Endocannabinoids and Fear-Related Behavior in Mice Selectively Bred for High or Low Alcohol Preference



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Drugs
2.3. Acoustic Startle Apparatus
2.4. FPS Procedures
2.5. Brain Tissue Preparation and Analysis
2.6. Estrous Cycle Monitoring
2.7. Study Procedures
2.7.1. Experiment 1: FPS and EC Brain Levels in Male and Female HAP1 and LAP1 Mice
2.7.2. Experiment 2: Effects of CP55940, rimonabant, alcohol, and diazepam on the expression of FPS in male and female HAP1 and LAP1 mice
2.8. Statistical Analyses
3. Results
3.1. Experiment 1: FPS and AEA and 2-AG Brain Levels in Male and Female HAP1 and LAP1 Mice
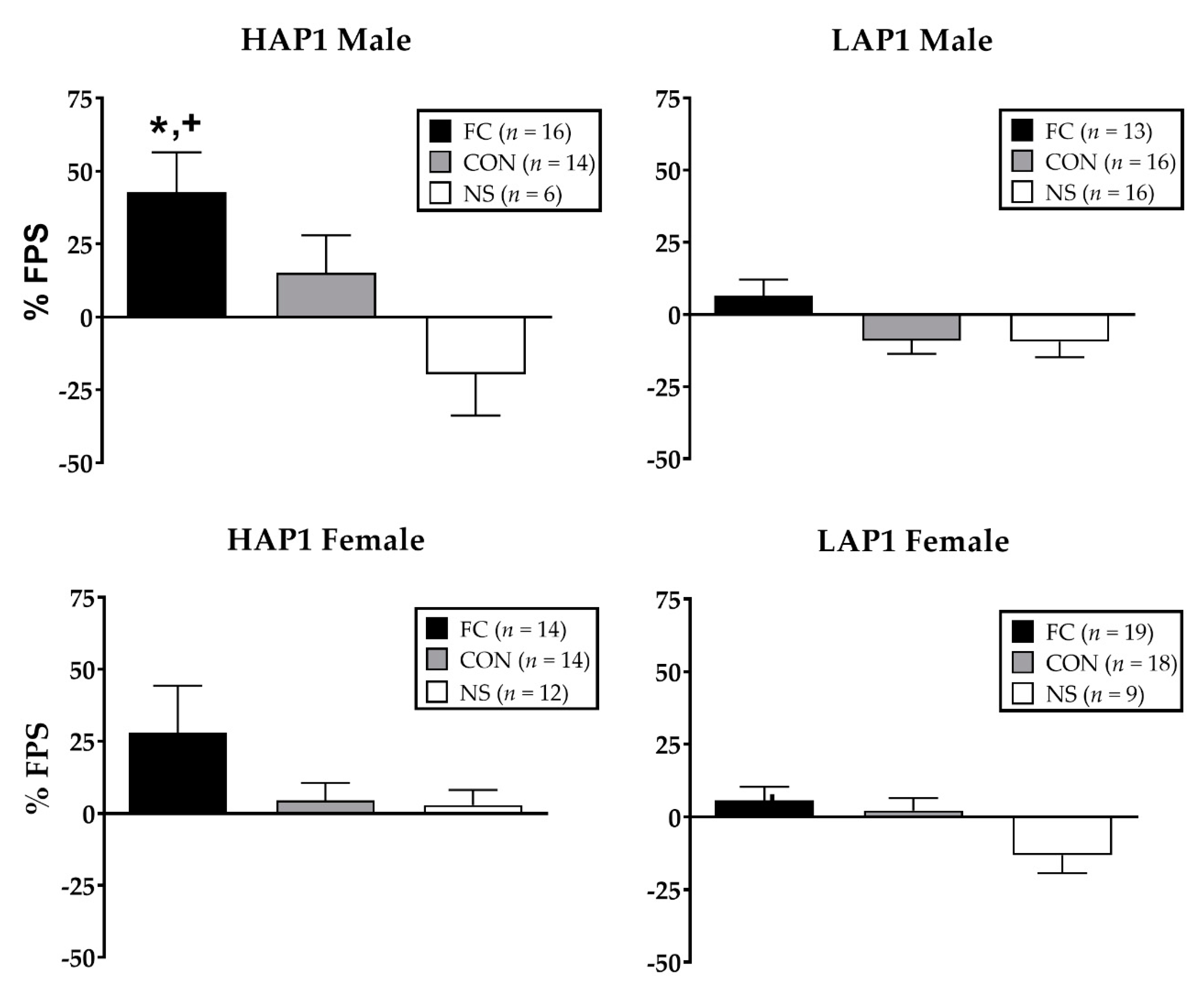
3.1.1. % FPS
3.1.2. AEA and 2-AG in PFC, AMG, and HIP Following FPS Testing
AEA
2-AG
3.2. Experiment 2: Effects of CP55940, Rimonabant, Alcohol, and Diazepam on the Expression of FPS in Male and Female HAP1 and LAP1 Mice % FPS
3.2.1. CP55940
3.2.2. Rimonabant
3.2.3. Alcohol and Diazepam
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Allen, J.P.; Crawford, E.F.; Kudler, H. Nature and treatment of comorbid alcohol problems and post traumatic stress disorder among American military personnel and veterans. Alcohol Res. 2016, 38, 133–140. [Google Scholar] [PubMed]
- Grant, B.F.; Saha, T.D.; Ruan, W.J.; Goldstein, R.B.; Chou, S.P.; Jung, J.; Zhang, H.; Smith, S.M.; Pickering, R.P.; Huang, B.; et al. Epidemiology of DSM-5 Drug Use Disorder: Results From the National Epidemiologic Survey on Alcohol and Related Conditions-III. JAMA Psychiatry 2016, 73, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Blanco, C.; Xu, Y.; Brady, K.; Perez-Fuentes, G.; Okuda, M.; Wang, S. Comorbidity of posttraumatic stress disorder with alcohol dependence among US adults: Results from National Epidemiological Survey on Alcohol and Related Conditions. Drug Alcohol Depend. 2013, 132, 630–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartor, C.E.; McCutcheon, V.V.; Pommer, N.E.; Nelson, E.C.; Grant, J.D.; Duncan, A.E.; Waldron, M.; Bucholz, K.K.; Madden, P.A.; Heath, A.C. Common genetic and environmental contributions to post-traumatic stress disorder and alcohol dependence in young women. Psychol. Med. 2011, 41, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Enoch, M.A. The influence of gene-environment interactions on the development of alcoholism and drug dependence. Curr. Psychiatry Rep. 2012, 14, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Greenwald, M.K. Anti-stress neuropharmacological mechanisms and targets for addiction treatment: A translational framework. Neurobiol. Stress 2018, 9, 84–104. [Google Scholar] [CrossRef] [PubMed]
- Ney, L.J.; Matthews, A.; Bruno, R.; Felmingham, K.L. Cannabinoid interventions for PTSD: Where to next? Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 93, 124–140. [Google Scholar] [CrossRef] [PubMed]
- Spagnolo, P.A.; Ramchandani, V.A.; Schwandt, M.L.; Kwako, L.E.; George, D.T.; Mayo, L.M.; Hillard, C.J.; Heilig, M. FAAH gene variation moderates stress response and symptom severity in patients with posttraumatic stress disorder and comorbid alcohol dependence. Alcohol Clin. Exp. Res. 2016, 40, 2426–2434. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Kreek, M.J. Involvement of activated brain stress responsive systems in excessive and “relapse” alcohol drinking in rodent models: Implications for therapeutics. J. Pharmacol. Exp. Ther. 2018, 366, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Devane, W.A.; Dysarz, F.A., 3rd; Johnson, M.R.; Melvin, L.S.; Howlett, A.C. Determination and characterization of a cannabinoid receptor in rat brain. Mol. Pharmacol. 1988, 34, 605–613. [Google Scholar]
- Matsuda, L.A.; Lolait, S.J.; Brownstein, M.J.; Young, A.C.; Bonner, T.I. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 1990, 346, 561–564. [Google Scholar] [CrossRef] [PubMed]
- Munro, S.; Thomas, K.L.; Abu-Shaar, M. Molecular characterization of a peripheral receptor for cannabinoids. Nature 1993, 365, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, D.G.; Ueda, N.; Yamamoto, S. The fatty acid amide hydrolase (FAAH). Prostaglandins Leukot Essent Fatty Acids 2002, 66, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Cravatt, B.F.; Giang, D.K.; Mayfield, S.P.; Boger, D.L.; Lerner, R.A.; Gilula, N.B. Molecular characterization of an enzyme that degrades neuromodulatory fatty-acid amides. Nature 1996, 384, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Sidhpura, N.; Parsons, L.H. Endocannabinoid-mediated synaptic plasticity and addiction-related behavior. Neuropharmacology 2011, 61, 1070–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zlebnik, N.E.; Cheer, J.F. Drug-induced alterations of endocannabinoid-mediated plasticity in brain reward regions. J. Neurosci. 2016, 36, 10230–10238. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.T.; Ogdie, M.N.; Jarvelin, M.R.; Moilanen, I.K.; Loo, S.K.; McCracken, J.T.; McGough, J.J.; Yang, M.H.; Peltonen, L.; Nelson, S.F.; et al. Association of the cannabinoid receptor gene (CNR1) with ADHD and post-traumatic stress disorder. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2008, 147B, 1488–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Moreno, J.A.; Echeverry-Alzate, V.; Buhler, K.M. The genetic basis of the endocannabinoid system and drug addiction in humans. J. Psychopharmacol. 2012, 26, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Sloan, M.E.; Gowin, J.L.; Yan, J.; Schwandt, M.L.; Spagnolo, P.A.; Sun, H.; Hodgkinson, C.A.; Goldman, D.; Ramchandani, V.A. Severity of alcohol dependence is associated with the fatty acid amide hydrolase Pro129Thr missense variant. Addict. Biol. 2018, 23, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Pava, M.J.; Woodward, J.J. A review of the interactions between alcohol and the endocannabinoid system: Implications for alcohol dependence and future directions for research. Alcohol 2012, 46, 185–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizrachi Zer-Aviv, T.; Segev, A.; Akirav, I. Cannabinoids and post-traumatic stress disorder: Clinical and preclinical evidence for treatment and prevention. Behav. Pharmacol. 2016, 27, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Akirav, I. Cannabinoids and glucocorticoids modulate emotional memory after stress. Neurosci. Biobehav. Rev. 2013, 37, 2554–2563. [Google Scholar] [CrossRef] [PubMed]
- Bluett, R.J.; Baldi, R.; Haymer, A.; Gaulden, A.D.; Hartley, N.D.; Parrish, W.P.; Baechle, J.; Marcus, D.J.; Mardam-Bey, R.; Shonesy, B.C.; et al. Endocannabinoid signalling modulates susceptibility to traumatic stress exposure. Nat. Commun. 2017, 8, 14782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, M.N.; Campolongo, P.; Yehuda, R.; Patel, S. Integrating endocannabinoid signaling and cannabinoids into the biology and treatment of posttraumatic stress disorder. Neuropsychopharmacology 2018, 43, 80–102. [Google Scholar] [CrossRef] [PubMed]
- Hillard, C.J. Circulating endocannabinoids: From whence do they come and where are they going? Neuropsychopharmacology 2018, 43, 155–172. [Google Scholar] [CrossRef] [PubMed]
- Morena, M.; Patel, S.; Bains, J.S.; Hill, M.N. Neurobiological interactions between stress and the endocannabinoid system. Neuropsychopharmacology 2016, 41, 80–102. [Google Scholar] [CrossRef] [PubMed]
- Worley, N.B.; Hill, M.N.; Christianson, J.P. Prefrontal endocannabinoids, stress controllability and resilience: A hypothesis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 85, 180–188. [Google Scholar] [CrossRef]
- Bluett, R.J.; Gamble-George, J.C.; Hermanson, D.J.; Hartley, N.D.; Marnett, L.J.; Patel, S. Central anandamide deficiency predicts stress-induced anxiety: Behavioral reversal through endocannabinoid augmentation. Transl. Psychiatry 2014, 8, e408. [Google Scholar] [CrossRef]
- Hill, M.N.; Kumar, S.A.; Filipski, S.B.; Iverson, M.; Stuhr, K.L.; Keith, J.M.; Cravatt, B.F.; Hillard, C.J.; Chattarji, S.; McEwen, B.S. Disruption of fatty acid amide hydrolase activity prevents the effects of chronic stress on anxiety and amygdalar microstructure. Mol. Psychiatry 2013, 18, 1125–1135. [Google Scholar] [CrossRef]
- Patel, S.; Kingsley, P.J.; Mackie, K.; Marnett, L.J.; Winder, D.G. Repeated homotypic stress elevates 2-arachidonoylglycerol levels and enhances short-term endocannabinoid signaling at inhibitory synapses in basolateral amygdala. Neuropsychopharmacology 2009, 34, 2699–2709. [Google Scholar] [CrossRef]
- Moreira, F.A.; Kaiser, N.; Monory, K.; Lutz, B. Reduced anxiety-like behaviour induced by genetic and pharmacological inhibition of the endocannabinoid-degrading enzyme fatty acid amide hydrolase (FAAH) is mediated by CB1 receptors. Neuropharmacology 2008, 54, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Gunduz-Cinar, O.; Hill, M.N.; McEwen, B.S.; Holmes, A. Amygdala FAAH and anandamide: Mediating protection and recovery from stress. Trends Pharmacol. Sci. 2013, 34, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Jenniches, I.; Ternes, S.; Albayram, O.; Otte, D.M.; Bach, K.; Bindila, L.; Michel, K.; Lutz, B.; Bilkei-Gorzo, A.; Zimmer, A. Anxiety, stress, and fear response in mice with reduced endocannabinoid levels. Biol. Psychiatry 2016, 79, 858–868. [Google Scholar] [CrossRef] [PubMed]
- Lissek, S.; van Meurs, B. Learning models of PTSD: Theoretical accounts and psychobiological evidence. Int. J. Psychophysiol. 2015, 98, 594–605. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.; Falls, W.A.; Campeau, S.; Kim, M. Fear-potentiated startle: A neural and pharmacological analysis. Behav. Brain Res. 1993, 58, 175–198. [Google Scholar] [CrossRef]
- Grillon, C. Models and mechanisms of anxiety: Evidence from startle studies. Psychopharmacology 2008, 199, 421–437. [Google Scholar] [CrossRef] [PubMed]
- Groenink, L.; Bijlsma, E.Y.; Olivier, B. Fear-potentiated startle and light-enhanced startle models in drug discovery. Curr. Protoc. Pharmacol. 2008, 48. [Google Scholar] [CrossRef]
- Hijzen, T.H.; Houtzager, S.W.; Joordens, R.J.; Olivier, B.; Slangen, J.L. Predictive validity of the potentiated startle response as a behavioral model for anxiolytic drugs. Psychopharmacology 1995, 118, 150–154. [Google Scholar] [CrossRef]
- Barrenha, G.D.; Chester, J.A. Genetic correlation between innate alcohol preference and fear-potentiated startle in selected mouse lines. Alcohol. Clin. Exp. Res. 2007, 31, 1081–1088. [Google Scholar] [CrossRef]
- Barrenha, G.D.; Coon, L.E.; Chester, J.A. Effects of alcohol on the acquisition and expression of fear-potentiated startle in mouse lines selectively bred for high and low alcohol preference. Psychopharmacology 2011, 218, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Chester, J.A.; Kirchhoff, A.M.; Barrenha, G.D. Relation between corticosterone and fear-related behavior in mice selectively bred for high or low alcohol preference. Addict. Biol. 2014, 19, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Crabbe, J.C.; Phillips, T.J.; Kosobud, A.; Belknap, J.K. Estimation of genetic correlation: Interpretation of experiments using selectively bred and inbred animals. Alcohol Clin. Exp. Res. 1990, 14, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Powers, M.S.; Barrenha, G.D.; Mlinac, N.S.; Barker, E.L.; Chester, J.A. Effects of the novel endocannabinoid uptake inhibitor, LY2183240, on fear-potentiated startle and alcohol-seeking behaviors in mice selectively bred for high alcohol preference. Psychopharmacology 2010, 212, 571–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherma, M.; Masia, P.; Satta, V.; Fratta, W.; Fadda, P.; Tanda, G. Brain activity of anandamide: A rewarding bliss? Acta Pharmacol. Sin. 2019, 40, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Arnold, J.C.; Topple, A.N.; Mallet, P.E.; Hunt, G.E.; McGregor, I.S. The distribution of cannabinoid-induced Fos expression in rat brain: Differences between the Lewis and Wistar strain. Brain Res. 2001, 921, 240–255. [Google Scholar] [CrossRef]
- Herkenham, M.; Lynn, A.B.; Johnson, M.R.; Melvin, L.S.; de Costa, B.R.; Rice, K.C. Characterization and localization of cannabinoid receptors in rat brain: A quantitative in vitro autoradiographic study. J. Neurosci. 1991, 11, 563–583. [Google Scholar] [CrossRef]
- Cippitelli, A.; Bilbao, A.; Hansson, A.C.; del Arco, I.; Sommer, W.; Heilig, M.; Massi, M.; Bermudez-Silva, F.J.; Navarro, M.; Ciccocioppo, R.; et al. Cannabinoid CB1 receptor antagonism reduces conditioned reinstatement of ethanol-seeking behavior in rats. Eur. J. Neurosci. 2005, 21, 2243–2251. [Google Scholar] [CrossRef] [PubMed]
- Oberlin, B.; Best, C.; Matson, L.; Henderson, A.; Grahame, N. Derivation and characterization of replicate high- and low-alcohol preferring lines of mice and a high-drinking crossed HAP line. Behav. Genet. 2011, 41, 288–302. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates, 2nd ed.; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- Han, B.; Wright, R.; Kirchhoff, A.M.; Chester, J.A.; Cooper, B.R.; Davisson, V.J.; Barker, E. Quantitative LC-MS/MS analysis of arachidonoyl amino acids in mouse brain with treatment of FAAH inhibitor. Anal. Biochem. 2013, 432, 74–81. [Google Scholar] [CrossRef]
- Cooper, R.L.; Goldman, J.M.; Vandenbergh, J.G. Monitoring of the estrous cycle in the laboratory rodent by vaginal lavage. In Methods in Toxicology; Chapin, R.E., Heindel, J.J., Eds.; Academic Press: Orlando, FL, USA, 1993; Volume 3B, pp. 45–56. [Google Scholar]
- Walker, D.L.; Davis, M. Quantifying fear potentiated startle using absolute versus proportional increase scoring methods: Implications for the neurocircuitry of fear and anxiety. Psychopharmacology 2002, 164, 318–328. [Google Scholar] [CrossRef]
- Dixon, W.J. Analysis of extreme values. Ann. Mathemat. Stat. 1950, 21, 488–506. [Google Scholar] [CrossRef]
- Hill, M.N.; McLaughlin, R.J.; Morrish, A.C.; Viau, V.; Floresco, S.B.; Hillard, C.J.; Gorzalka, B.B. Suppression of amygdalar endocannabinoid signaling by stress contributes to activation of the hypothalamic-pituitary-adrenal axis. Neuropsychopharmacology 2009, 34, 2733–2745. [Google Scholar] [CrossRef] [PubMed]
- Rademacher, D.J.; Meier, S.E.; Shi, L.; Ho, W.S.; Jarrahian, A.; Hillard, C.J. Effects of acute and repeated restraint stress on endocannabinoid content in the amygdala, ventral striatum, and medial prefrontal cortex in mice. Neuropharmacology 2008, 54, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Cohen, H.; Yehuda, R. Gender differences in animal models of posttraumatic stress disorder. Dis. Markers 2011, 30, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Mazor, A.; Matar, M.A.; Kaplan, Z.; Kozlovsky, N.; Zohar, J.; Cohen, H. Gender-related qualitative differences in baseline and post-stress anxiety responses are not reflected in the incidence of criterion-based PTSD-like behaviour patterns. World J. Biol. Psychiatry 2009, 10, 856–869. [Google Scholar] [CrossRef] [PubMed]
- Ney, L.J.; Matthews, A.; Bruno, R.; Felmingham, K.L. Modulation of the endocannabinoid system by sex hormones: Implications for posttraumatic stress disorder. Neurosci. Biobehav. Rev. 2018, 94, 302–320. [Google Scholar] [CrossRef]
- Zer-Aviv, T.M.; Akirav, I. Sex differences in hippocampal response to endocannabinoids after exposure to severe stress. Hippocampus 2016, 26, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Sabatucci, A.; Simonetti, M.; Tortolani, D.; Angelucci, C.B.; Dainese, E.; Maccarrone, M. Role of steroids on the membrane binding ability of fatty acid amide hydrolase. Cannabis Cannabinoid Res. 2019, 4, 42–50. [Google Scholar] [CrossRef]
- Sumislawski, J.J.; Ramikie, T.S.; Patel, S. Reversible gating of endocannabinoid plasticity in the amygdala by chronic stress: A potential role for monoacylglycerol lipase inhibition in the prevention of stress-induced behavioral adaptation. Neuropsychopharmacology 2011, 36, 2750–2761. [Google Scholar] [CrossRef]
- Morena, M.; Aukema, R.J.; Leitl, K.D.; Rashid, A.J.; Vecchiarelli, H.A.; Josselyn, S.A.; Hill, M.N. Upregulation of anandamide hydrolysis in the basolateral complex of amygdala reduces fear memory expression and indices of stress and anxiety. J. Neurosci. 2019, 39, 1275–1292. [Google Scholar] [CrossRef]
- Segev, A.; Korem, N.; Mizrachi Zer-Aviv, T.; Abush, H.; Lange, R.; Sauber, G.; Hillard, C.J.; Akirav, I. Role of endocannabinoids in the hippocampus and amygdala in emotional memory and plasticity. Neuropsychopharmacology 2018, 43, 2017–2027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, M.N.; Tasker, J.G. Endocannabinoid signaling, glucocorticoid-mediated negative feedback, and regulation of the hypothalamic-pituitary-adrenal axis. Neuroscience 2012, 204, 5–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Oliveira Alvares, L.; Engelke, D.S.; Diehl, F.; Scheffer-Teixeira, R.; Haubrich, J.; de Freitas Cassini, L.; Molina, V.A.; Quillfeldt, J.A. Stress response recruits the hippocampal endocannabinoid system for the modulation of fear memory. Learn. Mem. 2010, 17, 202–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutz, B.; Marsicano, G.; Maldonado, R.; Hillard, C.J. The endocannabinoid system in guarding against fear, anxiety and stress. Nat. Rev. Neurosci. 2015, 16, 705–718. [Google Scholar] [CrossRef] [PubMed]
- Campolongo, P.; Roozendaal, B.; Trezza, V.; Hauer, D.; Schelling, G.; McGaugh, J.L.; Cuomo, V. Endocannabinoids in the rat basolateral amygdala enhance memory consolidation and enable glucocorticoid modulation of memory. Proc. Natl. Acad. Sci. USA 2009, 106, 4888–4893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganon-Elazar, E.; Akirav, I. Cannabinoids and traumatic stress modulation of contextual fear extinction and GR expression in the amygdala-hippocampal-prefrontal circuit. Psychoneuroendocrinology 2013, 38, 1675–1687. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.M.; Vecchiarelli, H.A.; Morena, M.; Lee, T.T.; Hermanson, D.J.; Kim, A.B.; McLaughlin, R.J.; Hassan, K.I.; Kuhne, C.; Wotjak, C.T.; et al. Corticotropin-releasing hormone drives anandamide hydrolysis in the amygdala to promote anxiety. J. Neurosci. 2015, 35, 3879–3892. [Google Scholar] [CrossRef]
- Hill, M.N.; McLaughlin, R.J.; Pan, B.; Fitzgerald, M.L.; Roberts, C.J.; Lee, T.T.; Karatsoreos, I.N.; Mackie, K.; Viau, V.; Pickel, V.M.; et al. Recruitment of prefrontal cortical endocannabinoid signaling by glucocorticoids contributes to termination of the stress response. J. Neurosci. 2011, 31, 10506–10515. [Google Scholar] [CrossRef]
- Bitencourt, R.M.; Pamplona, F.A.; Takahashi, R.N. Corticosteroid-endocannabinoid loop supports decrease of fear-conditioned response in rats. Eur. Neuropsychopharmacol. 2014, 24, 1091–1102. [Google Scholar] [CrossRef]
- Rey, A.A.; Purrio, M.; Viveros, M.P.; Lutz, B. Biphasic effects of cannabinoids in anxiety responses: CB1 and GABA(B) receptors in the balance of GABAergic and glutamatergic neurotransmission. Neuropsychopharmacology 2012, 37, 2624–2634. [Google Scholar] [CrossRef]
- Tambaro, S.; Tomasi, M.L.; Bortolato, M. Long-term CB(1) receptor blockade enhances vulnerability to anxiogenic-like effects of cannabinoids. Neuropharmacology 2013, 70, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Franklin, J.M.; Mathew, M.; Carrasco, G.A. Cannabinoid-induced upregulation of serotonin 2A receptors in the hypothalamic paraventricular nucleus and anxiety-like behaviors in rats. Neurosci. Lett. 2013, 548, 165–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caligioni, C.S. Assessing reproductive status/stages in mice. Curr. Protoc. Neurosci. 2009, 48. [Google Scholar] [CrossRef]
- Gupta, R.R.; Sen, S.; Diepenhorst, L.L.; Rudick, C.N.; Maren, S. Estrogen modulates sexually dimorphic contextual fear conditioning and hippocampal long-term potentiation (LTP) in rats. Brain Res. 2001, 888, 356–365. [Google Scholar] [CrossRef]
- Zhao, Y.; Bijlsma, E.Y.; Verdouw, M.P.; Groenink, L. No effect of sex and estrous cycle on the fear potentiated startle response in rats. Behav. Brain Res. 2018, 351, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Hiroi, R.; Neumaier, J.F. Differential effects of ovarian steroids on anxiety versus fear as measured by open field test and fear-potentiated startle. Behav. Brain Res. 2006, 166, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Voulo, M.E.; Parsons, R.G. Gonadal hormone fluctuations do not affect the expression or extinction of fear-potentiated startle in female rats. Behav. Neurosci. 2019. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirchhoff, A.M.; Barker, E.L.; Chester, J.A. Endocannabinoids and Fear-Related Behavior in Mice Selectively Bred for High or Low Alcohol Preference. Brain Sci. 2019, 9, 254. https://doi.org/10.3390/brainsci9100254
Kirchhoff AM, Barker EL, Chester JA. Endocannabinoids and Fear-Related Behavior in Mice Selectively Bred for High or Low Alcohol Preference. Brain Sciences. 2019; 9(10):254. https://doi.org/10.3390/brainsci9100254
Chicago/Turabian StyleKirchhoff, Aaron M., Eric L. Barker, and Julia A. Chester. 2019. "Endocannabinoids and Fear-Related Behavior in Mice Selectively Bred for High or Low Alcohol Preference" Brain Sciences 9, no. 10: 254. https://doi.org/10.3390/brainsci9100254
APA StyleKirchhoff, A. M., Barker, E. L., & Chester, J. A. (2019). Endocannabinoids and Fear-Related Behavior in Mice Selectively Bred for High or Low Alcohol Preference. Brain Sciences, 9(10), 254. https://doi.org/10.3390/brainsci9100254