Electrical Stimulation of C6 Glia-Precursor Cells In Vitro Differentially Modulates Gene Expression Related to Chronic Pain Pathways

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. MTT Cell Viability Assay
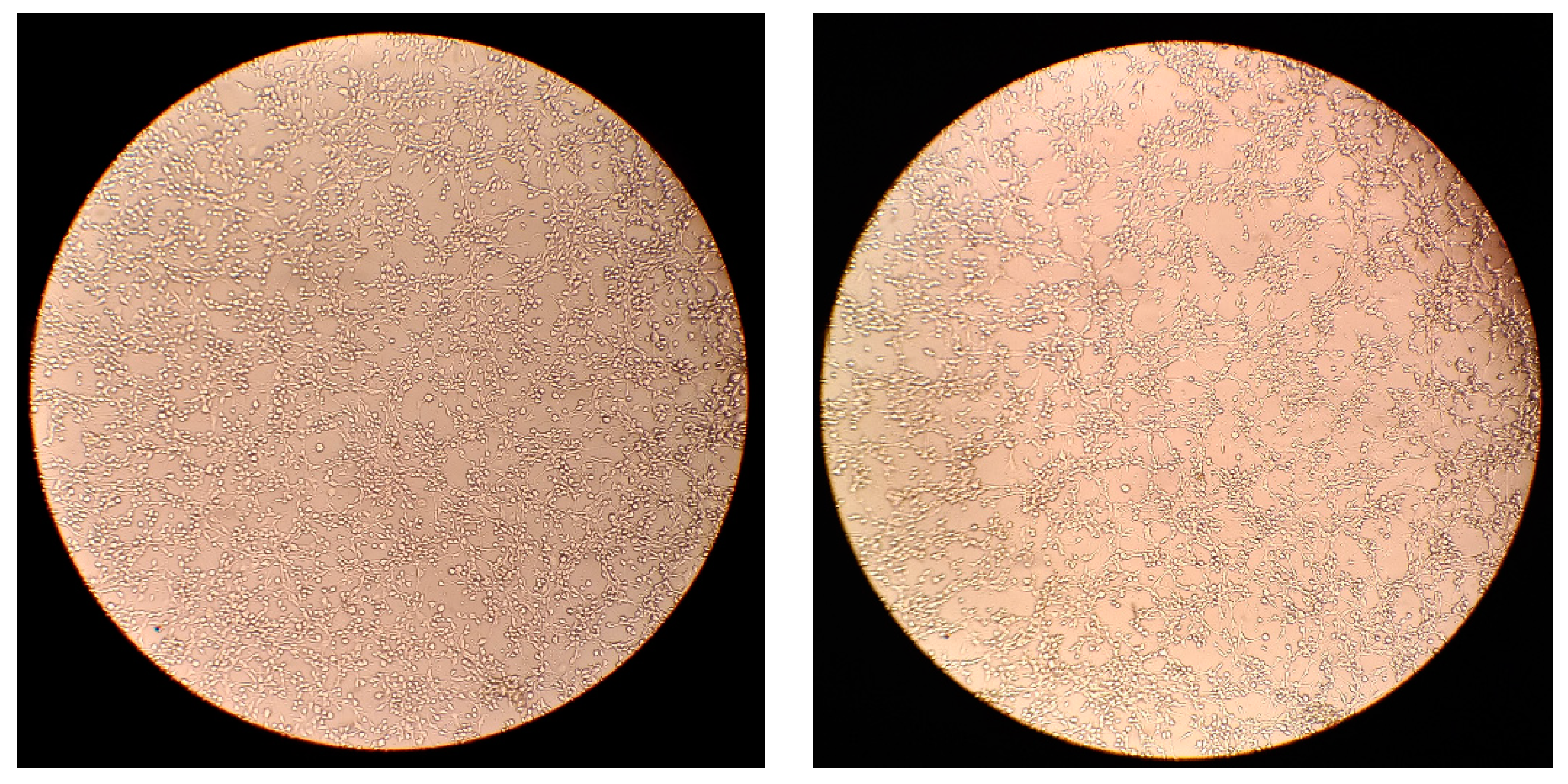
3.2. Light Microscopy
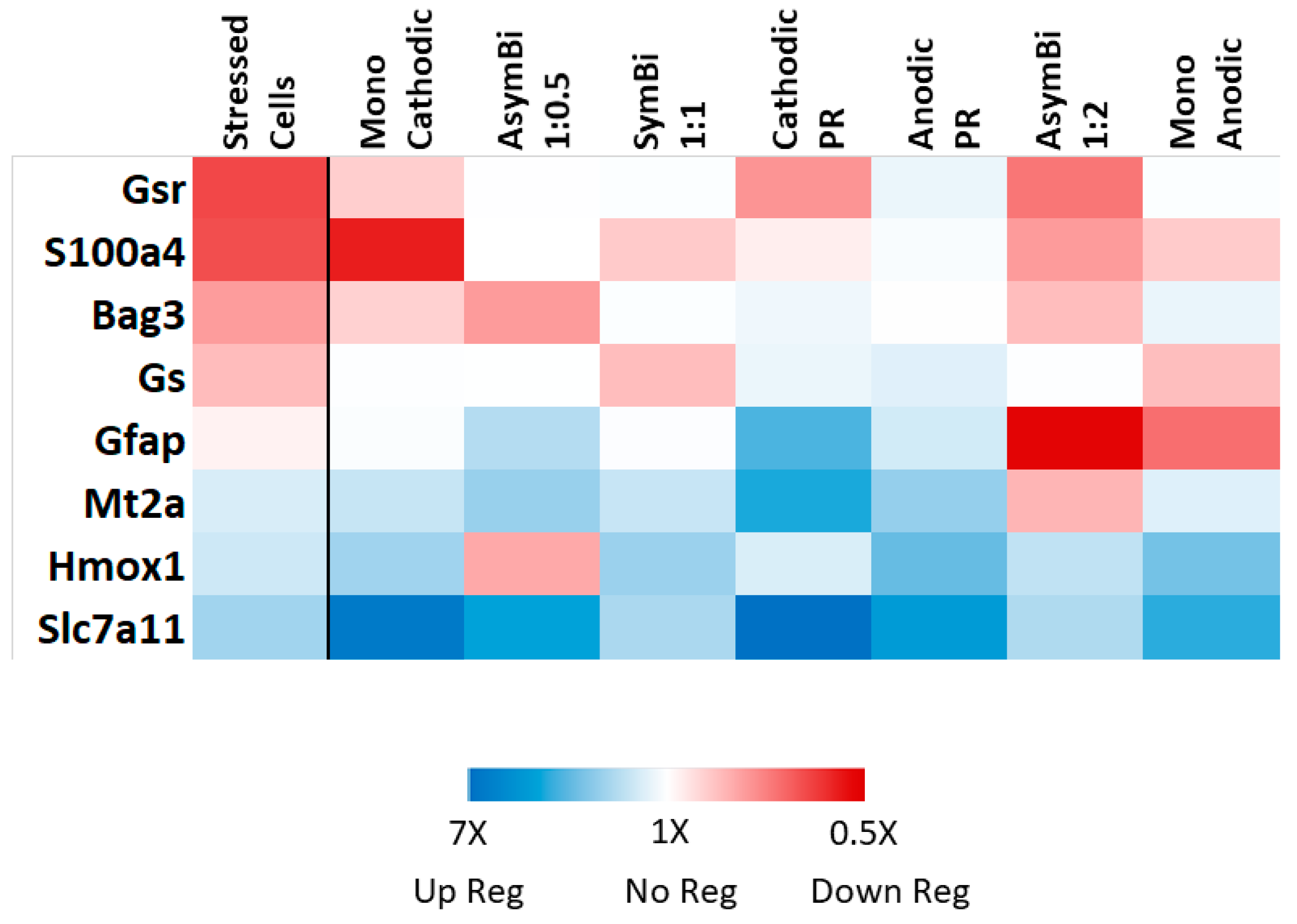
3.3. RT-qPCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Verkhratsky, A.; Kirchhoff, F. Glutamate-mediated neuronal–glial transmission. J. Anat. 2007, 210, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Salvemini, D.; Little, J.W.; Doyle, T.; Neumann, W.L. Roles of reactive oxygen and nitrogen species in pain. Free Radic. Biol. Med. 2011, 51, 951–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kung, L.-H.; Gong, K.; Adedoyin, M.; Ng, J.; Bhargava, A.; Ohara, P.T.; Jasmin, L. Evidence for glutamate as a neuroglial transmitter within sensory ganglia. PLoS ONE 2013, 8, e68312. [Google Scholar] [CrossRef] [PubMed]
- Verkhratsky, A.; Kettenmann, H. Calcium signalling in glial cells. Trends Neurosci. 1996, 19, 346–352. [Google Scholar] [CrossRef]
- Milligan, E.D.; Watkins, L.R. Pathological and protective roles of glia in chronic pain. Nat. Rev. Neurosci. 2009, 10, 23–36. [Google Scholar] [CrossRef]
- Roitbak, A.I.; Fanardjian, V.V. Depolarization of cortical glial cells in response to electrical stimulation of the cortical surface. Neuroscience 1981, 6, 2529–2537. [Google Scholar] [CrossRef]
- Agnesi, F.; Blaha, C.D.; Lin, J.; Lee, K.H. Local glutamate release in the rat ventral lateral thalamus evoked by high-frequency stimulation. J. Neural Eng. 2010, 7, 26009. [Google Scholar] [CrossRef]
- Tawfik, V.L.; Chang, S.Y.; Hitti, F.L.; Roberts, D.W.; Leiter, J.C.; Jovanovic, S.; Lee, K.H. Deep brain stimulation results in local glutamate and adenosine release: Investigation into the role of astrocytes. Neurosurgery 2010, 67, 367–375. [Google Scholar] [CrossRef]
- Lee, K.H.; Hitti, F.L.; Chang, S.Y.; Lee, D.C.; Roberts, D.W.; McIntyre, C.C.; Leiter, J.C. High frequency stimulation abolishes thalamic network oscillations: An electrophysiological and computational analysis. J. Neural Eng. 2011, 8, 046001. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Hozumi, Y.; Kaneko, K.; Sugihara, T.; Fujii, S.; Goto, K.; Kato, H. Modulatory effects of oligodendrocytes on the conduction velocity of action potentials along axons in the alveus of the rat hippocampal CA1 region. Neuron Glia Biol. 2007, 3, 325–334. [Google Scholar] [CrossRef]
- Vallejo, R.; Bradley, K.; Kapural, L. Spinal cord stimulation in chronic pain: Mode of action. Spine 2017, 42, S53–S60. [Google Scholar] [CrossRef] [PubMed]
- Vallejo, R.; Tilley, D.M.; Vogel, L.; Benyamin, R. The role of glia and the immune system in the development and maintenance of neuropathic pain. Pain Pract. 2010, 10, 167–184. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Sauri, A.; Orduña-Valls, J.M.; Blasco-Serra, A.; Tornero-Tornero, C.; Cedeño, D.L.; Bejarano-Quisoboni, D.; Valverde-Navarro, A.A.; Benyamin, R.; Vallejo, R. Glia to neuron ratio in the posterior aspect of the human spinal cord at thoracic segments relevant to spinal cord stimulation. J. Anat. 2019, 235, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Stephens, K.E.; Chen, Z.; Sivanesan, E.; Raja, S.N.; Linderoth, B.; Taverna, S.D.; Guan, Y. RNA-seq of spinal cord from nerve-injured rats after spinal cord stimulation. Mol. Pain 2018, 14. [Google Scholar] [CrossRef] [Green Version]
- Tilley, D.M.; Cedeño, D.L.; Kelley, C.A.; Benyamin, R.; Vallejo, R. Spinal cord stimulation modulates gene expression in the spinal cord of an animal model of peripheral nerve injury. Reg. Anesth. Pain Med. 2016, 41, 750–756. [Google Scholar] [CrossRef]
- Vallejo, R.; Tilley, D.M.; Cedeño, D.L.; Kelley, C.A.; DeMaegd, M.; Benyamin, R. Genomics of the Effect of Spinal Cord Stimulation on an Animal Model of Neuropathic Pain. Neuromodulation 2016, 19, 576–586. [Google Scholar] [CrossRef]
- Benda, P.; Lightbody, J.; Sato, G.; Levine, L.; Sweet, W. Differentiated rat glial cell strain in tissue culture. Science 1968, 161, 370–371. [Google Scholar] [CrossRef]
- Brodie, C.; Kuperstein, I.; Acs, P.; Blumberg, P.M. Differential role of specific PKC isoforms in the proliferation of glial cells and the expression of the astrocytic markers GFAP and glutamine synthetase. Mol. Brain Res. 1998, 56, 108–117. [Google Scholar] [CrossRef]
- Chao, C.; Kan, D.; Lo, T.; Lu, K.; Chien, C. Induction of neural differentiation in rat C6 glioma cells with taxol. Brain Behav. 2015, 5, e00414. [Google Scholar] [CrossRef]
- Mawatari, K.; Yasui, Y.; Sugitani, K.; Takadera, T.; Kato, S. Reactive oxygen species involved in the glutamate toxicity of C6 glioma cells via xc antiporter system. Neuroscience 1996, 73, 201–208. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Kirchhoff, F. NMDA receptors in glia. Neuroscientist 2007, 13, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Sen, C.K.; Roy, S.; Kobayashi, M.S.; Tritschler, H.J.; Packer, L. Protection against glutamate-induced cytotoxicity in C6 glial cells by thiol antioxidants. Am. J. Physiol. Integr. Comp. Physiol. 1997, 273, R1771–R1778. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Negishi, K.; Mawatari, K.; Kuo, C.-H. A mechanism for glutamate toxicity in the C6 glioma cells involving inhibition of cystine uptake leading to glutathione depletion. Neuroscience 1992, 48, 903–914. [Google Scholar] [CrossRef]
- Sribnick, E.A.; Ray, S.K.; Banik, N.L. Estrogen prevents glutamate-induced apoptosis in C6 glioma cells by a receptor-mediated mechanism. Neuroscience 2006, 137, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Cookson, M.R.; Mead, C.; Austwick, S.M.; Pentreath, V.W. Use of the MTT assay for estimating toxicity in primary astrocyte and C6 glioma cell cultures. Toxicol. In Vitro 1995, 9, 39–48. [Google Scholar] [CrossRef]
- Vallejo, R.; Tilley, D.M.; Williams, J.; Labak, S.; Aliaga, L.; Benyamin, R.M. Pulsed radiofrequency modulates pain regulatory gene expression along the nociceptive pathway. Pain Physician 2013, 16, E601–E613. [Google Scholar] [PubMed]
- Sabolić, I.; Škarica, M.; Ljubojević, M.; Breljak, D.; Herak-Kramberger, C.M.; Crljen, V.; Ljubešić, N. Expression and immunolocalization of metallothioneins MT1, MT2 and MT3 in rat nephron. J. Trace Elem. Med. Biol. 2018, 46, 62–75. [Google Scholar] [CrossRef] [Green Version]
- Kent, W.J.; Sugnet, C.W.; Furey, T.S.; Roskin, K.M.; Pringle, T.H.; Zahler, A.M.; Haussler, D. The human genome browser at UCSC. Genome Res. 2002, 12, 996–1006. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]
- Kibbe, W.A. OligoCalc: An online oligonucleotide properties calculator. Nucleic Acids Res. 2007, 35, W43–W46. [Google Scholar] [CrossRef]
- Haimes, J.; Kelley, M. Demonstration of a ΔΔCq Calculation Method to Compute Relative Gene Expression from qPCR Data; GE Healthcare Tech Note; GE Healthcare UK Limited: Buckinghamshire, UK, 2015. [Google Scholar]
- Poss, K.D.; Tonegawa, S. Reduced stress defense in heme oxygenase 1-deficient cells. Proc. Natl. Acad. Sci. USA 1997, 94, 10925–10930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, X.-B.; Wei, H.-W.; Wang, J.; Kong, Y.-Q.; Wu, Y.-Y.; Guo, J.-L.; Li, T.-F.; Li, J.-K. Mammalian metallothionein-2A and oxidative stress. Int. J. Mol. Sci. 2016, 17, 1483. [Google Scholar] [CrossRef] [PubMed]
- Shin, C.-S.; Mishra, P.; Watrous, J.D.; Carelli, V.; D’Aurelio, M.; Jain, M.; Chan, D.C. The glutamate/cystine xCT antiporter antagonizes glutamine metabolism and reduces nutrient flexibility. Nat. Commun. 2017, 8, 15074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosati, A.; Graziano, V.; De Laurenzi, V.; Pascale, M.; Turco, M.C. BAG3: A multifaceted protein that regulates major cell pathways. Cell Death Dis. 2011, 2, e141. [Google Scholar] [CrossRef] [PubMed]
- Dmytriyeva, O.; Pankratova, S.; Owczarek, S.; Sonn, K.; Soroka, V.; Ridley, C.M.; Marsolais, A.; Lopez-Hoyos, M.; Ambartsumian, N.; Lukanidin, E. The metastasis-promoting S100A4 protein confers neuroprotection in brain injury. Nat. Commun. 2012, 3, 1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallejo, R.; Gupta, A.; Kelley, C.A.; Vallejo, A.; Rink, J.; Williams, J.M.; Cass, C.L.; Smith, W.J.; Benyamin, R.; Cedeño, D.L. Effects of Phase Polarity and Charge Balance Spinal Cord Stimulation on Behavior and Gene Expression in a Rat Model of Neuropathic Pain. Neuromodulation 2019. [Google Scholar] [CrossRef]
- Ji, R.R.; Berta, T.; Nedergaard, M. Glia and pain: Is chronic pain a gliopathy? Pain 2013, 154, S10–S28. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Group Name | Stimulation Parameters | Charge Balance | Duty Cycle | Cathodic Charge | Anodic Charge | Waveform Shape |
|---|---|---|---|---|---|---|
| Monophasic Cathodic | F = 50 Hz PW = 50 μs | Unbalanced | 0.25% | 7.5 nC | 0 |  |
| Asymmetric Biphasic 1:0.5 (AsymBi 1:0.5) | F = 50 Hz PW = 50 μs cathodic, 25 μs anodic | Active unbalanced | 0.38% | 7.5 nC | 3.8 nC |  |
| Symmetric Biphasic 1:1 (SymBi 1:1) | F = 50 Hz PW = 50 μs per phase | Active balanced | 0.50% | 7.5 nC | 7.5 nC |  |
| Cathodic PR | F = 50 Hz PW = 60μs ‡ | Passive balanced | 5% * | 9.0 nC | 9.0 nC |  |
| Anodic PR | F = 50 Hz PW = 60 μs ‡ | Passive balanced | 5% * | 9.0 nC | 9.0 nC |  |
| Asymmetric Biphasic 1:2 (AsymBi 1:2) | F = 50 Hz PW = 50 μs cathodic, 100 μs anodic | Active unbalanced | 0.75% | 7.5 nC | 15 nC |  |
| Monophasic Anodic | F = 50 Hz PW = 50 μs | Unbalanced | 0.25% | 0 | 7.5 nC |  |
| Gene | Codes for | Accession Number | Sequence | Direction | Role |
|---|---|---|---|---|---|
| Gfap | Glial fibrillary acidic protein | NM_017009 | 5′-CAGGAAATTGCTGGAGGGCGAA-3′ | Forward | Immune function |
| 5′-CTTGAGGTGGCCTTCTGACACAG-3′ | Reverse | ||||
| Slc7a11 | Glutamate cystine transporter | NM_001107673 | 5′-ACAAACGCCCAGATATGCATCGTC-3′ | Forward | Synaptic transmission |
| 5′-GGTGCTAAACGGATCCGAGTAAAGG-3′ | Reverse | ||||
| Glul | Glutamine synthetase | M29579 | 5’-GCCTTCTAATGGCTTCCCTGGAC-3’ | Forward | Synaptic transmission |
| 5’-ACCTCGGCATTTGTCCCTGTG-3’ | Reverse | ||||
| S100a4 | S100 calcium binding protein A4 | NM_012618 | 5′-GCACTTCCTCTCTCTTGGTCT-3′ | Forward | Neuro-protection |
| 5′-GTCTGTCCTTCTCCCCAGGA-3′ | Reverse | ||||
| Mt2a | Metallothionein 2A | NM_001137564 | 5′-CACAGATGGATCCTGCTCCT-3′ | Forward | Redox process |
| 5′-AAGTGTGGAGAACCGGTCAG-3′ | Reverse | ||||
| Gsr | Glutathione-disulfide reductase | NM_053906 | 5′-GATGTATCACGCTGTGACCACGAG-3′ | Forward | Redox process |
| 5′-AGCATCTCATCGCAGCCAATCC-3′ | Reverse | ||||
| Hmox1 | Heme oxygenase 1 | NM_012580 | 5′-TGCTCGCATGAACACTCTGGAG-3′ | Forward | Redox process |
| 5′-GACTCTGGTCTTTGTGTTCCTCTGTC-3′ | Reverse | ||||
| Bag3 | Bcl2 associated athanogene 3 | NM_001011936 | 5′-CAGACAGATAAACAGTGTGGACAGGTG-3′ | Forward | Cell adaptive response |
| 5′-AGGACGAGGATGAGCAGTCAGAG-3′ | Reverse | ||||
| Gapdh | Glyceraldehyde-3-phosphate dehydrogenase | NM_017008 | 5′-CTCATGACCACAGTCCATGC-3′ | Forward | House-keeping/Control |
| 5′-TTCAGCTCTGGGATGACCTT-3′ | Reverse |
| Biological Process | Gene | No-ES_Glu vs. No-ES_No-Glu |
|---|---|---|
| Immune Function | Gfap | 0.965 |
| Synaptic Transmission | Slc7a11 | 2.859 * |
| Glul | 0.847 | |
| Neuroprotection | S100a4 | 0.653 * |
| Redox Processes | Mt2a | 1.797 * |
| Gsr | 0.639 * | |
| Hmox1 | 1.991 * | |
| Cell Adaptive Response | Bag3 | 0.801 * |
| Biological Process | Gene | ES_Glu vs. No-ES_Glu | ||||||
|---|---|---|---|---|---|---|---|---|
| Monophasic Cathodic | AsymBi 1:0.5 | SymBi 1:1 | Cathodic PR | Anodic PR | AsymBi 1:2 | Monophasic Anodic | ||
| Immune Function | Gfap | 1.125 | 2.531 ‡ | 1.099 | 4.335 * | 1.931 | 0.532 | 0.723 |
| Synaptic Transmission | Slc7a11 | 2.274 * | 1.108 | 0.928 | 2.477 * | 1.903 ‡ | 0.907 | 1.589 |
| Glul | 1.198 | 1.171 | 1.001 | 1.596 | 1.889 * | 1.221 | 0.998 | |
| Neuroprotection | S100a4 | 0.861 | 1.492 | 1.351 | 1.432 | 1.710 | 1.221 | 1.351 |
| Redox Processes | Mt2a | 1.185 | 1.588 | 1.166 | 2.574 * | 1.660 | 0.470 ‡ | 0.914 |
| Gsr | 1.386 | 1.592 | 1.655 | 1.263 | 2.154 * | 1.133 | 1.667 | |
| Hmox1 | 1.421 | 0.401 * | 1.468 | 0.879 | 1.910 | 1.128 | 1.795 | |
| Cell Adaptive Response | Bag3 | 1.120 | 0.975 | 1.319 | 1.654 | 1.231 | 1.076 | 1.794 |
| Biological Process | Gene | ES_No-Glu vs. No-ES_No-Glu | ||||||
|---|---|---|---|---|---|---|---|---|
| Monophasic Cathodic | AsymBi 1:0.5 | SymBi 1:1 | Cathodic PR | Anodic PR | AsymBi 1:2 | Monophasic Anodic | ||
| Immune Function | Gfap | 0.821 | 0.745 | 0.802 | 1.469 | 1.653 | 0.488 * | 0.700 |
| Synaptic Transmission | Slc7a11 | 0.914 | 0.690 | 0.596 | 1.204 | 1.924 ‡ | 0.843 | 0.921 |
| Glul | 0.659 | 1.469 | 1.068 | 1.111 | 1.108 | 0.724 | 0.580 ‡ | |
| Neuroprotection | S100a4 | 0.632 ‡ | 1.072 | 0.923 | 1.162 | 1.468 | 0.617 ‡ | 0.449 * |
| Redox Processes | Mt2a | 0.521 | 1.310 | 1.392 | 2.445 ‡ | 2.875 * | 0.722 | 0.444 |
| Gsr | 0.875 | 0.654 | 1.134 | 1.179 | 1.106 | 0.807 | 0.980 | |
| Hmox1 | 1.342 | 0.679 | 0.625 | 0.712 | 1.136 | 0.701 | 2.029 | |
| Cell Adaptive Response | Bag3 | 0.948 | 0.939 | 0.880 | 1.102 | 1.484 | 1.154 | 1.057 |
| Biological Process | Gene | ES_Glu vs. No-ES_No-Glu | ||||||
|---|---|---|---|---|---|---|---|---|
| Monophasic Cathodic | AsymBi 1:0.5 | SymBi 1:1 | Cathodic PR | Anodic PR | AsymBi 1:2 | Monophasic Anodic | ||
| Immune Function | Gfap | 1.085 | 2.442 * | 1.061 | 4.184 * | 1.864 ‡ | 0.513 ‡ | 0.698 |
| Synaptic Transmission | Slc7a11 | 6.502 * | 3.170 * | 2.653 * | 7.083 * | 5.443 * | 2.592 * | 4.545 * |
| Glul | 1.015 | 0.991 | 0.848 | 1.352 | 1.600 | 1.034 | 0.846 | |
| Neuroprotection | S100a4 | 0.563 | 0.975 | 0.883 | 0.935 | 1.118 | 0.798 | 0.883 |
| Redox Processes | Mt2a | 2.129 | 2.855 * | 2.095 | 4.627 * | 2.983 * | 0.845 | 1.643 |
| Gsr | 0.886 | 1.017 | 1.058 | 0.807 | 1.377 | 0.724 | 1.065 | |
| Hmox1 | 2.830 * | 0.798 | 2.923 * | 1.750 | 3.804 * | 2.246 * | 3.574 * | |
| Cell Adaptive Response | Bag3 | 0.897 | 0.781 | 1.057 | 1.325 | 0.986 | 0.862 | 1.437 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vallejo, R.; Platt, D.C.; Rink, J.A.; Jones, M.A.; Kelley, C.A.; Gupta, A.; Cass, C.L.; Eichenberg, K.; Vallejo, A.; Smith, W.J.; et al. Electrical Stimulation of C6 Glia-Precursor Cells In Vitro Differentially Modulates Gene Expression Related to Chronic Pain Pathways. Brain Sci. 2019, 9, 303. https://doi.org/10.3390/brainsci9110303
Vallejo R, Platt DC, Rink JA, Jones MA, Kelley CA, Gupta A, Cass CL, Eichenberg K, Vallejo A, Smith WJ, et al. Electrical Stimulation of C6 Glia-Precursor Cells In Vitro Differentially Modulates Gene Expression Related to Chronic Pain Pathways. Brain Sciences. 2019; 9(11):303. https://doi.org/10.3390/brainsci9110303
Chicago/Turabian StyleVallejo, Ricardo, David C. Platt, Jonathan A. Rink, Marjorie A. Jones, Courtney A. Kelley, Ashim Gupta, Cynthia L. Cass, Kirk Eichenberg, Alejandro Vallejo, William J. Smith, and et al. 2019. "Electrical Stimulation of C6 Glia-Precursor Cells In Vitro Differentially Modulates Gene Expression Related to Chronic Pain Pathways" Brain Sciences 9, no. 11: 303. https://doi.org/10.3390/brainsci9110303
APA StyleVallejo, R., Platt, D. C., Rink, J. A., Jones, M. A., Kelley, C. A., Gupta, A., Cass, C. L., Eichenberg, K., Vallejo, A., Smith, W. J., Benyamin, R., & Cedeño, D. L. (2019). Electrical Stimulation of C6 Glia-Precursor Cells In Vitro Differentially Modulates Gene Expression Related to Chronic Pain Pathways. Brain Sciences, 9(11), 303. https://doi.org/10.3390/brainsci9110303