Time-Frequency Analysis of Mu Rhythm Activity during Picture and Video Action Naming Tasks


Abstract
:1. Introduction
1.1. Mirror Activity in Response to Action Verbs
1.2. Stimuli Effects
1.3. Neurophysiological Measures
1.4. Mu Rhythm Activity
2. Methods
2.1. Participants
2.2. Stimuli
2.3. Data Collection
2.4. Data Analysis
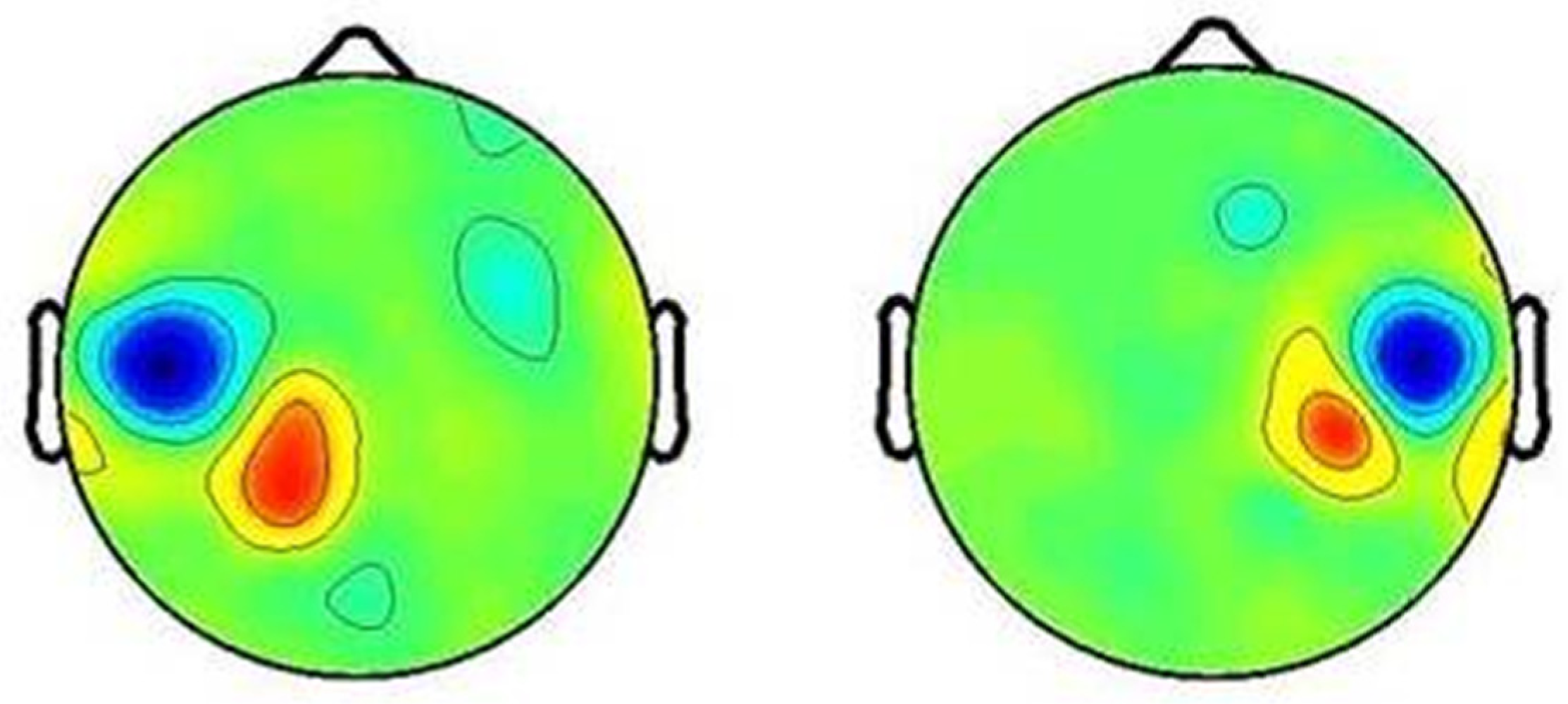
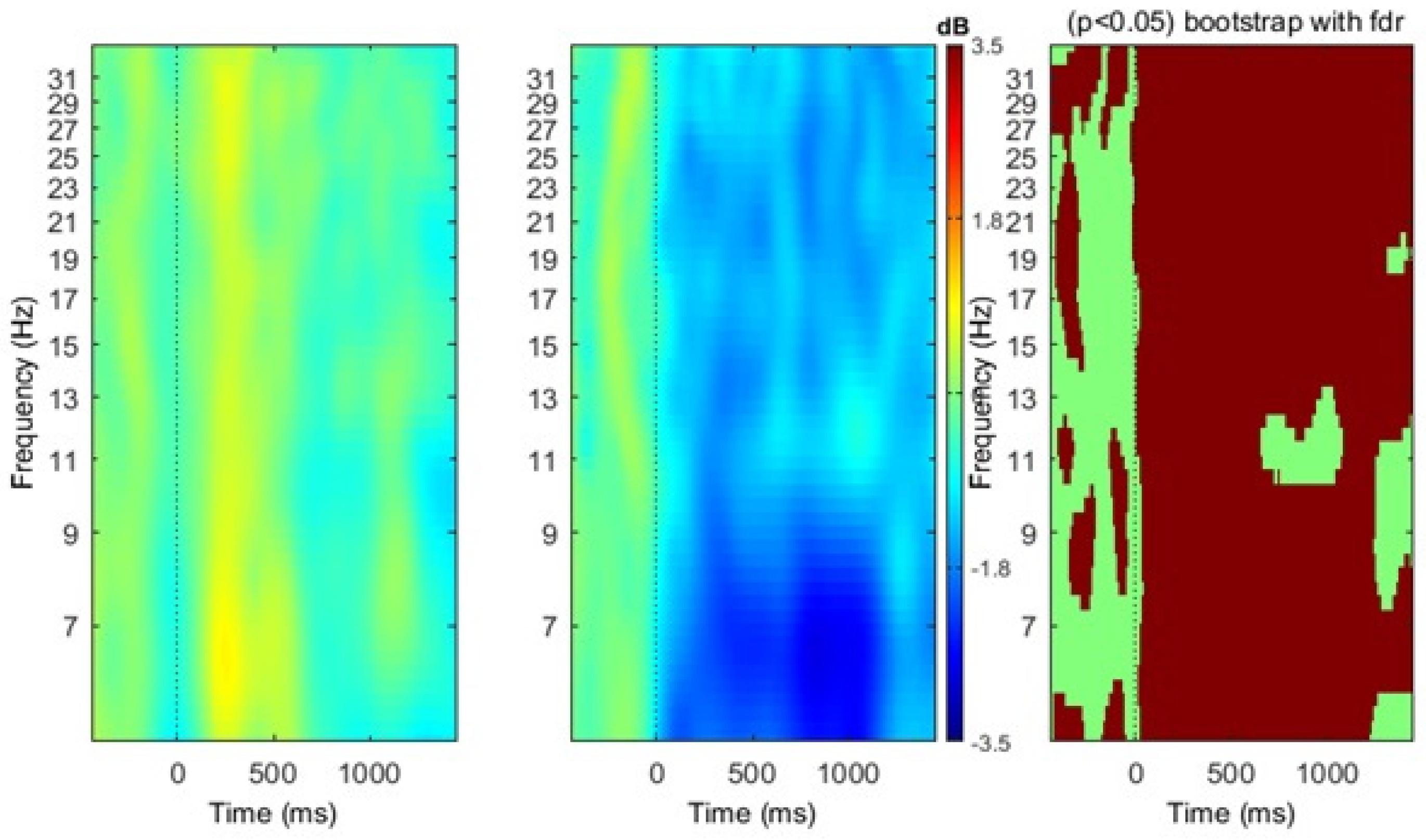
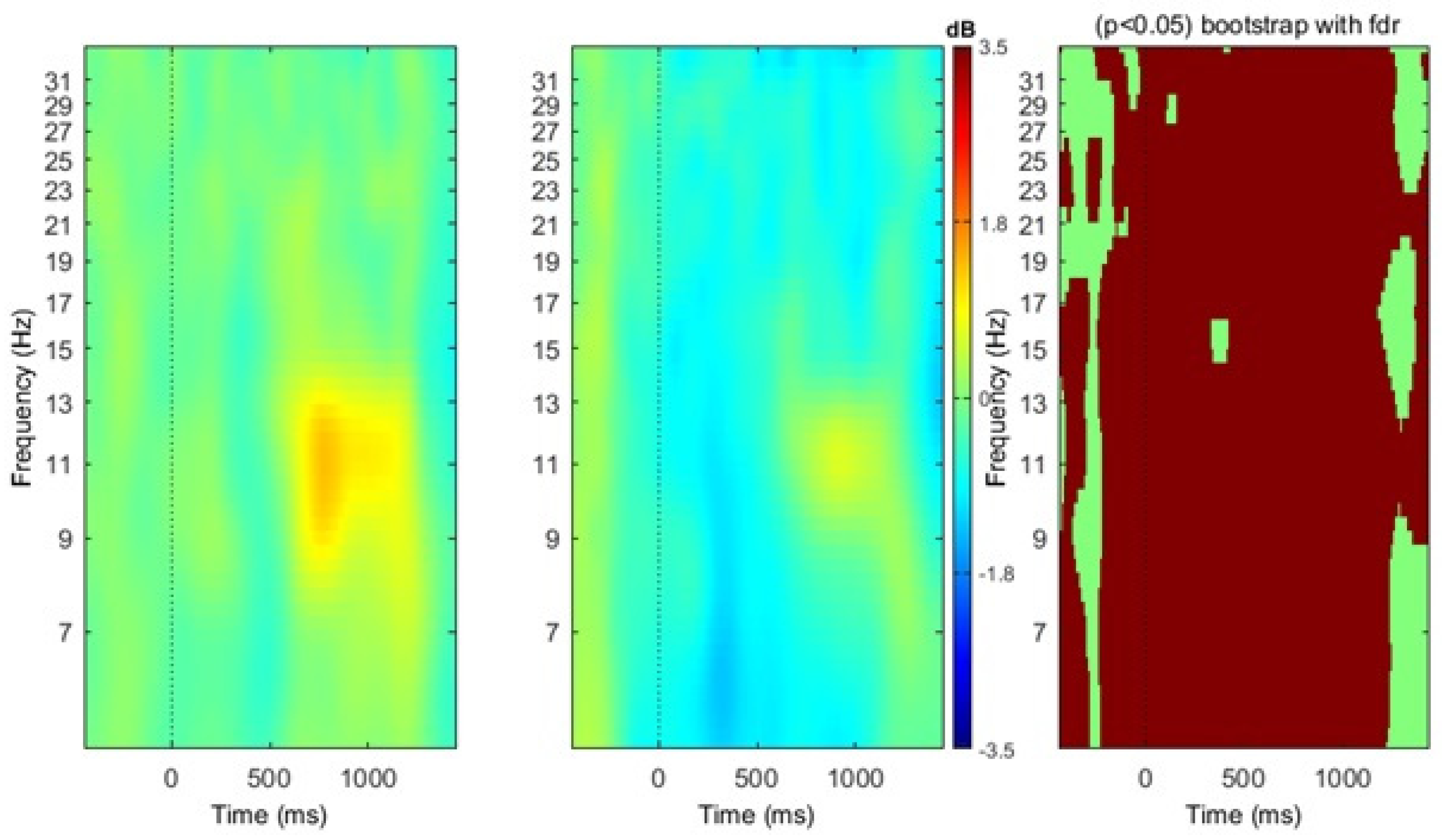
3. Results
4. Discussion
4.1. Localization of Mu Rhythm ERD during Action Verb Naming
4.2. Action Observation and Action Semantics
4.3. Limitations and Future Directions
5. Conclusions
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | FK Conc. | BN Conc. | Familiarity | Imageability | FK Freq. | BN Freq. |
|---|---|---|---|---|---|---|
| Typing | 376 | 567 | 395 | 3.24 | ||
| Pouring | 356 | 3.8 | 545 | 495 | 5.92 | 302 |
| Stapling | 3.61 | 0.18 | 9 | |||
| Cutting | 430 | 4.11 | 581 | 460 | 22.08 | 1126 |
| Folding | 3.89 | 2.08 | 106 | |||
| Throwing | 400 | 3.69 | 548 | 477 | 29.18 | 1488 |
| Painting | 577 | 4.59 | 551 | 567 | 29.20 | 1489 |
| Stirring | 2.93 | 2.61 | 133 | |||
| Catching | 3.42 | 12.71 | 648 | |||
| Hammering | 605 | 3.85 | 515 | 618 | 1.57 | 80 |
| Ironing | 584 | 4.1 | 555 | 561 | 1.47 | 75 |
| Mixing | 3.45 | |||||
| Mowing | 3.63 | 0.94 | 48 | |||
| Rolling | 457 | 4.04 | 547 | 496 | 21.55 | 1099 |
| Shaving | 4.63 | 5.84 | 298 | |||
| Stacking | 0.90 | |||||
| Stamping | 3.66 | 0.55 | 28 | |||
| Opening | 381 | 3.79 | 581 | 425 | 38.43 | 1960 |
| Peeling | 432 | 3.82 | 507 | 436 | 1.10 | 56 |
| Knitting | 583 | 4.14 | 501 | 578 | 2.25 | 115 |
References
- Rizzolatti, G.; Craighero, L. The mirror-neuron system. Ann. Rev. Neurosci. 2004, 27, 169–192. [Google Scholar] [CrossRef] [PubMed]
- Rizzolatti, G.; Fabbri-Destro, M. Mirror neurons and mirror systems in monkeys and humans. Am. Physiol. Soc. 2008, 23, 171–179. [Google Scholar]
- Rizzolatti, G.; Fadiga, L.; Gallese, V.; Fogassi, L. Premotor cortex and the recognition of motor actions. Cogn. Brain Res. 1996, 3, 131–141. [Google Scholar] [CrossRef]
- Buccino, G.; Binkofski, F.; Fink, G.R.; Fadiga, L.; Gallese, V.; Seitz, R.J.; Zilles, K.; Rizzolatti, G.; Freund, H.J. Action observation activates premotor and parietal areas in a somatotopic manner: An fMRI study. Eur. J. Neurosci. 2001, 13, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Chong, T.T.J.; Cunnington, R.; Williams, M.A.; Kanwisher, N.; Mattingley, J.B. fMRI adaptation reveals mirror neurons in human inferior parietal cortex. Curr. Biol. 2008, 18, 1576–1580. [Google Scholar] [CrossRef] [PubMed]
- Molengerphs, P.; Cunnington, R.; Mattingley, J.B. Brain regions with mirror properties: A meta-analysis of 125 human fMRI studies. Neurosci. Biobehav. Rev. 2012, 36, 341–349. [Google Scholar] [Green Version]
- Rozzi, S.; Ferrari, P.F.; Bonini, L.; Rizzolatti, G.; Fogassi, L. Functional organization of inferior parietal lobule convexity in the macaque monkey: Electrophysiological characterization of motor, sensory and mirror responses and their correlation with cytoarchitectonic areas. Eur. J. Neurosci. 2008, 28, 1569–1588. [Google Scholar] [CrossRef] [PubMed]
- Fadiga, L.; Fogassi, L.; Pavesi, G.; Rizzolatti, G. Motor facilitation during action observation: A magnetic stimulation study. J. Neurophysiol. 1995, 73, 2608–2611. [Google Scholar] [PubMed]
- Hari, R.; Forss, N.; Avikainen, S.; Kirveskari, E.; Salenius, S.; Rizzolatti, G. Activation of human primary motor cortex during action observation: A neuromagnetic study. Proc. Natl. Acad. Sci. USA 1998, 95, 15061–15065. [Google Scholar] [CrossRef] [PubMed]
- Enticott, P.G.; Johnston, P.J.; Herring, S.E.; Hoy, K.E.; Fitzgerald, P.B. Mirror neuron activation is associated with facial emotion processing. Neuropsychologia 2008, 46, 2851–2854. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.; Gorodnitsky, I.; Pineda, J. EEG mu component responses to viewing emotional faces. Behav. Brain Res. 2012, 226, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Ogura, M.; Watanabe, Y.; Sanjo, Y.; Edahiro, A.; Sato, K.; Katakura, A. Mirror neurons activated during swallowing and finger movements: An fMRI study. J. Oral Maxillofac. Surg. Med. Pathol. 2014, 26, 188–197. [Google Scholar] [CrossRef]
- Ushioda, T.; Watanabe, Y.; Sanjo, Y.; Yamane, G.Y.; Abe, S.; Tsuji, Y.; Ishiyama, A. Visual and auditory stimuli associated with swallowing activate mirror neurons: A magnetoencephalography study. Dysphagia 2012, 27, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Restle, J.; Ziemann, U. Observation-execution matching and action inhibition in human primary motor cortex during viewing of speech-related lip movements or listening to speech. Neuropsychologia 2011, 49, 2045–2054. [Google Scholar] [CrossRef] [PubMed]
- Boulenger, V.; Hauk, O.; Pulvermuller, F. Grasping ideas with the motor system: Semantic somatotopy in idiom comprehension. Cereb. Cortex 2009, 19, 1905–1914. [Google Scholar] [CrossRef] [PubMed]
- Aziz-Zadeh, L.; Wilson, S.M.; Rizzolatti, G.; Iacoboni, M. Congruent embodied representations for visually presented actions and linguistic phrases describing actions. Curr. Biol. 2006, 16, 1818–1823. [Google Scholar] [CrossRef] [PubMed]
- Pulvermuller, F.; Harle, M.; Hummel, F. Walking or talking? Behavioral and neurophysiological correlates of action verb processing. Brain Lang. 2001, 78, 143–168. [Google Scholar] [CrossRef] [PubMed]
- Andric, M.; Solodkin, A.; Buccino, G.; Goldin-Meadow, S.; Rizzolatti, G.; Small, S.L. Brain function overlaps when people observe emblems, speech, and grasping. Neuropsychologia 2013, 51, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Willems, R.M.; Hagoort, P.; Casasanto, D. Body-specific representations of action verbs neural evidence from right- and left-handers. Psychol. Sci. 2010, 21, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Perry, A.; Bentin, S. Mirror activity in the human brain while observing hand movements: A comparison between EEG desynchronization in the μ-range and previous fMRI results. Brain Res. 2009, 1282, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Marcus, N.; Cleary, B.; Wong, A.; Ayres, P. Should hand actions be observed when learning hand motor skills from instructional animations? Comput. Hum. Behav. 2013, 29, 2172–2178. [Google Scholar] [CrossRef]
- Ulloa, E.R.; Pineda, J.A. Recognition of point-light biological motion: Mu rhythms and mirror neuron activity. Behav. Brain Res. 2007, 183, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Van Overwalle, F.; Baetens, K. Understanding others’ actions and goals by mirror and mentalizing systems: A meta-analysis. Neuroimage 2009, 48, 564–584. [Google Scholar] [CrossRef] [PubMed]
- Gangitano, M.; Mottaghy, F.M.; Pascual-Leone, A. Phase-specific modulation of cortical motor output during movement observation. Neuroreport 2001, 12, 1489–1492. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, K.J.; Isenberg, N.; Haxby, J.V. Communicative hand gestures and object-directed hand movements activated the mirror neuron system. Soc. Cogn. Affect. Neurosci. 2007, 2, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Muthukumaraswamy, S.D.; Johnson, B.W.; McNair, N.A. Mu rhythm modulation during observation of an object-directed grasp. Cogn. Brain Res. 2004, 19, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Buccino, G.; Lui, F.; Canessa, N.; Patteri, I.; Lagravinese, G.; Benuzzi, F.; Porro, C.; Rizzolatti, G. Neural circuits involved in the recognition of actions performed by nonconspecifics: An fMRI study. J. Cogn. Neurosci. 2004, 16, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Umilta, M.A.; Kohler, E.; Gallese, V.; Fogassi, L.; Fadiga, L.; Keysers, C.; Rizzolatti, G. I know what you are doing: A neurophysiological study. Neuron 2001, 31, 155–165. [Google Scholar] [CrossRef]
- Den Ouden, D.B.; Fix, S.; Parrish, T.B.; Thompson, C.K. Argument structure effects in action verb naming in static and dynamic conditions. J. Neurolinguist. 2009, 22, 196–215. [Google Scholar] [CrossRef] [PubMed]
- Hauk, O.; Pulvermuller, F. Neurophysiological distinction of action words in the fronto-central cortex. Hum. Brain Mapp. 2004, 21, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Indefrey, P.; Levelt, W.J.M. The spatial and temporal signatures of word production components. Cognition 2004, 92, 101–144. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Bin, G.; Daly, I.; Gao, X. Event-related desynchronization (ERD) in the alpha band during hand mental rotation task. Neurosci. Lett. 2013, 541, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Hari, R. Action–perception connection and the cortical mu rhythm. Prog. Brain Res. 2006, 159, 253–260. [Google Scholar] [PubMed]
- Jensen, O.; Goel, P.; Kopell, N.; Pohja, M.; Hari, R.; Ermentrout, B. On the human sensorimotor-cortex beta rhythm: Sources and modeling. Neuroimage 2005, 26, 347–355. [Google Scholar] [CrossRef] [PubMed]
- McGarry, L.M.; Russo, F.A.; Schalles, M.D.; Pineda, J.A. Audio-visual facilitation of the mu rhythm. Exp. Brain Res. 2012, 218, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Pineda, J.A. The functional significance of my rhythms: Translating “seeing” and “hearing” into “doing”. Brain Res. Rev. 2005, 50, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Pineda, J.A. Sensorimotor cortex as a critical component of an ‘extended’ mirror neuron system: Does it solve development, correspondence, and control problems in mirroring? Behav. Brain Funct. 2008, 4. [Google Scholar] [CrossRef] [PubMed]
- Neuper, C.; Pfurtscheller, G. Event-related dynamics of cortical rhythms: Frequency-specific features and functional correlates. Int. J. Psychophysiol. 2001, 43, 41–58. [Google Scholar] [CrossRef]
- Pfurtscheller, G.; Lopes da Silva, F.H. Event-related EEG/MEG synchronization and desynchronization: Basic principles. Clin. Neurophysiol. 1999, 110, 1842–1857. [Google Scholar] [CrossRef]
- Onton, J.; Westerfield, M.; Townsend, J.; Makeig, S. Imaging human EEG dynamics using independent component analysis. Neurosci. Biobehav. Rev. 2006, 30, 808–822. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, P.N.; Shanamugan, D.; Chourasia, A.; Ghole, A.R.; Acharyya, A.A.; Naik, G. Automated detection and correction of eye blink and muscular artefacts in EEG signal for analysis of Autism Spectrum Disorder. In Proceedings of the 2014 36th Annual International Conference of the IEEE on Engineering in Medicine and Biology Society (EMBC), Chicago, IL, USA, 26–30 August 2014; pp. 1881–1884. [Google Scholar]
- Bhardwaj, S.; Jadhav, P.; Adapa, B.; Acharyya, A.; Naik, G.R. Online and automated reliable system design to remove blink and muscle artefact in EEG. In Proceedings of the 2015 37th Annual International Conference of the IEEE on Engineering in Medicine and Biology Society (EMBC), Milan, Italy, 25–29 August 2015; pp. 6784–6787. [Google Scholar]
- Mammone, N.; Morabito, F.C. Independent component analysis and high-order statistics for automatic artifact rejection. In Proceedings of the IEEE International Joint Conference on Neural Networks, Montreal, QC, Canada, 31 July–4 August 2005; Volume 4, pp. 2447–2452. [Google Scholar]
- Greco, A.; Mammone, N.; Morabito, F.C.; Versaci, M. Kurtosis, Renyi’s entropy and independent component scalp maps for the automatic artifact rejection from EEG data. Int. J. Signal Process. 2006, 2, 240–244. [Google Scholar]
- Bowers, A.; Saltuklaroglu, T.; Harkrider, A.; Cuellar, M. Suppression of the µ rhythm during speech and non-speech discrimination revealed by independent component analysis: Implications for sensorimotor integration in speech processing. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Chuang, C.H.; Ko, L.W.; Jung, T.P.; Lin, C.T. Kinesthesia in a sustained-attention driving task. Neuroimage 2014, 91, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Makeig, S.; Delorme, A.; Westerfield, M.; Jung, T.P.; Townsend, J.; Courchesne, E.; Sejnowski, T.J. Electroencephalographic brain dynamics following manually responded visual targets. PLoS Biol. 2004, 2, 747–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naeem, M.; Brunner, C.; Leeb, R.; Graimann, B.; Pfurtscheller, G. Seperability of four-class motor imagery data using independent components analysis. J. Neural Eng. 2006, 3, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Tsai, A.C.; Jung, T.P.; Chien, V.S.C.; Savostyanov, A.N.; Makeig, S. Cortical surface alignment in multi-subject spatiotemporal independent EEG source imaging. Neuroimage 2014, 87, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Onton, J.; Makeig, S. Information-based modeling of event-related brain dynamics. Prog. Brain Res. 2006, 159, 99–120. [Google Scholar] [PubMed]
- Makeig, S.; Debener, S.; Onton, J.; Delorme, A. Mining event-related brain dynamics. Trends Cogn. Sci. 2004, 8, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Moreno, I.; De Vega, M.; León, I. Understanding action language modulates oscillatory mu and beta rhythms in the same way as observing actions. Brain Cogn. 2013, 82, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Moreno, I.; De Vega, M.; León, I.; Bastiaansen, M.; Lewis, A.G.; Magyari, L. Brain dynamics in the comprehension of action-related language. A time-frequency analysis of mu rhythms. Neuroimage 2015, 109, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Vukovic, N.; Shtyrov, Y. Cortical motor systems are involved in second-language comprehension: Evidence from rapid mu-rhythm desynchronisation. Neuroimage 2014, 102, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Goodglass, H.; Kaplan, E.; Weintraub, S.; Segal, O. Boston Naming Test; Lippincott Willams Wilkins: Philadelphia, PA, USA, 2001. [Google Scholar]
- Nasreddine, Z.S.; Phillips, N.A.; Bedirian, V.; Charbonneau, S.; Whitehead, V.; Collin, I.; Cummings, J.L.; Chertkow, H. The montreal cognitive assessment, MoCA: A brief screening tool for mild cognitive impairment. J. Am. Geriatr. Soc. 2005, 53, 695–699. [Google Scholar] [CrossRef] [PubMed]
- Umla-Runge, K.; Zimmer, H.D.; Fu, X.; Wang, L. An action video clip database rated for familiarity in China and Germany. Behav. Res. Methods 2012, 44, 946–953. [Google Scholar] [CrossRef] [PubMed]
- iMovie, Version 10.0; video editing software; Apple Inc.: Cupertino, CA, USA, 2015.
- Coltheart, M. The MRC Psycholinguistic Database. Q. J. Exp. Psychol. 1981, 33A, 497–505. [Google Scholar] [CrossRef]
- Francis, W.N.; Kucera, H. Computational Analysis of Present-Day American English; Brown University Press: Providence, RI, USA, 1967. [Google Scholar]
- Brysbaert, M.; New, B. Moving beyond Kucera and Francis: A critical evaluation of current word frequency norms and the introduction of a new and improved word frequency measure for American English. Behav. Res. Methods 2009, 41, 977–990. [Google Scholar] [CrossRef] [PubMed]
- Brysbaert, M.; Warriner, A.B.; Kuperman, V. Concreteness ratings for 40 thousand generally known English word lemmas. Behav. Res. Methods 2014, 46, 904–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Psychology Software Tools, Inc. [E-Prime 2.0]. Available online: http://www.pstnet.com (accessed on 13 October 2013).
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Makeig, S.; Onton, J. ERP features and EEG dynamics: An ICA perspective. In The Oxford Handbook of Event-Related Potential Components; Luck, S.J., Kappenman, E.S., Eds.; Oxford University Press: New York, NY, USA, 2011; pp. 51–86. [Google Scholar]
- Manly, J.J. Critical issues in cultural neuropsychology: Profit from diversity. Neuropsychol. Rev. 2008, 18, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Singh, K. Confidence distribution, the frequentist distribution estimator of a parameter: A review. Int. Stat. Rev. 2013, 81, 3–39. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. Royal Stat. Soc. Series B (Methodol.) 1995, 57, 289–300. [Google Scholar]
- Pulvermuller, F. Brain mechanisms linking language and action. Nat. Rev. Neurosci. 2005, 6, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Rose, M.L.; Raymer, A.M.; Lanyon, L.E.; Attard, M.C. A systematic review of gesture treatments for post-stroke aphasia. Aphasiology 2013, 27, 1090–1127. [Google Scholar] [CrossRef]
- Marangolo, P.; Caltagirone, C. Options to enhance recovery from aphasia by means of non-invasive brain stimulation and action observation therapy. Exp. Rev. Neurother. 2014, 14, 75–91. [Google Scholar] [CrossRef] [PubMed]
- Marangolo, P.; Cipollari, S.; Fiori, V.; Razzano, C.; Caltagirone, C. Walking but not barking improves verb recovery: Implication for action observation treatment in aphasia rehabilitation. PLoS ONE 2010, 7. [Google Scholar] [CrossRef] [PubMed]
- Marangolo, P.; Bonifazi, S.; Tomaiuolo, F.; Craighero, L.; Coccia, M.; Altoe, G.; Provinciali, L.; Cantagallo, A. Improving language without words: First evidence from aphasia. Neuropsychologia 2010, 48, 3824–3833. [Google Scholar] [CrossRef] [PubMed]
- Bonifazi, S.; Tomaiuolo, F.; Altoe, G.; Ceravolo, M.G.; Provinciali, L.; Marangolo, P. Action observation as a useful approach for enhancing recovery of verb production: New evidence from aphasia. Eur. J. Phys. Rehabil. Med. 2013, 49, 473–481. [Google Scholar] [PubMed]
- Faroqi-Shah, Y.; Graham, L. Semantic treatment of verb naming in aphasia: Acquisition and generalization effects. Clin. Ling. Phon. 2011, 25, 399–418. [Google Scholar] [CrossRef] [PubMed]
- Conroy, P.; Sage, K.; Lambon Ralph, M.A. The effects of decreasing and increasing cue therapy on improving naming speed and accuracy for verbs and nouns in aphasia. Aphasiology 2009, 23, 707–730. [Google Scholar] [CrossRef]
- Fridriksson, J.; Morrow, K.L.; Moser, D.; Baylis, G.C. Age-related variability in cortical activity during language processing. J. Speech Lang. Hear. Res. 2006, 49, 690–697. [Google Scholar] [CrossRef]




| Age | Education | BNT | MOCA | Handedness | Monolingual | |
|---|---|---|---|---|---|---|
| Average | 36.1 | 17.4 | 15 | 28.8 | 19 Right | 17 Yes |
| Standard Deviation | 16.1 | 1.6 | 1 | 1.2 | 2 Left | 4 No |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuellar, M.E.; Del Toro, C.M. Time-Frequency Analysis of Mu Rhythm Activity during Picture and Video Action Naming Tasks. Brain Sci. 2017, 7, 114. https://doi.org/10.3390/brainsci7090114
Cuellar ME, Del Toro CM. Time-Frequency Analysis of Mu Rhythm Activity during Picture and Video Action Naming Tasks. Brain Sciences. 2017; 7(9):114. https://doi.org/10.3390/brainsci7090114
Chicago/Turabian StyleCuellar, Megan E., and Christina M. Del Toro. 2017. "Time-Frequency Analysis of Mu Rhythm Activity during Picture and Video Action Naming Tasks" Brain Sciences 7, no. 9: 114. https://doi.org/10.3390/brainsci7090114
APA StyleCuellar, M. E., & Del Toro, C. M. (2017). Time-Frequency Analysis of Mu Rhythm Activity during Picture and Video Action Naming Tasks. Brain Sciences, 7(9), 114. https://doi.org/10.3390/brainsci7090114