Attention Modulates Electrophysiological Responses to Simultaneous Music and Language Syntax Processing


Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Stimuli
2.3. Procedure
2.4. Behavioral Data Analysis
2.5. EEG Preprocessing
2.6. Event-Related Potential Analysis
3. Results
3.1. Behavioral Results
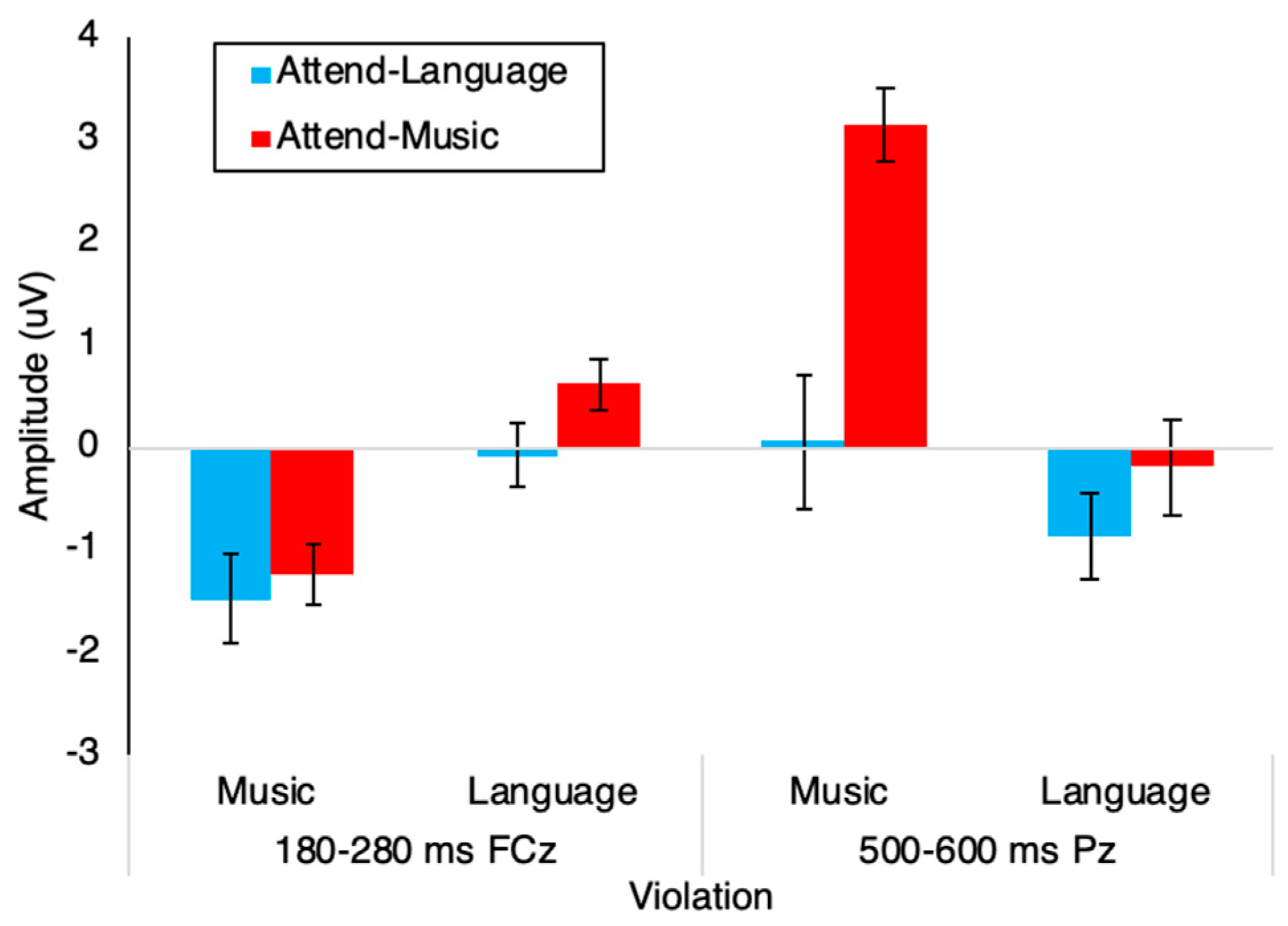
3.2. Event-Related Potentials
4. Discussion
5. Limitations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Patel, A.D. Language, music, syntax and the brain. Nat. Neurosci. 2003, 6, 674–681. [Google Scholar] [CrossRef]
- Slevc, L.R.; Rosenberg, J.C.; Patel, A.D. Making psycholinguistics musical: Self-paced reading time evidence for shared processing of linguistic and musical syntax. Psychon. Bull. Rev. 2009, 16, 374–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roncaglia-Denissen, M.P.; Bouwer, F.L.; Honing, H. Decision Making Strategy and the Simultaneous Processing of Syntactic Dependencies in Language and Music. Front. Psychol. 2018, 9, 38. [Google Scholar] [CrossRef] [PubMed]
- Jentschke, S.; Koelsch, S.; Friederici, A.D. Investigating the relationship of music and language in children: Influences of musical training and language impairment. Ann. N. Y. Acad. Sci. 2005, 1060, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Koelsch, S.; Gunter, T.C.; Wittfoth, M.; Sammler, D. Interaction between Syntax Processing in Language and in Music: An ERP Study. J. Cogn. Neurosci. 2005, 17, 1565–1577. [Google Scholar] [CrossRef]
- Fedorenko, E.; Patel, A.; Casasanto, D.; Winawer, J.; Gibson, E. Structural integration in language and music: Evidence for a shared system. Mem. Cogn. 2009, 37, 1–9. [Google Scholar] [CrossRef]
- Perruchet, P.; Poulin-Charronnat, B. Challenging prior evidence for a shared syntactic processor for language and music. Psychon. Bull. Rev. 2012, 20, 310–317. [Google Scholar] [CrossRef] [Green Version]
- Slevc, L.R.; Okada, B.M. Processing structure in language and music: A case for shared reliance on cognitive control. Psychon. Bull. Rev. 2015, 22, 637–652. [Google Scholar] [CrossRef]
- Posner, M.I.; Petersen, S.E. The Attention System of the Human Brain. Annu. Rev. Neurosci. 1990, 13, 25–42. [Google Scholar] [CrossRef]
- Coull, J.T.; Nobre, A.C. Where and When to Pay Attention: The Neural Systems for Directing Attention to Spatial Locations and to Time Intervals as Revealed by Both PET and fMRI. J. Neurosci. 1998, 18, 7426–7435. [Google Scholar] [CrossRef]
- Koelsch, S.; Schmidt, B.-H.; Kansok, J. Effects of musical expertise on the early right anterior negativity: An event-related brain potential study. Psychophysiology 2002, 39, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Koelsch, S.; Gunter, T.; Friederici, A.D.; Schröger, E. Brain Indices of Music Processing: “Nonmusicians” are Musical. J. Cogn. Neurosci. 2000, 12, 520–541. [Google Scholar] [CrossRef] [PubMed]
- Sammler, D.; Koelsch, S.; Ball, T.; Brandt, A.; Grigutsch, M.; Huppertz, H.-J.; Knösche, T.R.; Wellmer, J.; Widman, G.; Elger, C.E.; et al. Co-localizing linguistic and musical syntax with intracranial EEG. NeuroImage 2013, 64, 134–146. [Google Scholar] [CrossRef] [PubMed]
- Hahne, A.B.; Friederici, A.D. Electrophysiological evidence for two steps in syntactic analysis. Early automatic and late controlled processes. J. Cogn. Neurosci. 1999, 11, 194–205. [Google Scholar] [PubMed]
- Friederici, A.D. Towards a neural basis of auditory sentence processing. Trends Cogn. Sci. 2002, 6, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Neville, H.; Nicol, J.L.; Barss, A.; Forster, K.I.; Garrett, M.F. Syntactically Based Sentence Processing Classes: Evidence from Event-Related Brain Potentials. J. Cogn. Neurosci. 1991, 3, 151–165. [Google Scholar] [CrossRef]
- Maess, B.; Koelsch, S.; Gunter, T.C.; Friederici, A.D. Musical syntax is processed in Broca’s area: An MEG study. Nat. Neurosci. 2001, 4, 540–545. [Google Scholar] [CrossRef]
- Cheung, V.K.M.; Meyer, L.; Friederici, A.D.; Koelsch, S. The right inferior frontal gyrus processes nested non-local dependencies in music. Sci. Rep. 2018, 8, 3822. [Google Scholar] [CrossRef]
- Tillmann, B.; Koelsch, S.; Escoffier, N.; Bigand, E.; Lalitte, P.; Friederici, A.; Von Cramon, D. Cognitive priming in sung and instrumental music: Activation of inferior frontal cortex. NeuroImage 2006, 31, 1771–1782. [Google Scholar] [CrossRef]
- Bianco, R.; Novembre, G.; Keller, P.; Kim, S.-G.; Scharf, F.; Friederici, A.; Villringer, A.; Sammler, D.; Keller, P. Neural networks for harmonic structure in music perception and action. NeuroImage 2016, 142, 454–464. [Google Scholar] [CrossRef]
- Sammler, D.; Koelsch, S.; Friederici, A.D. Are left fronto-temporal brain areas a prerequisite for normal music-syntactic processing? Cortex 2011, 47, 659–673. [Google Scholar] [CrossRef] [PubMed]
- Jentschke, S.; Koelsch, S.; Sallat, S.; Friederici, A.D. Children with Specific Language Impairment Also Show Impairment of Music-syntactic Processing. J. Cogn. Neurosci. 2008, 20, 1940–1951. [Google Scholar] [CrossRef] [PubMed]
- Koelsch, S.; Vuust, P.; Friston, K. Predictive Processes and the Peculiar Case of Music. Trends Cogn. Sci. 2018, 23, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Loui, P.; Grent-’t-Jong, T.; Torpey, D.; Woldorff, M. Effects of attention on the neural processing of harmonic syntax in Western music. Brain Res. Cogn. Brain Res. 2005, 25, 678–687. [Google Scholar] [CrossRef]
- Maidhof, C.; Koelsch, S. Effects of Selective Attention on Syntax Processing in Music and Language. J. Cogn. Neurosci. 2011, 23, 2252–2267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutas, M.; Hillyard, S.A. Brain potentials during reading reflect word expectancy and semantic association. Nature 1984, 307, 161–163. [Google Scholar] [CrossRef]
- Kutas, M.; Hillyard, S. Reading senseless sentences: Brain potentials reflect semantic incongruity. Science 1980, 207, 203–205. [Google Scholar] [CrossRef]
- Kuperberg, G.R. Neural mechanisms of language comprehension: Challenges to syntax. Brain Res. 2007, 1146, 23–49. [Google Scholar] [CrossRef]
- Osterhout, L.; Holcomb, P.J.; Swinney, D.A. Brain potentials elicited by garden-path sentences: Evidence of the application of verb information during parsing. J. Exp. Psychol. Learn. Mem. Cogn. 1994, 20, 786–803. [Google Scholar] [CrossRef]
- Patel, A.D.; Gibson, E.; Ratner, J.; Besson, M.; Holcomb, P.J. Processing syntactic relations in language and music: An event-related potential study. J. Cogn. Neurosci. 1998, 10, 717–733. [Google Scholar] [CrossRef]
- Polich, J. Updating P300: An integrative theory of P3a and P3b. Clin. Neurophysiol. 2007, 118, 2128–2148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knight, R.T.; Grabowecky, M.F.; Scabini, D. Role of human prefrontal cortex in attention control. Adv. Neurol. 1995, 66, 21–36. [Google Scholar] [PubMed]
- Przysinda, E.; Zeng, T.; Maves, K.; Arkin, C.; Loui, P. Jazz musicians reveal role of expectancy in human creativity. Brain Cogn. 2017, 119, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Shipley, W.C. A Self-Administering Scale for Measuring Intellectual Impairment and Deterioration. J. Psychol. 1940, 9, 371–377. [Google Scholar] [CrossRef]
- Loui, P.; Guenther, F.H.; Mathys, C.; Schlaug, G. Action-perception mismatch in tone-deafness. Curr. Boil. 2008, 18, R331–R332. [Google Scholar] [CrossRef] [PubMed]
- Peretz, I.; Champod, A.S.; Hyde, K. Varieties of musical disorders. The Montreal Battery of Evaluation of Amusia. Ann. N. Y. Acad. Sci. 2003, 999, 58–75. [Google Scholar] [CrossRef] [PubMed]
- Zicarelli, D. An extensible real-time signal processing environment for Max. In Proceedings of the International Computer Music Conference, University of Michigan, Ann Arbor, MI, USA, 1–6 October 1998; Available online: https://quod.lib.umich.edu/i/icmc/bbp2372.1998.274/1 (accessed on 25 October 2019).
- Loui, P.; Wu, E.H.; Wessel, D.L.; Knight, R.T. A generalized mechanism for perception of pitch patterns. J. Neurosci. 2009, 29, 454–459. [Google Scholar] [CrossRef]
- Widmann, A.; Schröger, E.; Maess, B. Digital filter design for electrophysiological data—A practical approach. J. Neurosci. Methods 2015, 250, 34–46. [Google Scholar] [CrossRef]
- Hillyard, S.A.; Hink, R.F.; Schwent, V.L.; Picton, T.W. Electrical Signs of Selective Attention in the Human Brain. Science 1973, 182, 177–180. [Google Scholar] [CrossRef] [Green Version]
- Donchin, E.; Heffley, E.; Hillyard, S.A.; Loveless, N.; Maltzman, I.; Ohman, A.; Rosler, F.; Ruchkin, D.; Siddle, D. Cognition and event-related potentials. II. The orienting reflex and P300. Ann. N. Y. Acad. Sci. 1984, 425, 39–57. [Google Scholar] [CrossRef]
- Gray, J.A.; Wedderburn, A.A.I. Shorter articles and notes grouping strategies with simultaneous stimuli. Q. J. Exp. Psychol. 1960, 12, 180–184. [Google Scholar] [CrossRef]
- Deutsch, J.A.; Deutsch, D. Attention: Some theoretical considerations. Psychol. Rev. 1963, 70, 51–60. [Google Scholar] [CrossRef]
- Treisman, A.M.; Gelade, G. A feature-integration theory of attention. Cogn. Psychol. 1980, 12, 97–136. [Google Scholar] [CrossRef]
- Woldorff, M.G.; Hillyard, S.A. Modulation of early auditory processing during selective listening to rapidly presented tones. Electroencephalogr. Clin. Neurophysiol. 1991, 79, 170–191. [Google Scholar] [CrossRef]
- Woldorff, M.G.; Gallen, C.C.; Hampson, S.A.; Hillyard, S.A.; Pantev, C.; Sobel, D.; Bloom, F.E. Modulation of early sensory processing in human auditory cortex during auditory selective attention. Proc. Natl. Acad. Sci. USA 1993, 90, 8722–8726. [Google Scholar] [CrossRef]
- Näätänen, R.; Gaillard, A.; Mäntysalo, S. Early selective-attention effect on evoked potential reinterpreted. Acta Psychol. 1978, 42, 313–329. [Google Scholar] [CrossRef]
- Falkenstein, M.; Hohnsbein, J.; Hoormann, J.; Blanke, L. Effects of crossmodal divided attention on late ERP components. II. Error processing in choice reaction tasks. Electroencephalogr. Clin. Neurophysiol. 1991, 78, 447–455. [Google Scholar] [CrossRef]
- Large, E.W.; Jones, M.R. The dynamics of attending: How people track time-varying events. Psychol. Rev. 1999, 106, 119–159. [Google Scholar] [CrossRef]
- Treisman, A.M. Contextual cues in selective listening. Q. J. Exp. Psychol. 1960, 12, 242–248. [Google Scholar] [CrossRef]
- O’Connell, R.G.; Dockree, P.M.; Kelly, S.P. A supramodal accumulation-to-bound signal that determines perceptual decisions in humans. Nat. Neurosci. 2012, 15, 1729–1735. [Google Scholar] [CrossRef]
- Van Vugt, M.K.; Beulen, M.A.; Taatgen, N.A. Relation between centro-parietal positivity and diffusion model parameters in both perceptual and memory-based decision making. Brain Res. 2019, 1715, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, H.; Hoeks, J.C.J. A time and place for language comprehension: Mapping the N400 and the P600 to a minimal cortical network. Front. Hum. Neurosci. 2013, 7, 758. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, H.; Crocker, M.W. On the Proper Treatment of the N400 and P600 in Language Comprehension. Front. Psychol. 2017, 8, 1327. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, H.; Crocker, M.W.; Venhuizen, N.J.; Hoeks, J.C.J. A Neurocomputational Model of the N400 and the P600 in Language Processing. Cogn. Sci. 2017, 41 (Suppl. 6), 1318–1352. [Google Scholar] [CrossRef]
- Bianco, R.; Novembre, G.; Keller, P.E.; Scharf, F.; Friederici, A.D.; Villringer, A.; Sammler, D. Syntax in Action Has Priority over Movement Selection in Piano Playing: An ERP Study. J. Cogn. Neurosci. 2016, 28, 41–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagoort, P. Interplay between Syntax and Semantics during Sentence Comprehension: ERP Effects of Combining Syntactic and Semantic Violations. J. Cogn. Neurosci. 2003, 15, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Koelsch, S.; Kasper, E.; Sammler, D.; Schulze, K.; Gunter, T.; Friederici, A.D. Music, language and meaning: Brain signatures of semantic processing. Nat. Neurosci. 2004, 7, 302–307. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variable | Attend-Language (N = 16) | Attend-Music (N = 19) |
|---|---|---|
| Age in years, M (SD) | 19.625 (2.029) | 19.389 (2.033) |
| Male, n | 11/16 | 8/19 |
| Music Training, years, M (SD) | 2.233 (3.422) | 3.105 (4.012) |
| Musically trained, n | 9/16 | 11/18 |
| Full-Scale IQ (Estimated from Shipley-Hartford IQ scale, M (SD)) | 100 (10) | 101 (7) |
| MBEA, M(SD) | 23.375 (3.828) | 25.11 (2.685) |
| Pitch Discrimination, ΔHz/500 Hz, M (SD) | 12.469 (11.895) | 11.087 (7.174) |
| Normal Hearing, % | 100% | 100% |
| English as First Language, n | 15/16 | 12/18 |
| Tests of Within-Subjects Contrasts | |||||
| Source | Time-Window | df | F | p | Partial eta2 |
| Violation | 180–280 ms | 1 | 33.198 | < 0.001 | 0.501 |
| 500–600 ms | 1 | 31.317 | < 0.001 | 0.487 | |
| Violation * Attend | 180–280 ms | 1 | 0.64 | 0.43 | 0.019 |
| 500–600 ms | 1 | 9.951 | 0.003 | 0.232 | |
| Tests of Between-Subjects Effects | |||||
| Source | Time-Window | df | F | p | Partial eta2 |
| Attend | 180–280 ms | 1 | 1.381 | 0.248 | 0.040 |
| 500–600 ms | 1 | 9.763 | 0.004 | 0.228 | |
| First Language English Speakers Only | Tests of Within-Subjects Contrasts | ||||
| Source | Time-Window | df | F | p | Partial eta2 |
| Violation | 180–280 ms | 1 | 14.216 | < 0.001 | 0.428 |
| 500–600 ms | 1 | 30.722 | < 0.001 | 0.618 | |
| Violation * Attend | 180–280 ms | 1 | 0.195 | 0.664 | 0.01 |
| 500–600 ms | 1 | 11.075 | 0.004 | 0.368 | |
| Tests of Between-Subjects Effects | |||||
| Source | Time-Window | df | F | p | Partial eta2 |
| Attend | 180–280 ms | 1 | 0.971 | 0.337 | 0.049 |
| 500–600 ms | 1 | 13.99 | < 0.001 | 0.424 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.J.; Jung, H.; Loui, P. Attention Modulates Electrophysiological Responses to Simultaneous Music and Language Syntax Processing. Brain Sci. 2019, 9, 305. https://doi.org/10.3390/brainsci9110305
Lee DJ, Jung H, Loui P. Attention Modulates Electrophysiological Responses to Simultaneous Music and Language Syntax Processing. Brain Sciences. 2019; 9(11):305. https://doi.org/10.3390/brainsci9110305
Chicago/Turabian StyleLee, Daniel J., Harim Jung, and Psyche Loui. 2019. "Attention Modulates Electrophysiological Responses to Simultaneous Music and Language Syntax Processing" Brain Sciences 9, no. 11: 305. https://doi.org/10.3390/brainsci9110305
APA StyleLee, D. J., Jung, H., & Loui, P. (2019). Attention Modulates Electrophysiological Responses to Simultaneous Music and Language Syntax Processing. Brain Sciences, 9(11), 305. https://doi.org/10.3390/brainsci9110305