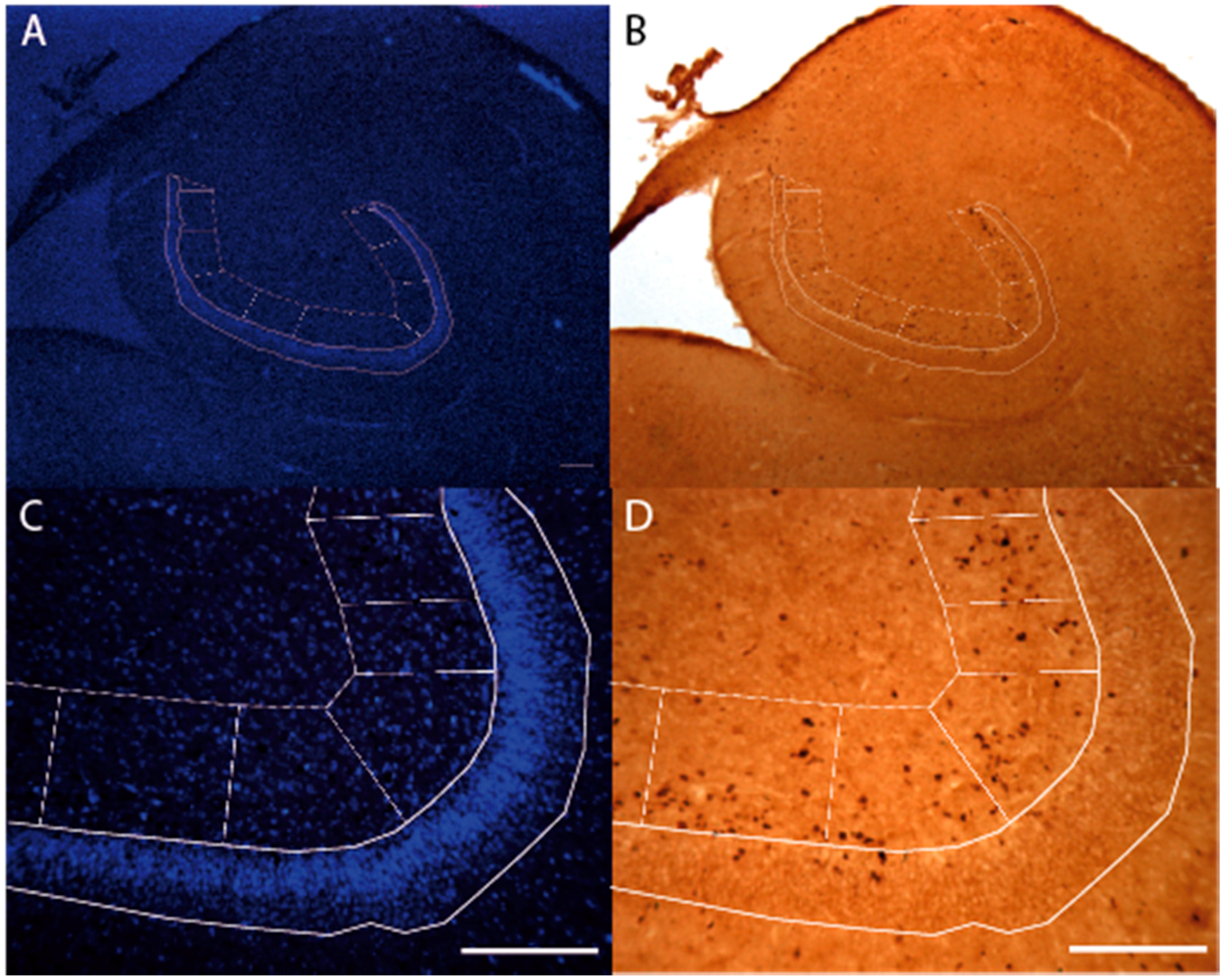
2.4. Stereology
2.5. Statistical Analysis
3. Results
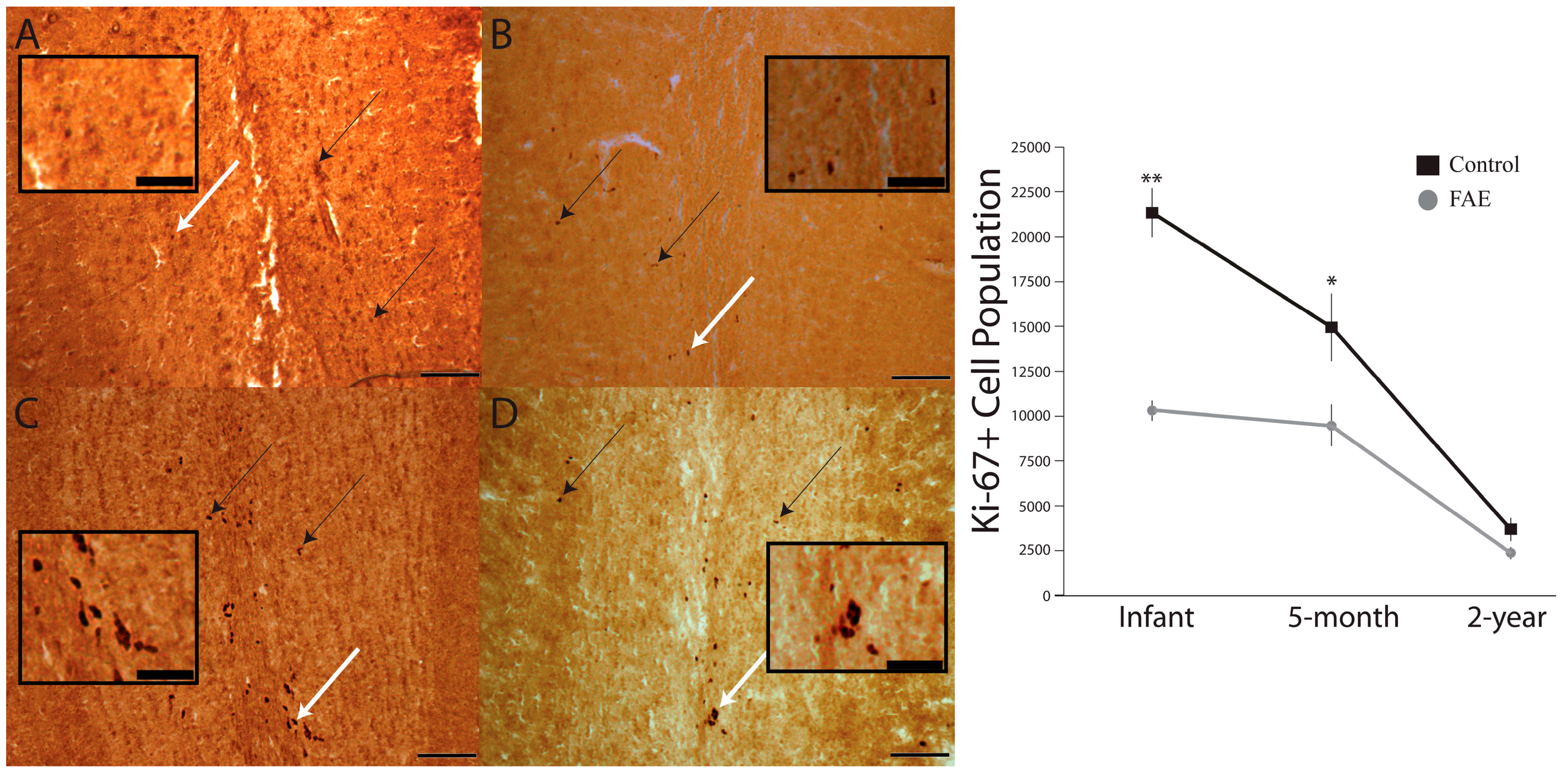
3.1. Olfactory Bulb
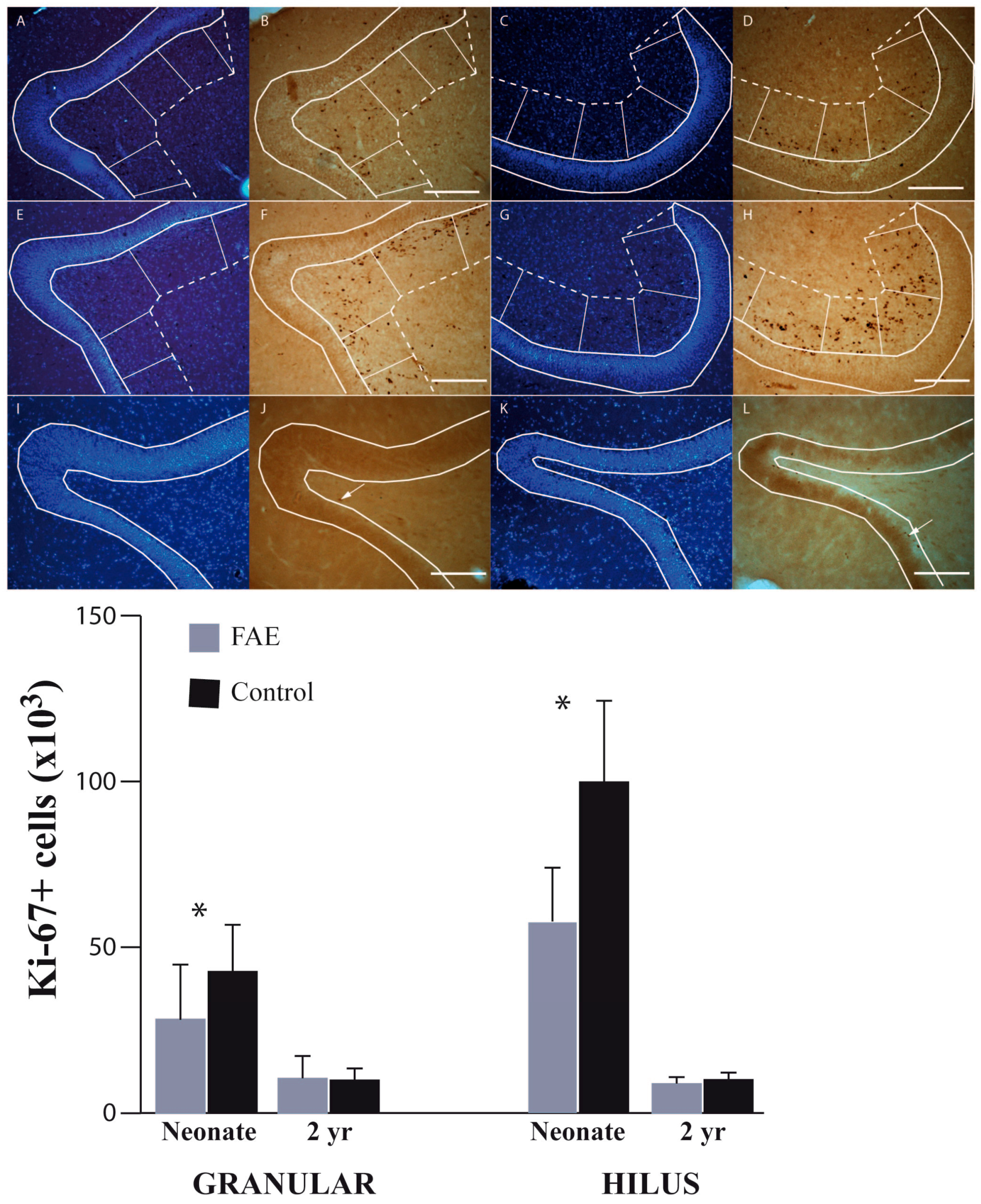
3.2. Dentate Gyrus
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tan, C.H.; Denny, C.H.; Cheal, N.E.; Sniezek, J.E.; Kanny, D. Alcohol use and binge drinking among women of childbearing age—United States. 2011–2013. MMWR Morb. Mortal. Wkly. Rep. 2015, 64, 1042–1046. [Google Scholar] [CrossRef] [PubMed]
- Streissguth, A.P.; Barr, H.M.; Sampson, P.D.; Bookstein, F.L. Prenatal alcohol and offspring development: The first fourteen years. Drug Alcohol Depend. 1994, 36, 89–99. [Google Scholar] [CrossRef]
- Mattson, S.N.; Crocker, N.; Nguyen, T.T. Fetal alcohol spectrum disorders: Neuropsychological and behavioral features. Neuropsychol. Rev. 2011, 21, 81–101. [Google Scholar] [CrossRef] [PubMed]
- Stratton, K.R.; Howe, C.I.; Battaglia, F.C.; Institite of Medicine (U.S.). Division of biobehavioral sciences and mental disorders, committee to study fetal alcohol syndrome. In Fetal Alcohol Syndrome: Diagnosis, Epidemiology, Prevention and Treatment; National Academy Press: Washington, DC, USA, 1996. [Google Scholar]
- Ericson, L.; Magnusson, L.; Horstadius, B. Societal costs of fetal alcohol syndrome in Sweden. Eur. J. Health Econ. 2016. [Google Scholar] [CrossRef] [PubMed]
- May, P.A.; Baete, A.; Russo, J.; Elliott, A.; Blankenship, J.; Kalberg, W.O.; Buckley, D.; Brooks, M.; Hasken, J.; Abdul-Rahman, O.; et al. Prevalence and characteristics of fetal alcohol spectrum disorders. Pediatrics 2014, 134, 855–866. [Google Scholar] [CrossRef] [PubMed]
- May, P.A.; Blankenship, J.; Marais, A.S.; Gossage, J.P.; Kalberg, W.O.; Barnard, R.; DeVries, M.; Robinson, L.K.; Adnams, C.M.; Buckley, D.; et al. Approaching the prevalence of the full spectrum of fetal alcohol spectrum disorders in a South African population-based study. Alcohol. Clin. Exp. Res. 2013, 37, 818–830. [Google Scholar] [CrossRef] [PubMed]
- Chudley, A.E.; Conry, J.; Cook, J.L.; Loock, C.; Rosales, T.; LeBlanc, N.; Public Health Agency of Canada’s National Advisory Committee on Fetal Alcohol Spectrum. Fetal alcohol spectrum disorder: Canadian guidelines for diagnosis. CMAJ 2005, 172, S1–S21. [Google Scholar] [CrossRef] [PubMed]
- Khan, B.A.; Robinson, R.F.; Smith, J.J.; Dillard, D.A. Prenatal alcohol exposure among Alaska native/American Indian infants. Int. J. Circumpolar Health 2013, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazotti, P.; George, M.A.; Szala-Meneok, K.; Morton, A.M.; Loock, C.; Van Bibber, M.; Ranford, J.; Fleming, M.; MacLeod, S. Preventing fetal alcohol spectrum disorder in aboriginal communities: A methods development project. PLoS Med. 2000, 3, e8. [Google Scholar] [CrossRef] [PubMed]
- Grant, T.M.; Higgins, J.E.; Sampson, P.D.; Ernst, C.C.; Barr, H.M.; Streissguth, A.P. Alcohol use before and during pregnancy in western Washington, 1989–2004: Implications for the prevention of fetal alcohol spectrum disorders. Am. J. Obstet. Gynecol. 2009, 200, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Lupton, C.; Burd, L.; Harwood, R. Cost of fetal alcohol spectrum disorders. Am. J. Med. Genet. C Semin. Med. Genet. 2004, 127, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Stade, B.; Ali, A.; Bennett, D.; Campbell, D.; Johnston, M.; Lens, C.; Tran, S.; Koren, G. The burden of prenatal exposure to alcohol: Revised measurement of cost. Can. J. Clin. Pharmacol. 2009, 16, e91–e102. [Google Scholar] [PubMed]
- Brown, N.N.; Burd, L.; Grant, T.; Edwards, W.; Adler, R.; Streissguth, A.P. Prenatal alcohol exposure: An assessment strategy for the legal context. Int. J. Law Psychiatry 2015, 42–43, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Streissguth, A.P.; O’Malley, K. Neuropsychiatric implications and long-term consequences of fetal alcohol spectrum disorders. Semin. Clin. Neuropsychiatry 2000, 5, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Olson, H.C.; Feldman, J.J.; Streissguth, A.P.; Sampson, P.D.; Bookstein, F.L. Neuropsychological deficits in adolescents with fetal alcohol syndrome: Clinical findings. Alcohol. Clin. Exp. Res. 1998, 22, 1998–2012. [Google Scholar] [CrossRef] [PubMed]
- Streissguth, A.P.; Bookstein, F.L.; Barr, H.M.; Sampson, P.D.; O’Malley, K.; Young, J.K. Risk factors for adverse life outcomes in fetal alcohol syndrome and fetal alcohol effects. J. Dev. Behav. Pediatr. 2004, 25, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Schonfeld, A.M.; Mattson, S.N.; Riley, E.P. Moral maturity and delinquency after prenatal alcohol exposure. J. Stud. Alcohol 2005, 66, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Kelly, S.J.; Day, N.; Streissguth, A.P. Effects of prenatal alcohol exposure on social behavior in humans and other species. Neurotoxicol. Teratol. 2000, 22, 143–149. [Google Scholar] [CrossRef]
- Baer, J.S.; Sampson, P.D.; Barr, H.M.; Connor, P.D.; Streissguth, A.P. A 21-year longitudinal analysis of the effects of prenatal alcohol exposure on young adult drinking. Arch. Gen. Psychiatry 2003, 60, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Connor, P.B.; Sampson, P.D.; Bookstein, F.L.; Barr, H.M.; Streissguth, A.P. Direct and indirect effects of prenatal alcohol damage on executive function. Dev. Neuropsychol. 2000, 18, 331–354. [Google Scholar] [CrossRef] [PubMed]
- Khoury, J.E.; Milligan, K.; Girard, T.A. Executive functioning in children and adolescents prenatally exposed to alcohol: A meta-analytic review. Neuropsychol. Rev. 2015, 25, 149–179. [Google Scholar] [CrossRef] [PubMed]
- Kerns, K.A.; Don, A.; Mateer, C.A.; Streissguth, A.P. Cognitive deficits in nonretarded adults with fetal alcohol syndrome. J. Learn. Disabil. 1997, 30, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Connor, P.D.; Streissguth, A.P.; Sampson, P.D.; Bookstein, F.L.; Barr, H.M. Individual differences in auditory and visual attention among fetal alcohol-affected adults. Alcohol. Clin. Exp. Res. 1999, 23, 1395–1402. [Google Scholar] [CrossRef] [PubMed]
- Connor, P.D.; Sampson, P.D.; Streissguth, A.P.; Bookstein, F.L.; Barr, H.M. Effects of prental alcohol exposure on fine motor coordination and balance. A study of two adult samples. Neuropsychologia 2006, 44, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Furtado, E.F.; Roriz, S.T. Inattention and impulsivity associated with prenatal alcohol exposure in a prospective cohort study of 11-year-old Brazilian children. Eur. Child Adolesc. Psychiatry 2016. [Google Scholar] [CrossRef] [PubMed]
- Astley, S.J.; Aylward, E.H.; Olson, H.C.; Kerns, K.; Brooks, A.; Coggins, T.E.; Davies, J.; Dorn, S.; Gendler, B.; Jirikowic, T.; et al. Functional magnetic imaging outcomes from a comprehensive magnetic resonance study of children with fetal alcohol spectrum disorders. J. Neurodev. Disord. 2009, 1, 61–80. [Google Scholar] [CrossRef] [PubMed]
- Nanson, J.L.; Hiscock, M. Attention deficits in children exposed to alcohol prenatally. Alcohol. Clin. Exp. Res. 1990, 14, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, K.; Brigman, J.L. The impact of prenatal alcohol exposure on social cognitive and affective domains: Insights from rodent models. Alcohol 2016, 51, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Donald, K.A.; Eastman, E.; Howells, F.M.; Adnams, C.; Riley, E.P.; Woods, R.P.; Narr, K.L.; Stein, D.J. Neuroimaging effects of prenatal alcohol exposure on the developing human brain: A magnetic resonance imaging review. Acta Neuropsychiatr. 2015, 27, 251–269. [Google Scholar] [CrossRef] [PubMed]
- Burke, M.W.; Palmour, R.M.; Ervin, F.R.; Ptito, M. Neuronal reduction in frontal cortex of primates after prenatal alcohol exposure. Neuroreport 2009, 20, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Burke, M.W.; Ptito, M.; Ervin, F.R.; Palmour, R.M. Hippocampal neuron populations are reduced in vervet monkeys with fetal alcohol exposure. Dev. Psychobiol. 2015, 57, 470–485. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.L.; Moore, C.F.; Adkins, M.M. The effects of prenatal alcohol exposure on behavior: Rodent and primate studies. Neuropsychol. Rev. 2011, 21, 186–203. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.; Warton, C.; Jacobson, S.W.; Jacobson, J.L.; Molteno, C.C.; Eicher, A.; Marais, P.; Phillips, O.R.; Narr, K.L.; Meintjes, E.M. Three-dimensional surface deformation-based shape analysis of hippocampus and caudate nucleus in children with fetal alcohol spectrum disorders. Hum. Brain Mapp. 2014, 35, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Astley, S.J.; Aylward, E.H.; Olson, H.C.; Kerns, K.; Brooks, A.; Coggins, T.E.; Davies, J.; Dorn, S.; Gendler, B.; Jirikowic, T.; et al. Magnetic resonance imaging outcomes from a comprehensive magnetic resonance study of children with fetal alcohol spectrum disorders. Alcohol. Clin. Exp. Res. 2009, 33, 1671–1689. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.D.; Kelly, S.J. Critical periods for ethanol induced cell loss in the hippocampal formation. Neurotoxicol. Teratol. 2003, 25, 519–528. [Google Scholar] [CrossRef]
- Curtis, M.A.; Kam, M.; Nannmark, U.; Anderson, M.F.; Axell, M.Z.; Wikkelso, C.; Holtas, S.; van Roon-Mom, W.M.; Bjork-Eriksson, T.; Nordborg, C.; et al. Human neuroblasts migrate to the olfactory bulb via a lateral ventricular extension. Science 2007, 315, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Gil-Mohapel, J.; Boehme, F.; Kainer, L.; Christie, B.R. Hippocampal cell loss and neurogenesis after fetal alcohol exposure: Insights from different rodent models. Brain Res. Rev. 2010, 64, 283–303. [Google Scholar] [CrossRef] [PubMed]
- Gil-Mohapel, J.; Boehme, F.; Patten, A.; Cox, A.; Kainer, L.; Giles, E.; Brocardo, P.S.; Christie, B.R. Altered adult hippocampal neuronal maturation in a rat model of fetal alcohol syndrome. Brain Res. 2011, 1384, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Klintsova, A.Y.; Helfer, J.L.; Calizo, L.H.; Dong, W.K.; Goodlett, C.R.; Greenough, W.T. Persistent impairment of hippocampal neurogenesis in young adult rats following early postnatal alcohol exposure. Alcohol. Clin. Exp. Res. 2007, 31, 2073–2082. [Google Scholar] [CrossRef] [PubMed]
- Ieraci, A.; Herrera, D.G. Single alcohol exposure in early life damages hippocampal stem/progenitor cells and reduces adult neurogenesis. Neurobiol. Dis. 2007, 26, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Gupta, S.; Jiang, Y.; Younus, M.; Ramzan, M. In vitro neurogenesis from neural progenitor cells isolated from the hippocampus region of the brain of adult rats exposed to ethanol during early development through their alcohol-drinking mothers. Alcohol Alcohol. 2009, 44, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Gould, E. How widespread is adult neurogenesis in mammals? Nat. Rev. Neurosci. 2007, 8, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Gil-Perotin, S.; Duran-Moreno, M.; Belzunegui, S.; Liquin, M.R.; Garcia-Verdugo, J.M. Ultrastructure of the subventricular zone in Macaca fascicularis and evidence of a mouse-like migratory stream. J. Comp. Neurol. 2009, 514, 533–554. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Kang, E.; Liu, C.Y.; Ming, G.L.; Song, H. Development of neural stem cells in the adult brain. Curr. Opin. Neurobiol. 2008, 18, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Yamashima, T.; Tonchev, A.B.; Yukie, M. Adult hippocampal neurogenesis in rodents and primates: Endogenous, enhanced and engrafted. Rev. Neurosci. 2007, 18, 67–82. [Google Scholar] [CrossRef] [PubMed]
- Rakic, P. Pre- and post-developmental neurogenesis in primates. Clin. Neurosci. Res. 2002, 2, 10. [Google Scholar] [CrossRef]
- Bedard, A.; Levesque, M.; Bernier, P.J.; Parent, A. The rostral migratory stream in adult squirrel monkeys: Contribution of new neurons to the olfactory tubercle and involvement of the antiapoptotic protein Bcl-2. Eur. J. Neurosci. 2002, 16, 1917–1924. [Google Scholar] [CrossRef] [PubMed]
- Gheusi, G.; Lledo, P.M. Control of early events in olfactory processing by adult neurogenesis. Chem. Senses 2007, 32, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Gheusi, G.; Lledo, P.M. Adult neurogenesis in the olfactory system shapes odor memory and perception. Prog. Brain Res. 2014, 208, 157–175. [Google Scholar] [PubMed]
- Gheusi, G.; Ortega-Perez, I.; Murray, K.; Lledo, P.M. A niche for adult neurogenesis in social behavior. Behav. Brain Res. 2009, 200, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Gil-Mohapel, J.; Titterness, A.K.; Patten, A.R.; Taylor, S.; Ratzlaff, A.; Ratzlaff, T.; Helfer, J.; Christie, B.R. Prenatal ethanol exposure differentially affects hippocampal neurogenesis in the adolescent and aged brain. Neuroscience 2014, 273, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Svanidze, I.K.; Museridze, D.P.; Didimova, E.V.; Sanikidze, T.V.; Gegnava, L.G.; Gvinadze, N.N. Disordered of neurogenesis of cortical and subcortical structures in rat brain limbic system during fetal alcohol syndrome formation. Morfologiia 2012, 141, 18–22. [Google Scholar] [PubMed]
- Choi, I.Y.; Allan, A.M.; Cunningham, L.A. Moderate fetal alcohol exposure impairs the neurogenic response to an enriched environment in adult mice. Alcohol. Clin. Exp. Res. 2005, 29, 2053–2062. [Google Scholar] [CrossRef] [PubMed]
- Huart, C.; Rombaux, P.; Hummel, T. Plasticity of the human olfactory system: The olfactory bulb. Molecules 2013, 18, 11586–11600. [Google Scholar] [CrossRef] [PubMed]
- Akers, K.G.; Kushner, S.A.; Leslie, A.T.; Clarke, L.; van der Kooy, D.; Lerch, J.P.; Frankland, P.W. Fetal alcohol exposure leads to abnormal olfactory bulb development and impaired odor discrimination in adult mice. Mol. Brain 2011, 4, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barron, S.; Riley, E.P. The effects of prenatal alcohol exposure on behavioral and neuroanatomical components of olfaction. Neurotoxicol. Teratol. 1992, 14, 291–297. [Google Scholar] [CrossRef]
- Maier, S.E.; Cramer, J.A.; West, J.R.; Sohrabji, F. Alcohol exposure during the first two trimesters equivalent alters granule cell number and neurotrophin expression in the developing rat olfactory bulb. J. Neurobiol. 1999, 41, 414–423. [Google Scholar] [CrossRef]
- Bothius, D.J.; Bonthius, N.E.; Napper, R.M.; West, J.R. Early postnatal alcohol exposure acutely and permanently reduces the number of granule cells and mitral cells in the rat olfactory bulb: A stereological study. J. Comp. Neurol. 1992, 324, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Gil-Perotin, S.; Alvarez-Buylla, A.; Garcia-Verdugo, J.M. Identification and characterization of neural progenitor cells in the adult mammalian brain. Adv. Anat. Embryol. Cell Biol. 2009, 203, 1–101. [Google Scholar] [PubMed]
- Eadie, B.D.; Redila, V.A.; Christie, B.R. Voluntary exercise alters the cytoarchitecture of the adult dentate gyrus by increasing cellular proliferation, dendritic complexity and spine density. J. Comp. Neurol. 2005, 486, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Scholzen, T.; Geredes, J. The Ki-67 protein: From the known and the unknown. J. Cell. Physiol. 2000, 182, 311–322. [Google Scholar] [CrossRef]
- West, M.J.; Gundersen, H.J. Unbiased stereological estimation of the number of neurons in the human hippocampus. J. Comp. Neurol. 1990, 296, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Papia, M.F.; Burke, M.W.; Zangenehpour, S.; Palmour, R.M.; Ervin, F.R.; Ptito, M. Reduced soma size of the M-neurons in the lateral geniculate nucleus following foetal alcohol exposure in non-human primates. Exp. Brain Res. 2010, 205, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Jabes, A.; Lavenex, P.B.; Amaral, D.G.; Lavenex, P. Quantitative analysis of postnatal neurogenesis and neuron number in the macaque monkey dentate gyrus. Eur. J. Neurosci. 2010, 31, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Kornack, D.R.; Rakic, P. The generation, migration and differentiation of olfactory neurons in the adult primate brain. Proc. Natl. Acad. Sci. USA 2001, 98, 4752–4757. [Google Scholar] [CrossRef] [PubMed]
- Pencea, V.; Bingaman, K.D.; Freedman, L.J.; Luskin, M.B. Neurogenesis in the subventricular zone and rostral migratory stream of the neonatal and adult primate forebrain. Exp. Neurol. 2001, 172, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Bedard, A.; Parent, A. Evidence of newly generated neurons in the human olfactory bulb. Brain Res. Dev. Brain Res. 2004, 151, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Enwere, E.; Shingo, T.; Gregg, C.; Fujikawa, H.; Ohta, S.; Weiss, S. Aging results in reduced epidermal growth factor receptor signaling, diminished olfactory neurogenesis and deficits in olfactory discrimination. J. Neurosci. 2004, 24, 8354–8365. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, G.C.; Barker, R.A.; Caldwell, M.A. Dopaminergic modulation of neurogenesis in the subventricular zone of the adult brain. Cell Cycle 2009, 8, 2888–2894. [Google Scholar] [CrossRef] [PubMed]
- Christie, B.R.; Swann, S.E.; Fox, C.J.; Froc, D.; Lieblich, S.E.; Redila, V.; Webber, A. Voluntary exercise rescues deficits in spatial memory and long-term potentiation in prenatal ethanol-exposed male rats. Eur. J. Neurosci. 2005, 21, 1719–1726. [Google Scholar] [CrossRef] [PubMed]
- Boehme, F.; Gil-Mohapel, J.; Cox, A.; Patten, A.; Giles, E.; Brocardo, P.S.; Christie, B.R. Voluntary exercise induces adult hippocampal neurogenesis and BDNF expression in a rodent model of fetal alcohol spectrum disorders. Eur. J. Neurosci. 2011, 33, 1799–1811. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, G.F.; Jablonski, S.A.; Schiffino, F.L.; St Cyr, S.A.; Stanton, M.E.; Klintsova, A.Y. Exercise and environment as an intervention for neonatal alcohol effects on hippocampal adult neurogenesis and learning. Neuroscience 2014, 265, 274–290. [Google Scholar] [CrossRef] [PubMed]
- Kajimoto, K.; Allan, A.; Cunningham, L.A. Fate analysis of adult hippocampal progenitors in a murine model of fetal alcohol spectrum disorder (FASD). PLoS ONE 2013, 8, e73788. [Google Scholar] [CrossRef] [PubMed]
- Roitbak, T.; Thomas, K.; Martin, A.; Allan, A.; Cunningham, L.A. Moderate fetal alcohol exposure impairs neurogenic capacity of murine neural stem cells isolated from the adult subventricular zone. Exp. Neurol. 2011, 229, 522–525. [Google Scholar] [CrossRef] [PubMed]
- Camarillo, C.; Miranda, R.C. Ethanol exposure during neurogenesis induces persistent effects on neural maturation: Evidence from an ex vivo model of fetal cerebral cortical neuroepithelial progenitor maturation. Gene Expr. 2008, 14, 159–171. [Google Scholar] [PubMed]
- Tyler, C.R.; Allan, A.M. Prenatal alcohol exposure alters expression of neurogenesis-related genes in an ex vivo cell culture model. Alcohol 2014, 48, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Alvarez, R.; Gayen, S.; Vadigepalli, R.; Anni, H. Ethanol diverts early neuronal differentiation trajectory of embryonic stem cells by disrupting the balance of lineage specifiers. PLoS ONE 2013, 8, e63794. [Google Scholar] [CrossRef] [PubMed]
- Anthony, B.; Zhou, F.C.; Ogawa, T.; Goodlett, C.R.; Ruiz, J. Alcohol exposure alters cell cycle and apoptotic events during early neurulation. Alcohol Alcohol. 2008, 43, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Santillano, D.R.; Kumar, L.S.; Prock, T.L.; Camarillo, C.; Tingling, J.D.; Miranda, R.C. Ethanol induces cell-cycle activity and reduces stem cell diversity to alter both regenerative capacity and differentiation potential of cerebral cortical neuroepithelial precursors. BMC Neurosci. 2005, 6, 59. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.C.; Balaraman, Y.; Teng, M.; Liu, Y.; Singh, R.P.; Nephew, K.P. Alcohol alters DNA methylation patterns and inhibits neural stem cell differentiation. Alcohol. Clin. Exp. Res. 2011, 35, 735–746. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Animal | Sex | Region | Day Alc Started | Drinking Days | Av Alc (g/kg/day) | Average BEC | Age at Sacrifice |
|---|---|---|---|---|---|---|---|
| O1808-1 | m | DG | 131 | 19 | 3.23 | 135 | 12 days |
| O2898-5 | m | DG | 96 | 40 | 3.19 | 108 | 15 days |
| O3245-3 | f | DG | 66 | 56 | 3.39 | 129 | 3 days |
| O3307-3 | f | OB/DG | 94 | 40 | 3.17 | 132 | 35 days |
| O5011-4 | m | OB/DG | 82 | 42 | 1.97 | 72 | 15 days |
| O2780-5 | m | OB | 93 | 36 | 3 | 118 | 10 days |
| O5232-2 | m | DG | 77 | 0 | sucrose | 0 | 9 days |
| O6228-1 | m | DG | 90 | 0 | sucrose | 0 | 18 days |
| O6332-1 | f | DG | 121 | 0 | sucrose | 0 | 12 days |
| O6692-1 | m | DG | 115 | 0 | sucrose | 0 | 30 days |
| O6712-1 | m | DG | 72 | 0 | sucrose | 0 | 1 days |
| O3295-5 | f | OB | 86 | 0 | sucrose | 0 | 15 days |
| O6172-1 | m | OB | 125 | 0 | sucrose | 0 | 10 days |
| O5329-1 | m | OB | 109 | 0 | sucrose | 0 | 6 days |
| O3065-8 | m | OB | 94 | 41 | 2.98 | 125 | 5 months |
| O5399-1 | m | OB | 106 | 35 | 2.48 | 101 | 5 months |
| O5859-1 | m | OB | 127 | 22 | 2.24 | 75 | 5 months |
| O5173-2 | f | OB | 118 | 0 | sucrose | 0 | 5 months |
| O6503-1 | m | OB | 103 | 0 | sucrose | 0 | 5 months |
| O9184-4-2 | m | OB | 108 | 0 | sucrose | 0 | 5 months |
| O3327-1 | m | DG | 115 | 30 | 2.98 | 120 | 21 months |
| O5011-3 | m | OB | 77 | 48 | 2.81 | 118 | 19 months |
| O3295-3 | m | OB/DG | 75 | 51 | 2.91 | 97 | 22 months |
| O3327-2 | m | OB/DG | 113 | 31 | 2.77 | 104 | 22 months |
| O2780-6 | m | OB | 113 | 30 | 2.98 | 95 | 26 months |
| O5603-2 | m | DG | 112 | 0 | sucrose | 0 | 22 months |
| O3060-5 | m | OB/DG | 116 | 0 | sucrose | 0 | 21 months |
| O5151-1 | m | OB/DG | 73 | 0 | sucrose | 0 | 19 months |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burke, M.W.; Inyatkin, A.; Ptito, M.; Ervin, F.R.; Palmour, R.M. Prenatal Alcohol Exposure Affects Progenitor Cell Numbers in Olfactory Bulbs and Dentate Gyrus of Vervet Monkeys. Brain Sci. 2016, 6, 52. https://doi.org/10.3390/brainsci6040052
Burke MW, Inyatkin A, Ptito M, Ervin FR, Palmour RM. Prenatal Alcohol Exposure Affects Progenitor Cell Numbers in Olfactory Bulbs and Dentate Gyrus of Vervet Monkeys. Brain Sciences. 2016; 6(4):52. https://doi.org/10.3390/brainsci6040052
Chicago/Turabian StyleBurke, Mark W., Alexey Inyatkin, Maurice Ptito, Frank R. Ervin, and Roberta M. Palmour. 2016. "Prenatal Alcohol Exposure Affects Progenitor Cell Numbers in Olfactory Bulbs and Dentate Gyrus of Vervet Monkeys" Brain Sciences 6, no. 4: 52. https://doi.org/10.3390/brainsci6040052
APA StyleBurke, M. W., Inyatkin, A., Ptito, M., Ervin, F. R., & Palmour, R. M. (2016). Prenatal Alcohol Exposure Affects Progenitor Cell Numbers in Olfactory Bulbs and Dentate Gyrus of Vervet Monkeys. Brain Sciences, 6(4), 52. https://doi.org/10.3390/brainsci6040052