
Seed Location Impacts Whole-Brain Structural Network Comparisons between Healthy Elderly and Individuals with Alzheimer’s Disease

Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Imaging Data
2.3. Network Construction
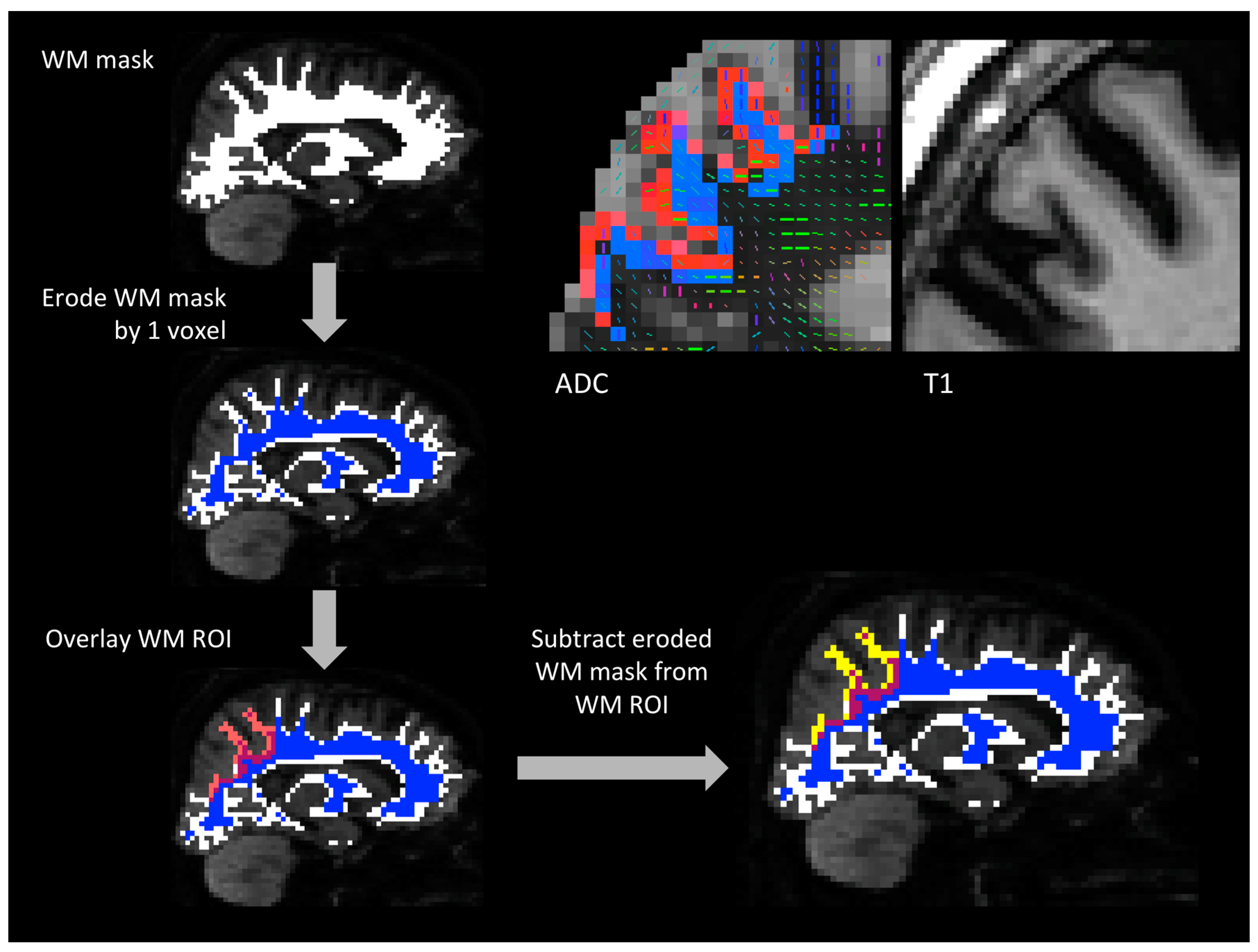
2.3.1. Parcellation of Gray Matter and White Matter
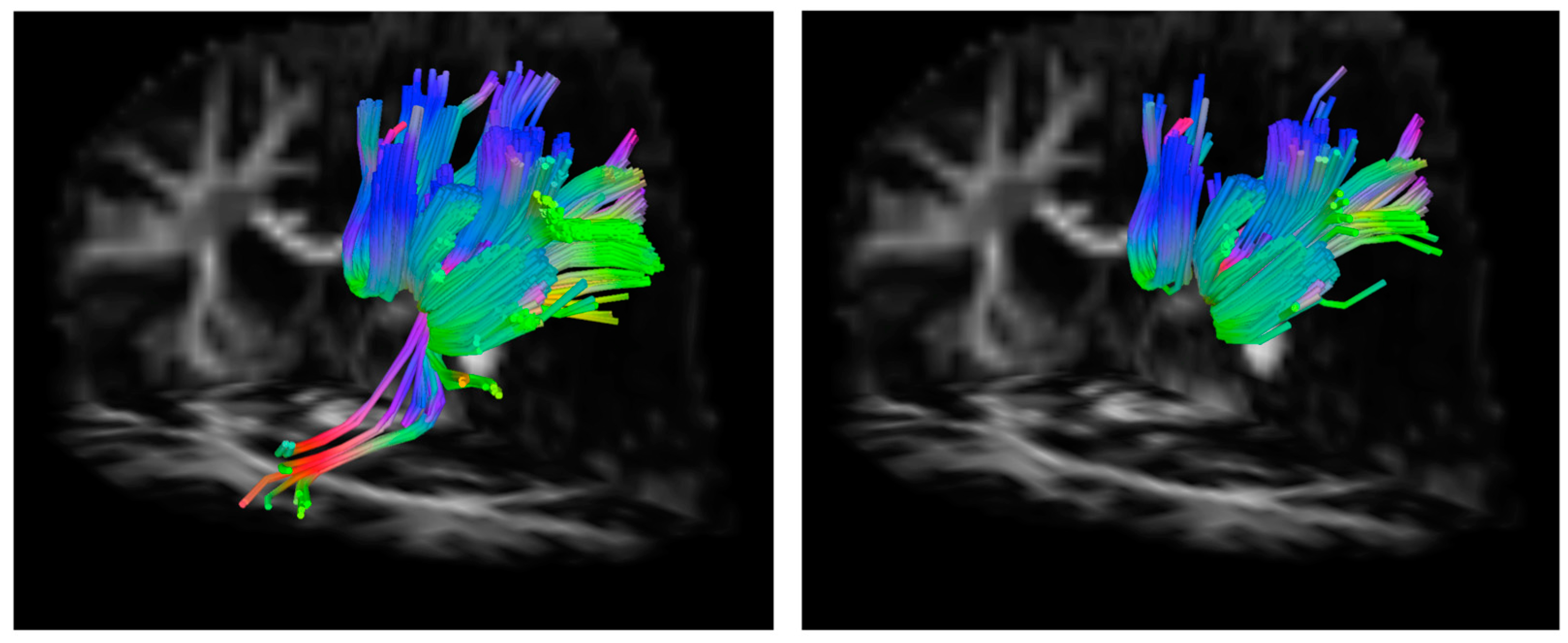
2.3.2. Tractography
2.4. Graph-Theory Network Measures
Whole-Network Measures
2.5. Statistics
2.5.1. Network Based Statistic (NBS)
2.6. Node-Based Measures
3. Results
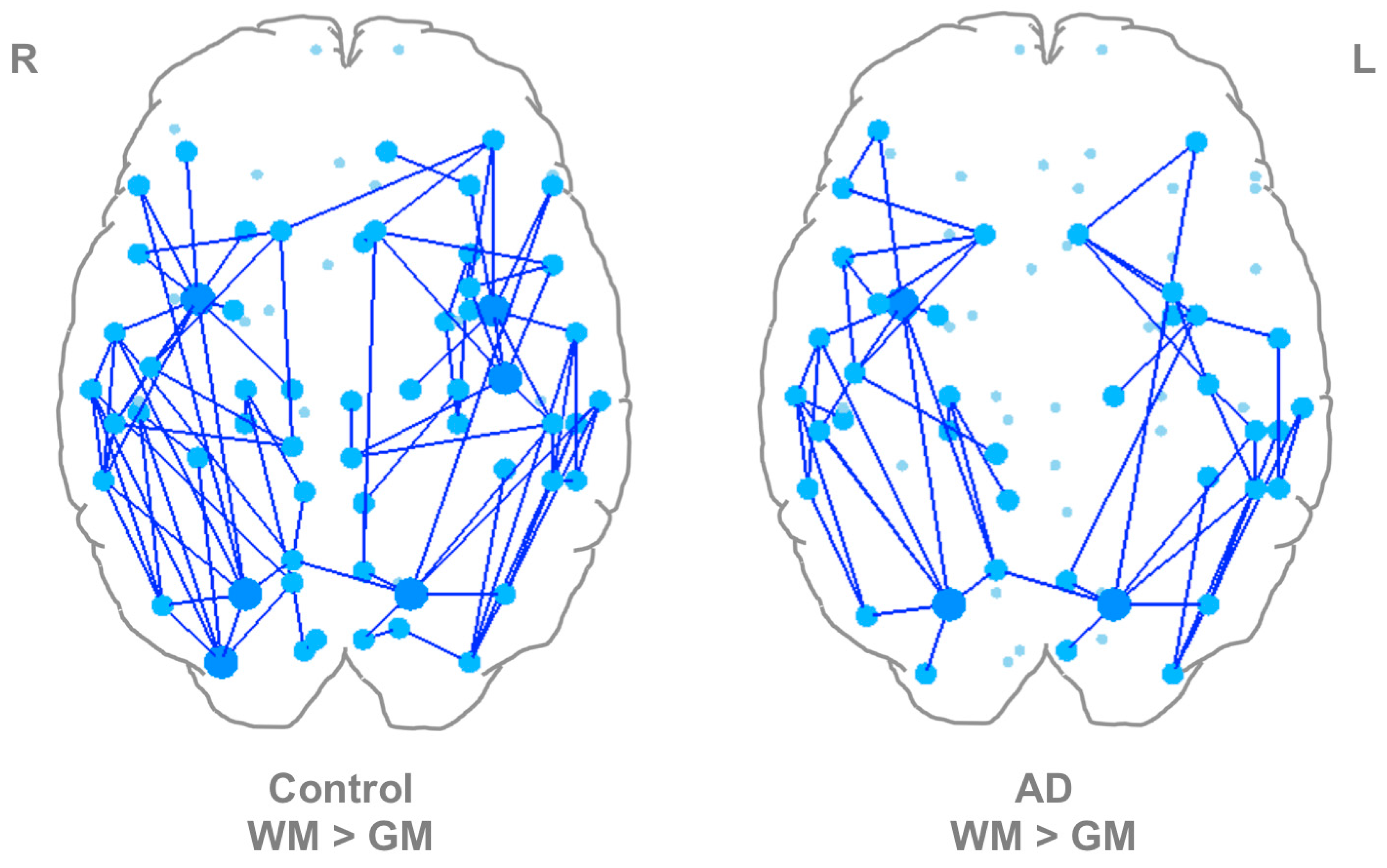
3.1. WM-Seed and GM-Seed Network Characterization and Comparisons
3.2. Within-Group Differences in WM-Seed versus GM-Seed Networks
3.3. WM-Seed and GM-Seed Weighted Whole-Network Measures
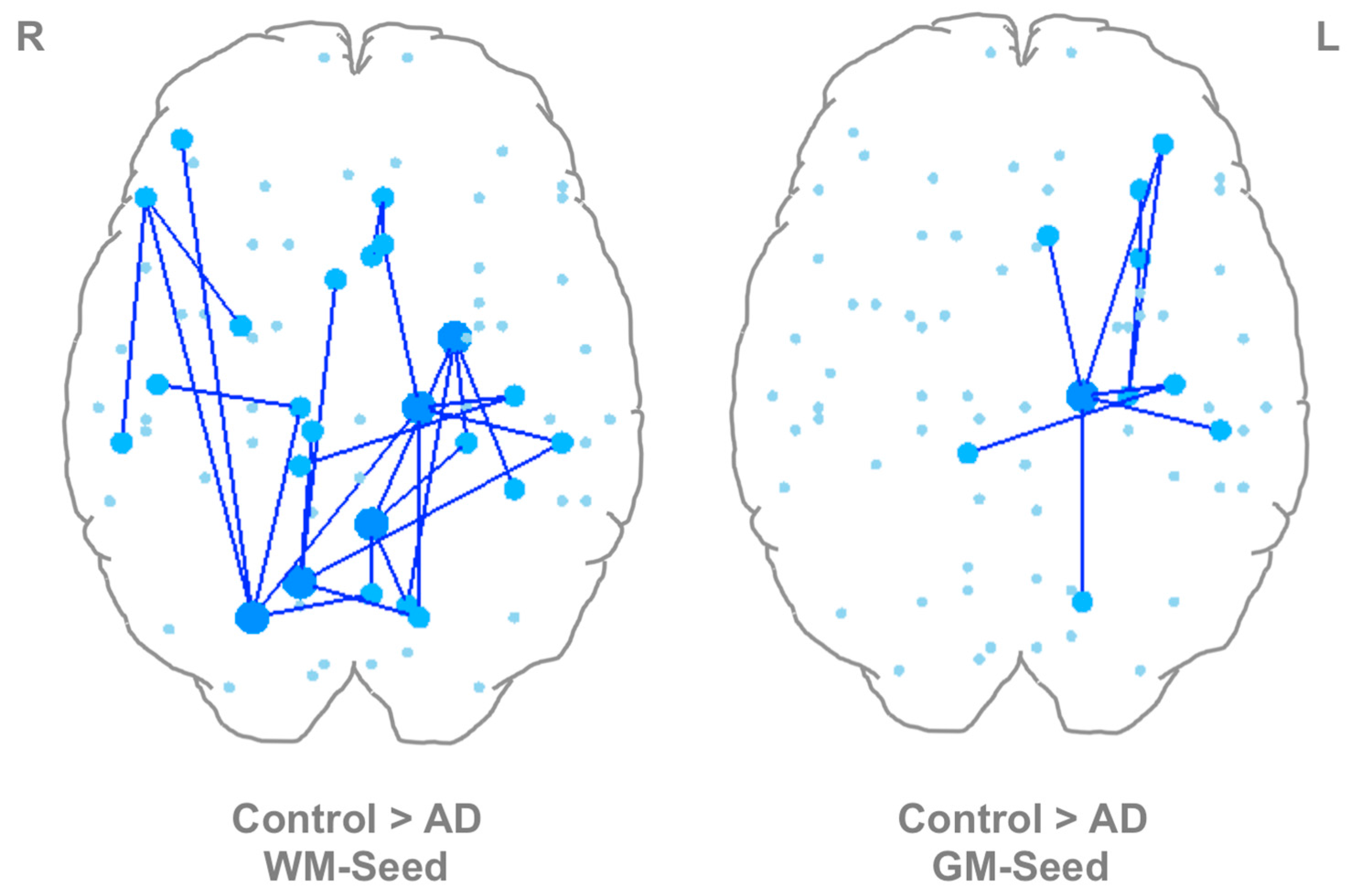
3.4. Between-Group Differences Using WM-Seed and GM-Seed Networks
3.5. Targeted Node-Based Differences between Controls and AD
4. Discussion
4.1. The Effects of Seed Location on Properties of WM-Seed and GM-Seed Networks
4.2. Differences between AD and Controls and the Effects of Seed Location on These Differences
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Basser, P.J.; Mattiello, J.; LeBihan, D. MR diffusion tensor spectroscopy and imaging. Biophys. J. 1994, 66, 259–267. [Google Scholar] [CrossRef]
- Mori, S.; Crain, B.J.; Chacko, V.; Van Zijl, P. Three-dimensional tracking of axonal projections in the brain by magnetic resonance imaging. Ann. Neurol. 1999, 45, 265–269. [Google Scholar] [CrossRef]
- Catani, M.; Howard, R.J.; Pajevic, S.; Jones, D.K. Virtual in vivo interactive dissection of white matter fasciculi in the human brain. Neuroimage 2002, 17, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Wakana, S.; Caprihan, A.; Panzenboeck, M.M.; Fallon, J.H.; Perry, M.; Gollub, R.L.; Hua, K.; Zhang, J.; Jiang, H.; Dubey, P.; et al. Reproducibility of quantitative tractography methods applied to cerebral white matter. Neuroimage 2007, 36, 630–644. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Schuff, N.; Jahng, G.H.; Bayne, W.; Mori, S.; Schad, L.; Mueller, S.; Du, A.T.; Kramer, J.H.; Weiner, M.W.; et al. Diffusion tensor imaging of cingulum fibers in mild cognitive impairment and Alzheimer disease. Neurology 2007, 68, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Iturria-Medina, Y.; Sotero, R.C.; Canales-Rodríguez, E.J.; Alemán-Gómez, Y.; Melie-García, L. Studying the human brain anatomical network via diffusion-weighted MRI and graph theory. Neuroimage 2008, 40, 1064–1076. [Google Scholar] [CrossRef] [PubMed]
- Hagmann, P.; Cammoun, L.; Gigandet, X.; Meuli, R.; Honey, C.J.; Wedeen, V.J.; Sporns, O. Mapping the structural core of human cerebral cortex. PLoS Biol. 2008, 6, e159. [Google Scholar] [CrossRef] [PubMed]
- Zalesky, A.; Fornito, A.; Harding, I.H.; Cocchi, L.; Yücel, M.; Pantelis, C.; Bullmore, E.T. Whole-brain anatomical networks: Does the choice of nodes matter? Neuroimage 2010, 50, 970–983. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Rilling, J.K.; Preuss, T.M.; Glasser, M.F.; Hu, X. The effects of connection reconstruction method on the interregional connectivity of brain networks via diffusion tractography. Hum. Brain Map. 2012, 33, 1894–1913. [Google Scholar] [CrossRef] [PubMed]
- Gigandet, X.; Griffa, A.; Kober, T.; Daducci, A.; Gilbert, G.; Connelly, A.; Hagmann, P.; Meuli, R.; Thiran, J.P.; Krueger, G. A connectome-based comparison of diffusion MRI schemes. PLoS ONE 2013, 8, e75061. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, C.R.; Pernet, C.R.; Gorgolewski, K.J.; Storkey, A.J.; Bastin, M.E. Test–retest reliability of structural brain networks from diffusion MRI. Neuroimage 2014, 86, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Parker, C.S.; Deligianni, F.; Cardoso, M.J.; Daga, P.; Modat, M.; Dayan, M.; Clark, C.A.; Ourselin, S.; Clayden, J.D. Consensus between pipelines in structural brain networks. PLoS ONE 2014, 9, e111262. [Google Scholar] [CrossRef] [PubMed]
- Zhan, L.; Zhou, J.; Wang, Y.; Jin, Y.; Jahanshad, N.; Prasad, G.; Nir, T.M.; Leonardo, C.D.; Ye, J.; Thompson, P.M. Comparison of nine tractography algorithms for detecting abnormal structural brain networks in Alzheimer’s disease. Front. Aging Neurosci. 2015, 7, 48. [Google Scholar] [PubMed]
- Qi, S.; Meesters, S.; Nicolay, K.; ter Haar Romeny, B.M.; Ossenblok, P. The influence of construction methodology on structural brain network measures: A review. J. Neurosci. Methods 2015, 253, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Craddock, R.C.; Jbabdi, S.; Yan, C.G.; Vogelstein, J.T.; Castellanos, F.X.; Di Martino, A.; Kelly, C.; Heberlein, K.; Colcombe, S.; Milham, M.P. Imaging human connectomes at the macroscale. Nat. Methods 2013, 10, 524–539. [Google Scholar] [CrossRef] [PubMed]
- De LaCoste, M.; White, C.L. The role of cortical connectivity in Alzheimer’s disease pathogenesis: A review and model system. Neurobiol. Aging 1993, 14, 1–16. [Google Scholar] [CrossRef]
- Delbeuck, X.; Van der Linden, M.; Collette, F. Alzheimer’s disease as a disconnection syndrome? Neuropsychol. Rev. 2003, 13, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.A.; Campbell, M.J.; Terry, R.D.; Morrison, J.H. Laminar and regional distributions of neurofibrillary tangles and neuritic plaques in Alzheimer’s disease: A quantitative study of visual and auditory cortices. J. Neurosci. Off. J. Soc. Neurosci. 1987, 7, 1799–1808. [Google Scholar]
- Arnold, S.E.; Hyman, B.T.; Flory, J.; Damasio, A.R.; Van Hoesen, G.W. The topographical and neuroanatomical distribution of neurofibrillary tangles and neuritic plaques in the cerebral cortex of patients with Alzheimer’s disease. Cereb. Cortex 1991, 1, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Hof, P.R.; Bouras, C.; Morrison, J.H. Cortical neuropathology in aging and dementing disorders. In Cerebral Cortex; Springer: New York, NY, USA, 1999; pp. 175–311. [Google Scholar]
- Buckner, R.L.; Snyder, A.Z.; Shannon, B.J.; LaRossa, G.; Sachs, R.; Fotenos, A.F.; Sheline, Y.I.; Klunk, W.E.; Mathis, C.A.; Morris, J.C.; et al. Molecular, structural, and functional characterization of Alzheimer’s disease: Evidence for a relationship between default activity, amyloid, and memory. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 7709–7717. [Google Scholar] [CrossRef] [PubMed]
- Jack, C.R.; Petersen, R.C.; Xu, Y.C.; Waring, S.C.; O’brien, P.C.; Tangalos, E.G.; Smith, G.E.; Ivnik, R.J.; Kokmen, E. Medial temporal atrophy on MRI in normal aging and very mild Alzheimer’s disease. Neurology 1997, 49, 786–794. [Google Scholar] [CrossRef] [PubMed]
- Killiany, R.J.; Gomez-Isla, T.; Moss, M.; Kikinis, R.; Sandor, T.; Jolesz, F.; Tanzi, R.; Jones, K.; Hyman, B.T.; Albert, M.S. Use of structural magnetic resonance imaging to predict who will get Alzheimer’s disease. Ann. Neurol. 2000, 47, 430–439. [Google Scholar] [CrossRef]
- Baron, J.; Chetelat, G.; Desgranges, B.; Perchey, G.; Landeau, B.; De La Sayette, V.; Eustache, F. In vivo mapping of gray matter loss with voxel-based morphometry in mild Alzheimer’s disease. Neuroimage 2001, 14, 298–309. [Google Scholar] [CrossRef] [PubMed]
- Thompson, P.M.; Mega, M.S.; Woods, R.P.; Zoumalan, C.I.; Lindshield, C.J.; Blanton, R.E.; Moussai, J.; Holmes, C.J.; Cummings, J.L.; Toga, A.W. Cortical change in Alzheimer’s disease detected with a disease-specific population-based brain atlas. Cereb. Cortex 2001, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, B.C.; Bakkour, A.; Salat, D.H.; Feczko, E.; Pacheco, J.; Greve, D.N.; Grodstein, F.; Wright, C.I.; Blacker, D.; Rosas, H.D.; et al. The cortical signature of Alzheimer’s disease: Regionally specific cortical thinning relates to symptom severity in very mild to mild AD dementia and is detectable in asymptomatic amyloid-positive individuals. Cereb. Cortex 2009, 19, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Bozzali, M.; Falini, A.; Franceschi, M.; Cercignani, M.; Zuffi, M.; Scotti, G.; Comi, G.; Filippi, M. White matter damage in Alzheimer’s disease assessed in vivo using diffusion tensor magnetic resonance imaging. J. Neurol. Neurosurg. Psychiatry 2002, 72, 742–746. [Google Scholar]
- Naggara, O.; Oppenheim, C.; Rieu, D.; Raoux, N.; Rodrigo, S.; Dalla Barba, G.; Meder, J. Diffusion tensor imaging in early Alzheimer’s disease. Psychiatry Res. Neuroimaging 2006, 146, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Stahl, R.; Dietrich, O.; Teipel, S.J.; Hampel, H.; Reiser, M.F.; Schoenberg, S.O. White matter damage in Alzheimer disease and mild cognitive impairment: Assessment with diffusion-tensor MR imaging and parallel imaging techniques 1. Radiology 2007, 243, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Friedland, R.P.; Auchus, A.P. Diffusion tensor imaging of normal-appearing white matter in mild cognitive impairment and early Alzheimer’s disease: Preliminary evidence of axonal degeneration in the temporal lobe. AJNR. Am. J. Neuroradiol. 2007, 28, 1943–1948. [Google Scholar] [CrossRef] [PubMed]
- Salat, D.H.; Greve, D.N.; Pacheco, J.L.; Quinn, B.T.; Helmer, K.G.; Buckner, R.L.; Fischl, B. Regional white matter volume differences in nondemented aging and Alzheimer’s disease. Neuroimage 2009, 44, 1247–1258. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Cabronero, J.; Williams, G.B.; Pengas, G.; Nestor, P.J. Absolute diffusivities define the landscape of white matter degeneration in Alzheimer’s disease. Brain 2010, 133 Pt 2, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Douaud, G.; Jbabdi, S.; Behrens, T.E.; Menke, R.A.; Gass, A.; Monsch, A.U.; Rao, A.; Whitcher, B.; Kindlmann, G.; Matthews, P.M.; et al. DTI measures in crossing-fibre areas: Increased diffusion anisotropy reveals early white matter alteration in MCI and mild Alzheimer’s disease. Neuroimage 2011, 55, 880–890. [Google Scholar] [CrossRef] [PubMed]
- Teipel, S.J.; Meindl, T.; Grinberg, L.; Grothe, M.; Cantero, J.L.; Reiser, M.F.; Möller, H.J.; Heinsen, H.; Hampel, H. The cholinergic system in mild cognitive impairment and Alzheimer’s disease: An in vivo MRI and DTI study. Hum. Brain Map. 2011, 32, 1349–1362. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Dougherty, J.H.; Hubner, K.F.; Bai, B.; Cannon, R.L.; Hutson, R.K. Abnormal connectivity in the posterior cingulate and hippocampus in early Alzheimer’s disease and mild cognitive impairment. Alzheimer’s Dement. 2008, 4, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Weiler, M.; de Campos, B.M.; Nogueira, M.H.; Damasceno, B.P.; Cendes, F.; Balthazar, M.L. Structural connectivity of the default mode network and cognition in Alzheimer’s disease. Psychiatry Res. Neuroimaging 2014, 223, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.Y.; Wang, P.N.; Chou, K.H.; Wang, J.; He, Y.; Lin, C.P. Diffusion tensor tractography reveals abnormal topological organization in structural cortical networks in Alzheimer’s disease. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 16876–16885. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Chen, Z.; Evans, A. Structural insights into aberrant topological patterns of large-scale cortical networks in Alzheimer’s disease. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 4756–4766. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Zhang, Y.; Lin, L.; Zhou, Y.; Xu, C.; Jiang, T.; Alzheimer’s Disease Neuroimaging Initiative. Abnormal cortical networks in mild cognitive impairment and Alzheimer’s disease. PLoS Comput. Biol. 2010, 6, e1001006. [Google Scholar] [CrossRef] [PubMed]
- Raj, A.; Kuceyeski, A.; Weiner, M. A network diffusion model of disease progression in dementia. Neuron 2012, 73, 1204–1215. [Google Scholar] [CrossRef] [PubMed]
- Raj, A.; LoCastro, E.; Kuceyeski, A.; Tosun, D.; Relkin, N.; Weiner, M.; Alzheimer’s Disease Neuroimaging Initiative (ADNI). Network diffusion model of progression predicts longitudinal patterns of atrophy and metabolism in Alzheimer’s disease. Cell Rep. 2015, 10, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Shi, F.; Jin, Y.; Yap, P.T.; Wee, C.Y.; Zhang, J.; Yang, C.; Li, X.; Xiao, S.; Shen, D. Multilevel deficiency of white matter connectivity networks in Alzheimer’s disease: A diffusion MRI study with DTI and HARDI models. Neural Plasticity 2016, 2016, 2947136. [Google Scholar] [CrossRef] [PubMed]
- Bastiani, M.; Shah, N.J.; Goebel, R.; Roebroeck, A. Human cortical connectome reconstruction from diffusion weighted MRI: The effect of tractography algorithm. Neuroimage 2012, 62, 1732–1749. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.E.; Tournier, J.; Calamante, F.; Connelly, A. Anatomically-constrained tractography: Improved diffusion MRI streamlines tractography through effective use of anatomical information. Neuroimage 2012, 62, 1924–1938. [Google Scholar] [CrossRef] [PubMed]
- Girard, G.; Whittingstall, K.; Deriche, R.; Descoteaux, M. Towards quantitative connectivity analysis: Reducing tractography biases. Neuroimage 2014, 98, 266–278. [Google Scholar] [CrossRef] [PubMed]
- Gong, G.; Rosa-Neto, P.; Carbonell, F.; Chen, Z.J.; He, Y.; Evans, A.C. Age- and gender-related differences in the cortical anatomical network. J. Neurosci. Off. J. Soc. Neurosci. 2009, 29, 15684–15693. [Google Scholar] [CrossRef] [PubMed]
- Johansen-Berg, H.; Behrens, T.E.J.; Robson, M.D.; Drobnjak, I.; Rushworth, M.F.S.; Brady, J.M.; Smith, S.M.; Higham, D.J.; Matthews, P.M. Changes in connectivity profiles define functionally distinct regions in human medial frontal cortex. Proc. Natl. Acad. Sci. USA 2004, 101, 13335–13340. [Google Scholar] [CrossRef] [PubMed]
- García-Pentón, L.; Fernández, A.P.; Iturria-Medina, Y.; Gillon-Dowens, M.; Carreiras, M. Anatomical connectivity changes in the bilingual brain. Neuroimage 2014, 84, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Shu, N.; An, L.; Wang, P.; Sun, L.; Xia, M.R.; Wang, J.H.; Gong, G.L.; Zang, Y.F.; Wang, Y.F.; et al. Probabilistic diffusion tractography and graph theory analysis reveal abnormal white matter structural connectivity networks in drug-naive boys with attention deficit/hyperactivity disorder. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 10676–10687. [Google Scholar] [CrossRef] [PubMed]
- Alzheimer’s Disease Neuroimaging Initiative (ADNI) Database. Available online: http://adni.loni.usc.edu/ (accessed on 19 August 2014).
- Alzheimer’s Disease Neuroimaging Initiative. Available online: http://www.adni-info.org/ (accessed on 19 August 2014).
- Jenkinson, M.; Bannister, P.; Brady, M.; Smith, S. Improved optimization for the robust and accurate linear registration and motion correction of brain images. Neuroimage 2002, 17, 825–841. [Google Scholar] [CrossRef] [PubMed]
- Jenkinson, M.; Smith, S. A global optimisation method for robust affine registration of brain images. Med. Image Anal. 2001, 5, 143–156. [Google Scholar] [CrossRef]
- Iturria-Medina, Y.; Canales-Rodriguez, E.; Melie-Garcia, L.; Valdes-Hernandez, P.; Martinez-Montes, E.; Alemán-Gómez, Y.; Sánchez-Bornot, J. Characterizing brain anatomical connections using diffusion weighted MRI and graph theory. Neuroimage 2007, 36, 645–660. [Google Scholar] [CrossRef] [PubMed]
- Hagmann, P.; Kurant, M.; Gigandet, X.; Thiran, P.; Wedeen, V.J.; Meuli, R.; Thiran, J. Mapping human whole-brain structural networks with diffusion MRI. PLoS ONE 2007, 2, e597. [Google Scholar] [CrossRef] [PubMed]
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef] [PubMed]
- FreeSurfer. Available online: https://surfer.nmr.mgh.harvard.edu/ (accessed on 15 May 2013).
- Desikan, R.S.; Ségonne, F.; Fischl, B.; Quinn, B.T.; Dickerson, B.C.; Blacker, D.; Buckner, R.L.; Dale, A.M.; Maguire, R.P.; Hyman, B.T.; et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. Neuroimage 2006, 31, 968–980. [Google Scholar] [CrossRef] [PubMed]
- Greve, D.N.; Fischl, B. Accurate and robust brain image alignment using boundary-based registration. Neuroimage 2009, 48, 63–72. [Google Scholar] [CrossRef] [PubMed]
- DSI Studio. Available online: http://dsi-studio.labsolver.org/ (accessed on 24 June 2013).
- Jiang, H.; van Zijl, P.C.; Kim, J.; Pearlson, G.D.; Mori, S. DtiStudio: Resource program for diffusion tensor computation and fiber bundle tracking. Comput. Methods Programs Biomed. 2006, 81, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Watts, D.J.; Strogatz, S.H. Collective dynamics of ‘small-world’networks. Nature 1998, 393, 440–442. [Google Scholar] [CrossRef] [PubMed]
- Bassett, D.S.; Bullmore, E. Small-world brain networks. Neuroscientist 2006, 12, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Latora, V.; Marchiori, M. Efficient behavior of small-world networks. Phys. Rev. Lett. 2001, 87, 198701. [Google Scholar] [CrossRef] [PubMed]
- Latora, V.; Marchiori, M. Economic small-world behavior in weighted networks. Eur. Phys. J. B-Condens. Matter Complex Syst. 2003, 32, 249–263. [Google Scholar] [CrossRef]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Zalesky, A.; Fornito, A.; Bullmore, E.T. Network-based statistic: Identifying differences in brain networks. Neuroimage 2010, 53, 1197–1207. [Google Scholar] [CrossRef] [PubMed]
- Achard, S.; Bullmore, E. Efficiency and cost of economical brain functional networks. PLoS Comput. Biol. 2007, 3, e17. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Chertkow, H.; Lerch, J.P.; Evans, A.C.; Dorr, A.E.; Kabani, N.J. Spatial patterns of cortical thinning in mild cognitive impairment and Alzheimer’s disease. Brain 2006, 129 Pt 11, 2885–2893. [Google Scholar] [CrossRef] [PubMed]
- Im, K.; Lee, J.M.; Seo, S.W.; Yoon, U.; Kim, S.T.; Kim, Y.H.; Kim, S.I.; Na, D.L. Variations in cortical thickness with dementia severity in Alzheimer’s disease. Neurosci. Lett. 2008, 436, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, B.C.; Feczko, E.; Augustinack, J.C.; Pacheco, J.; Morris, J.C.; Fischl, B.; Buckner, R.L. Differential effects of aging and Alzheimer’s disease on medial temporal lobe cortical thickness and surface area. Neurobiol. Aging 2009, 30, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Braak, E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991, 82, 239–259. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Isla, T.; Price, J.L.; McKeel, D.W., Jr.; Morris, J.C.; Growdon, J.H.; Hyman, B.T. Profound loss of layer II entorhinal cortex neurons occurs in very mild Alzheimer’s disease. J. Neurosci. Off. J. Soc. Neurosci. 1996, 16, 4491–4500. [Google Scholar]
- McEvoy, L.K.; Fennema-Notestine, C.; Roddey, J.C.; Hagler, D.J., Jr.; Holland, D.; Karow, D.S.; Pung, C.J.; Brewer, J.B.; Dale, A.M. Alzheimer disease: Quantitative structural neuroimaging for detection and prediction of clinical and structural changes in mild cognitive impairment 1. Radiology 2009, 251, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Galve, L.; Lehmann, M.; Hobbs, N.Z.; Clarkson, M.J.; Ridgway, G.R.; Crutch, S.; Ourselin, S.; Schott, J.M.; Fox, N.C.; Barnes, J. Patterns of cortical thickness according to APOE genotype in Alzheimer’s disease. Dement. Geriatr. Cognit. Disord. 2009, 28, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Fennema-Notestine, C.; Hagler, D.J.; McEvoy, L.K.; Fleisher, A.S.; Wu, E.H.; Karow, D.S.; Dale, A.M. Structural MRI biomarkers for preclinical and mild Alzheimer’s disease. Hum. Brain Map. 2009, 30, 3238–3253. [Google Scholar] [CrossRef] [PubMed]
- Karas, G.; Scheltens, P.; Rombouts, S.; van Schijndel, R.; Klein, M.; Jones, B.; van der Flier, W.; Vrenken, H.; Barkhof, F. Precuneus atrophy in early-onset Alzheimer’s disease: A morphometric structural MRI study. Neuroradiology 2007, 49, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Xuereb, J.H.; Perry, R.H.; Candy, J.M.; Perry, E.K.; Marshall, E.; Bonham, J.R. Nerve cell loss in the thalamus in Alzheimer’s disease and Parkinson’s disease. Brain 1991, 114 Pt 3, 1363–1379. [Google Scholar] [PubMed]
- De Jong, L.W.; Van der Hiele, K.; Veer, I.M.; Houwing, J.J.; Westendorp, R.G.J.; Bollen, E.L.E.M.; De Bruin, P.W.; Middelkoop, H.A.M.; Van Buchem, M.A.; Van Der Grond, J. Strongly reduced volumes of putamen and thalamus in Alzheimer’s disease: An MRI study. Brain 2008, 131, 3277–3285. [Google Scholar] [CrossRef] [PubMed]
- Zarei, M.; Patenaude, B.; Damoiseaux, J.; Morgese, C.; Smith, S.; Matthews, P.M.; Barkhof, F.; Rombouts, S.; Sanz-Arigita, E.; Jenkinson, M. Combining shape and connectivity analysis: An MRI study of thalamic degeneration in Alzheimer’s disease. Neuroimage 2010, 49, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hyman, B.T.; Van Hoesen, G.W.; Damasio, A.R.; Barnes, C.L. Alzheimer’s disease: Cell-specific pathology isolates the hippocampal formation. Science 1984, 225, 1168–1170. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.E.; Hyman, B.T.; Van Hoesen, G.W. Neuropathologic changes of the temporal pole in Alzheimer’s disease and Pick’s disease. Arch. Neurol. 1994, 51, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Karas, G.B.; Burton, E.J.; Rombouts, S.A.R.B.; Van Schijndel, R.A.; O’Brien, J.; Scheltens, P.H.; McKeith, I.G.; Williams, D.; Ballard, C.; Barkhof, F. A comprehensive study of gray matter loss in patients with Alzheimer’s disease using optimized voxel-based morphometry. Neuroimage 2003, 18, 895–907. [Google Scholar] [CrossRef]
- Tuch, D.S. Q-ball imaging. Magn. Resonan. Med. 2004, 52, 1358–1372. [Google Scholar] [CrossRef] [PubMed]
- Amunts, K.; Hawrylycz, M.J.; Van Essen, D.C.; Van Horn, J.D.; Harel, N.; Poline, J.B.; De Martino, F.; Bjaalie, J.G.; Dehaene-Lambertz, G.; Dehaene, S.; et al. Interoperable atlases of the human brain. Neuroimage 2014, 99, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Foundation for the National Institutes of Health. Available online: https://fnih.org/ (accessed on 20 January 2017).
- Acknowledgement List for ADNI Publications. Available online: http://adni.loni.usc.edu/wp-content/uploads/how_to_apply/ADNI_Acknowledgement_List.pdf (accessed on 20 January 2017).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WM-Seed Total Connections | GM-Seed Total Connections | WM-Seed Connections Not in GM-Seed | GM-Seed Connections Not in WM-Seed | Percent of GM-Seed Connections in WM-Seed Networks | |
|---|---|---|---|---|---|
| Control | 1110 (80) | 776 (49) | 380 (59) | 46 (21) | 94.1% (2.5%) |
| AD | 1089 (74) | 723 (64) | 411 (51) | 45 (17) | 93.7% (2.5%) |
| WM-Seed Network Density | GM-Seed Network Density | WM-Seed Average Network Strength | GM-Seed Average Network Strength | |
|---|---|---|---|---|
| Control | 0.195 (0.0141) * | 0.136 (0.00858) | 1361.5 (307.9) * | 572.4 (117.6) |
| AD | 0.191 (0.0131) * | 0.127 (0.0112) | 1251.6 (197.3) * | 579.9 (99.2) |
| Node | Number of Differential Connections |
|---|---|
| Controls | |
| Left precentral | 6 |
| Left superior parietal | 7 |
| Left insula | 8 |
| Right inferior parietal | 6 |
| Right lateral occipital | 6 |
| Right superior parietal | 7 |
| Right insula | 11 |
| AD | |
| Left superior parietal | 7 |
| Right superior parietal | 7 |
| Right insula | 6 |
| Seed ROI | FA-Thresholded Seed ROI Volume (Voxel Number) | Number of Streamlines Generated between Seed ROI and GM Target ROI |
|---|---|---|
| Subject 1 | ||
| WM left inferior parietal lobe | 304 | 904 |
| GM left inferior parietal lobe | 269 | 264 |
| WM left entorhinal cortex | 20 | 1248 |
| GM left entorhinal cortex | 48 | 45 |
| WM right rostral anterior cingulate | 55 | 112 |
| GM right rostral anterior cingulate | 68 | 9 |
| Subject 2 | ||
| WM left inferior parietal lobe | 275 | 99 |
| GM left inferior parietal lobe | 276 | 66 |
| WM left entorhinal cortex | 22 | 232 |
| GM left entorhinal cortex | 32 | 456 |
| WM right rostral anterior cingulate | 63 | 61 |
| GM right rostral anterior cingulate | 82 | 6 |
| Network | Weighted Global Efficiency | Weighted Global Efficiency (Random) | Weighted Local Efficiency | Weighted Local Efficiency (Random) |
|---|---|---|---|---|
| Control WM-seed | 72.8 (20.3) | 84.7 (20.3) | 52.8 (11.3) | 30.6 (7.78) |
| Control GM-seed | 29.2 (7.47) | 36.5 (7.89) | 26.2 (4.52) | 11.5 (3.01) |
| AD WM-seed | 62.6 (10.4) | 77.5 (12.6) | 46.8 (8.71) | 27.3 (4.71) |
| AD GM-seed | 27.9 (5.23) | 36.5 (6.54) | 26.8 (5.38) | 10.5 (2.36) |
| WM-Seed Network Node | Degree in Significant NBS Network |
|---|---|
| Left entorhinal cortex | 4 |
| Left isthmus cingulate | 5 |
| Left thalamus | 5 |
| Right precuneus | 4 |
| Right superior parietal cortex | 5 |
| GM-Seed Network Node | Degree in Significant NBS Network |
| Left precentral gyrus | 2 |
| Left rostral middle frontal gyrus | 2 |
| Left temporal pole | 2 |
| Left hippocampus | 2 |
| Left thalamus | 5 |
| Node | Control WM-Seed Nodal Efficiency | AD WM-Seed Nodal Efficiency | Control GM-Seed Nodal Efficiency | AD GM-Seed Nodal Efficiency |
|---|---|---|---|---|
| Left entorhinal | 108 (44.3) ** | 63.2 (27.5) | 51.2 (26.5) | 35.2 (14.8) |
| Left isthmus cingulate | 81.6 (32.5) * | 62.3 (16.1) | 28.8 (10.5) | 29.5 (9.44) |
| Left thalamus | 89.9 (33.4) * | 66.1 (23.1) | 52.2 (20.3) ** | 31.1 (11.1) |
| Right precuneus | 97.1 (41) | 74.1 (29.6) | 28.4 (11.5) | 25.9 (9.15) |
| Right superior parietal cortex | 103 (35.7) | 88.6 (30.4) | 32.2 (10.4) | 31.8 (8.8) |
| Left precentral gyrus | 106 (30.4) * | 80.9 (20.3) | 49.4 (17.5) | 37.1 (11) |
| Left rostral middle frontal gyrus | 66.7 (14.4) ** | 51.7 (9.46) | 28.2 (5.44) | 25.6 (7.48) |
| Left temporal pole | 95.6 (35) * | 73.6 (25.1) | 52.9 (16.3) | 44.1 (14.2) |
| Left hippocampus | 95 (25.5) ** | 67.1 (25.2) | 40.2 (14.4) | 31.3 (13.4) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zajac, L.; Koo, B.-B.; Bauer, C.M.; Killiany, R.; Behalf of the Alzheimer’s Disease Neuroimaging Initiative. Seed Location Impacts Whole-Brain Structural Network Comparisons between Healthy Elderly and Individuals with Alzheimer’s Disease. Brain Sci. 2017, 7, 37. https://doi.org/10.3390/brainsci7040037
Zajac L, Koo B-B, Bauer CM, Killiany R, Behalf of the Alzheimer’s Disease Neuroimaging Initiative. Seed Location Impacts Whole-Brain Structural Network Comparisons between Healthy Elderly and Individuals with Alzheimer’s Disease. Brain Sciences. 2017; 7(4):37. https://doi.org/10.3390/brainsci7040037
Chicago/Turabian StyleZajac, Lauren, Bang-Bon Koo, Corinna M. Bauer, Ron Killiany, and Behalf of the Alzheimer’s Disease Neuroimaging Initiative. 2017. "Seed Location Impacts Whole-Brain Structural Network Comparisons between Healthy Elderly and Individuals with Alzheimer’s Disease" Brain Sciences 7, no. 4: 37. https://doi.org/10.3390/brainsci7040037
APA StyleZajac, L., Koo, B. -B., Bauer, C. M., Killiany, R., & Behalf of the Alzheimer’s Disease Neuroimaging Initiative. (2017). Seed Location Impacts Whole-Brain Structural Network Comparisons between Healthy Elderly and Individuals with Alzheimer’s Disease. Brain Sciences, 7(4), 37. https://doi.org/10.3390/brainsci7040037