The Influence of Subclinical Neck Pain on Neurophysiological and Behavioral Measures of Multisensory Integration



Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Procedure
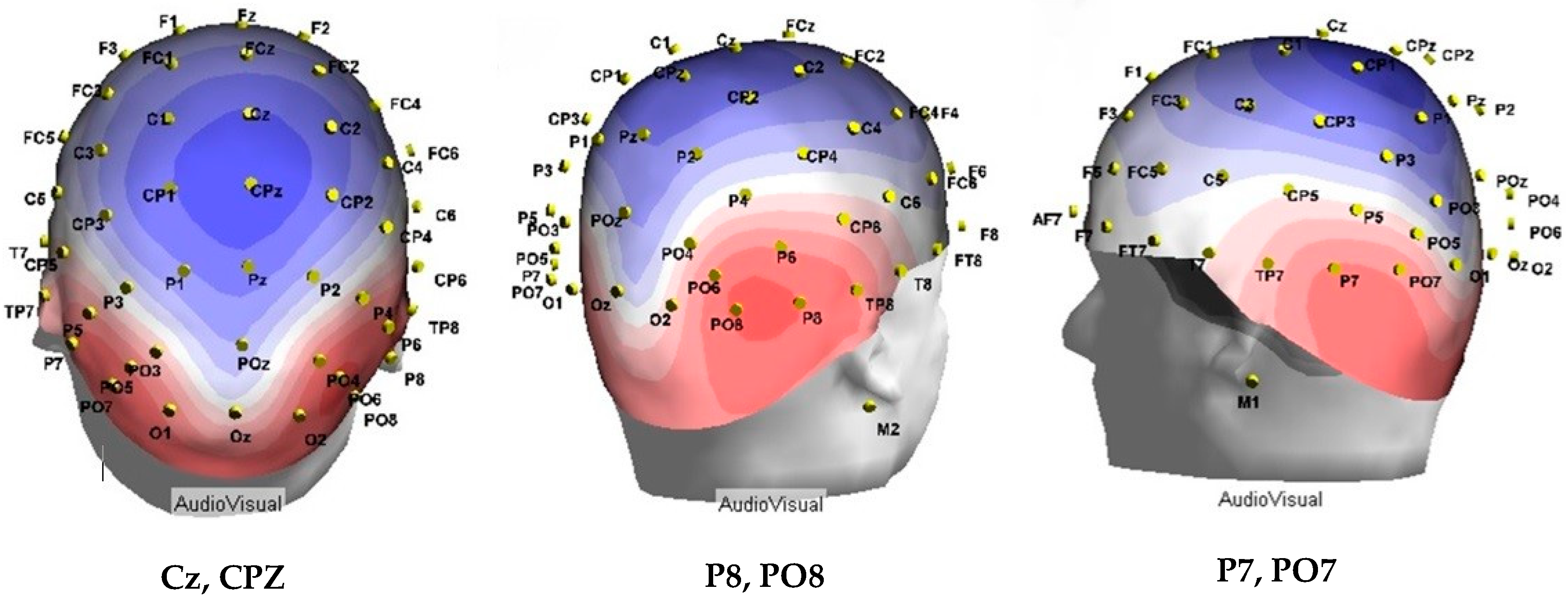
2.2.1. Electroencephalography Set-up and Acquisition
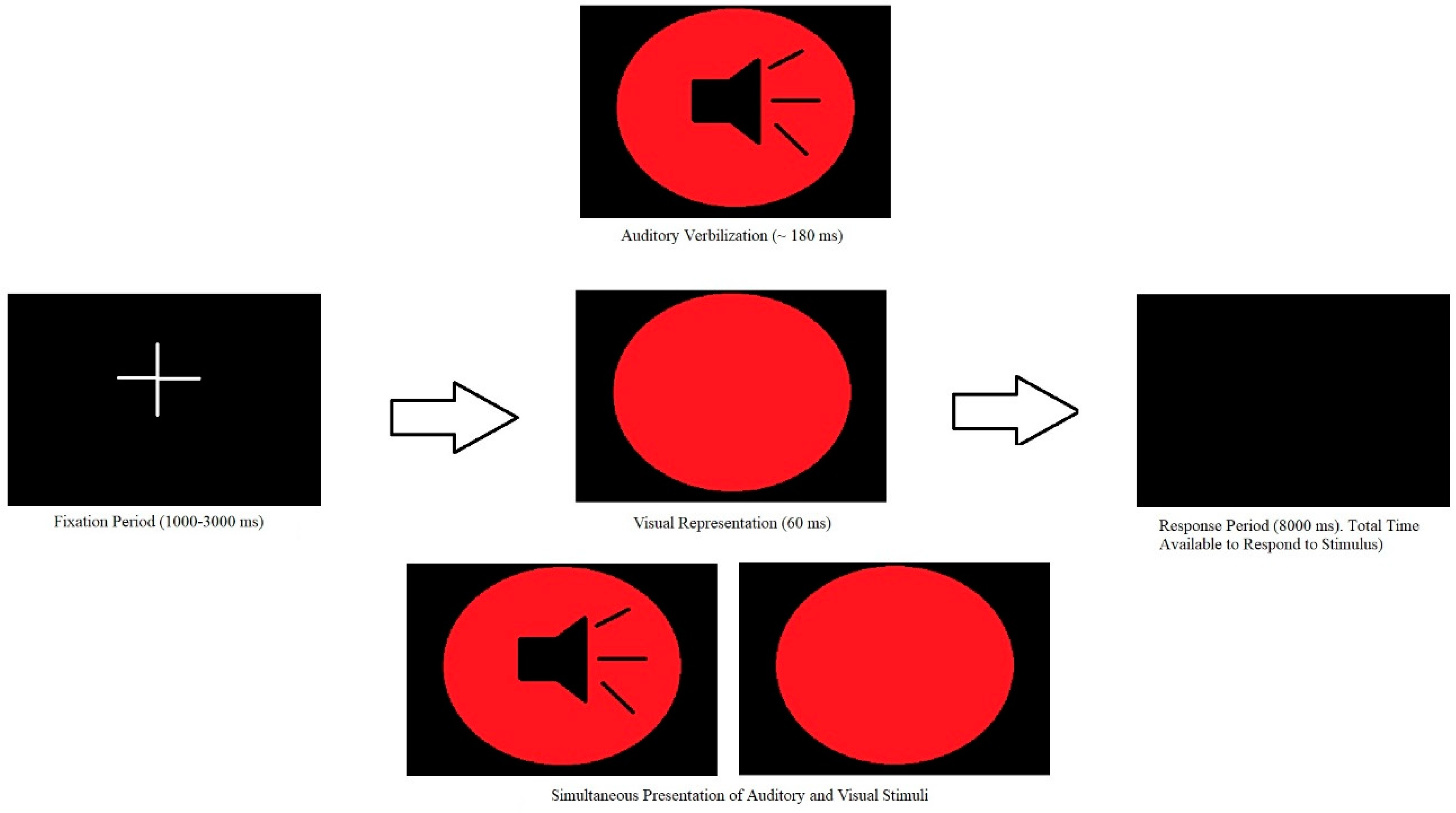
2.2.2. Simple Reaction Time Multisensory Integration Task
2.3. BehaviouralAnalysis
2.3.1. Task Performance Analysis
2.3.2. The Race Model
2.4. Neurophysiological Analysis
2.4.1. Artifact Removal
2.4.2. EEG/Epochs Analysis Method 1
2.4.3. EEG/Epochs Analysis Method 2
3. Results
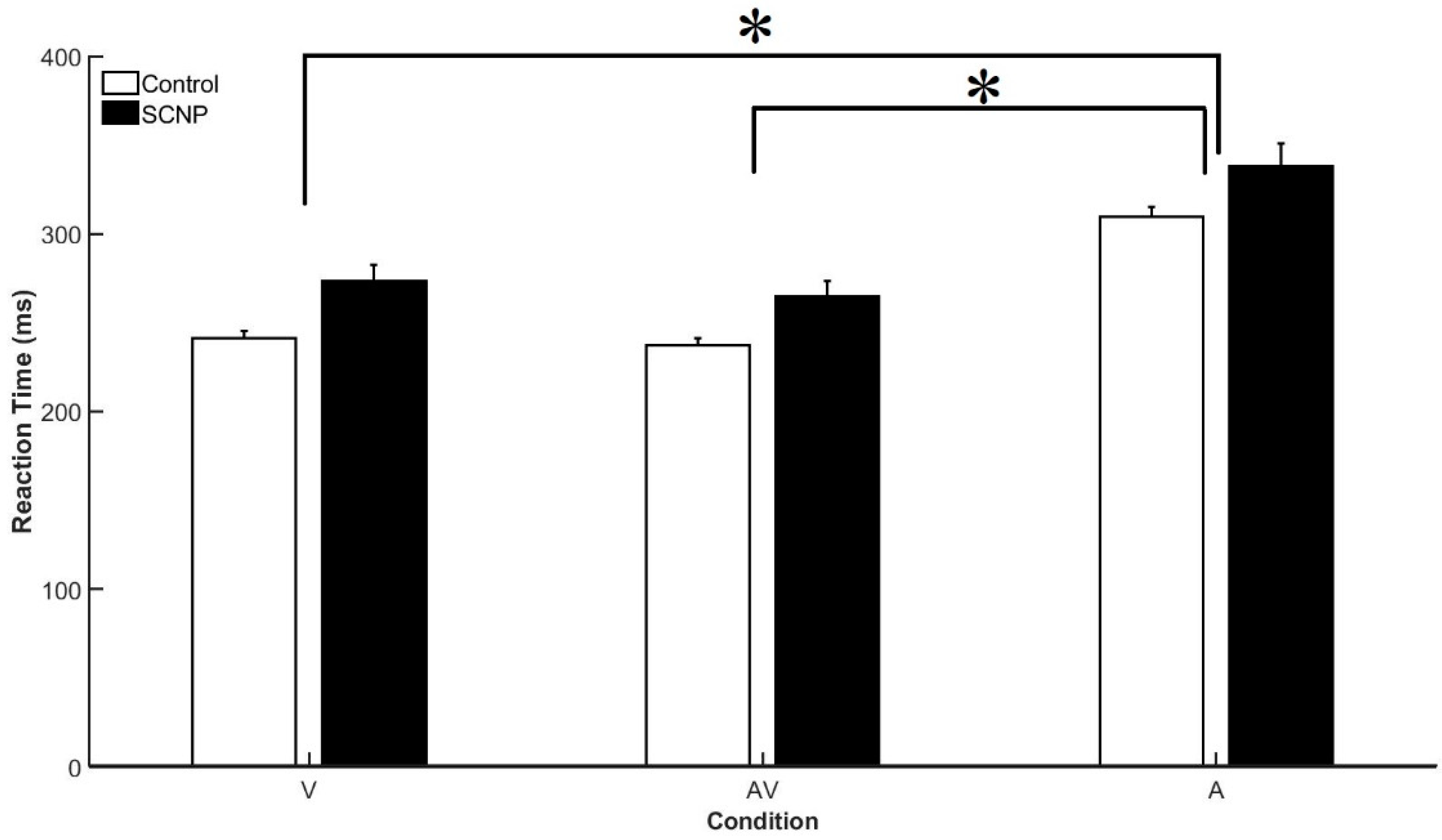
3.1. Behavioral Data
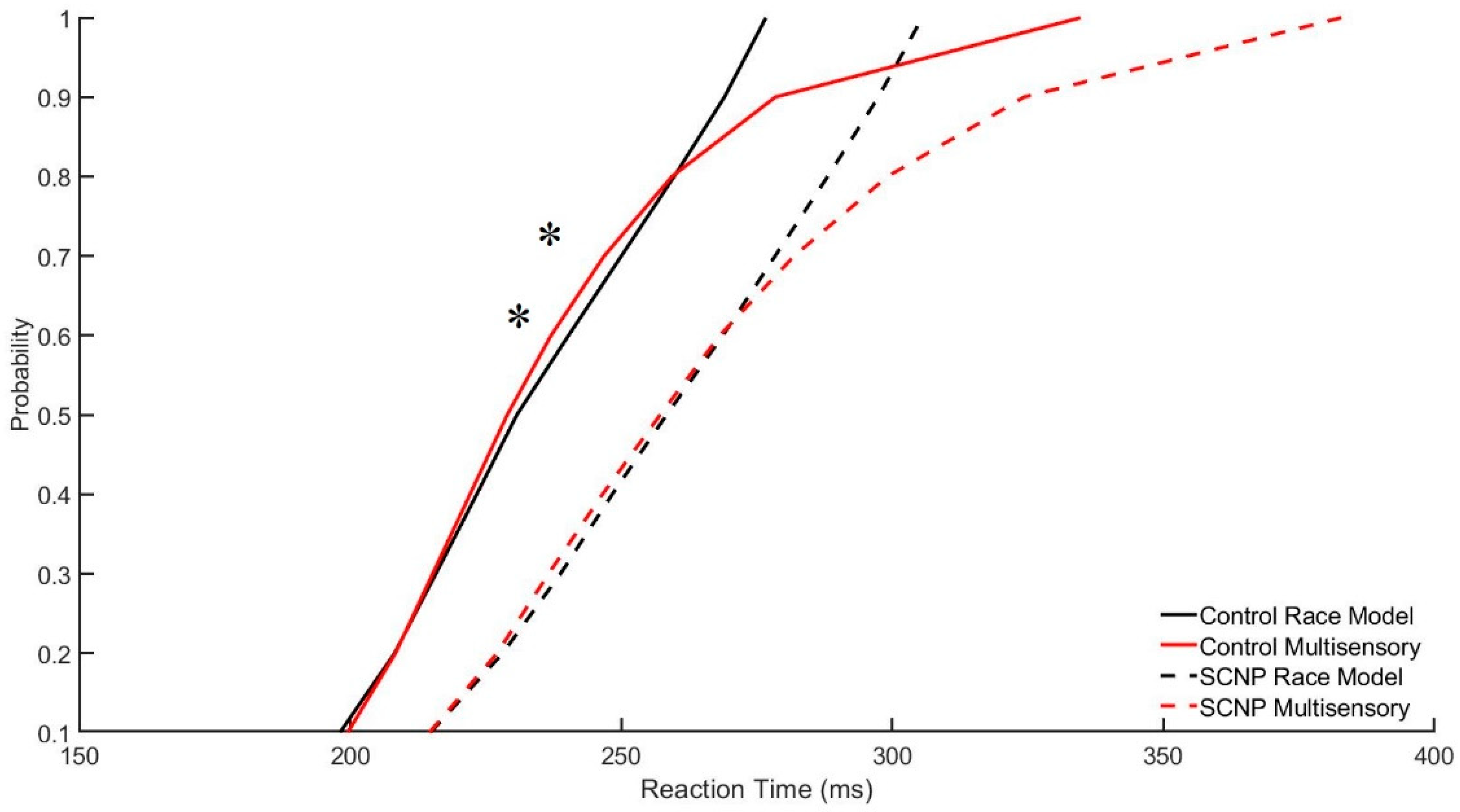
Race Model
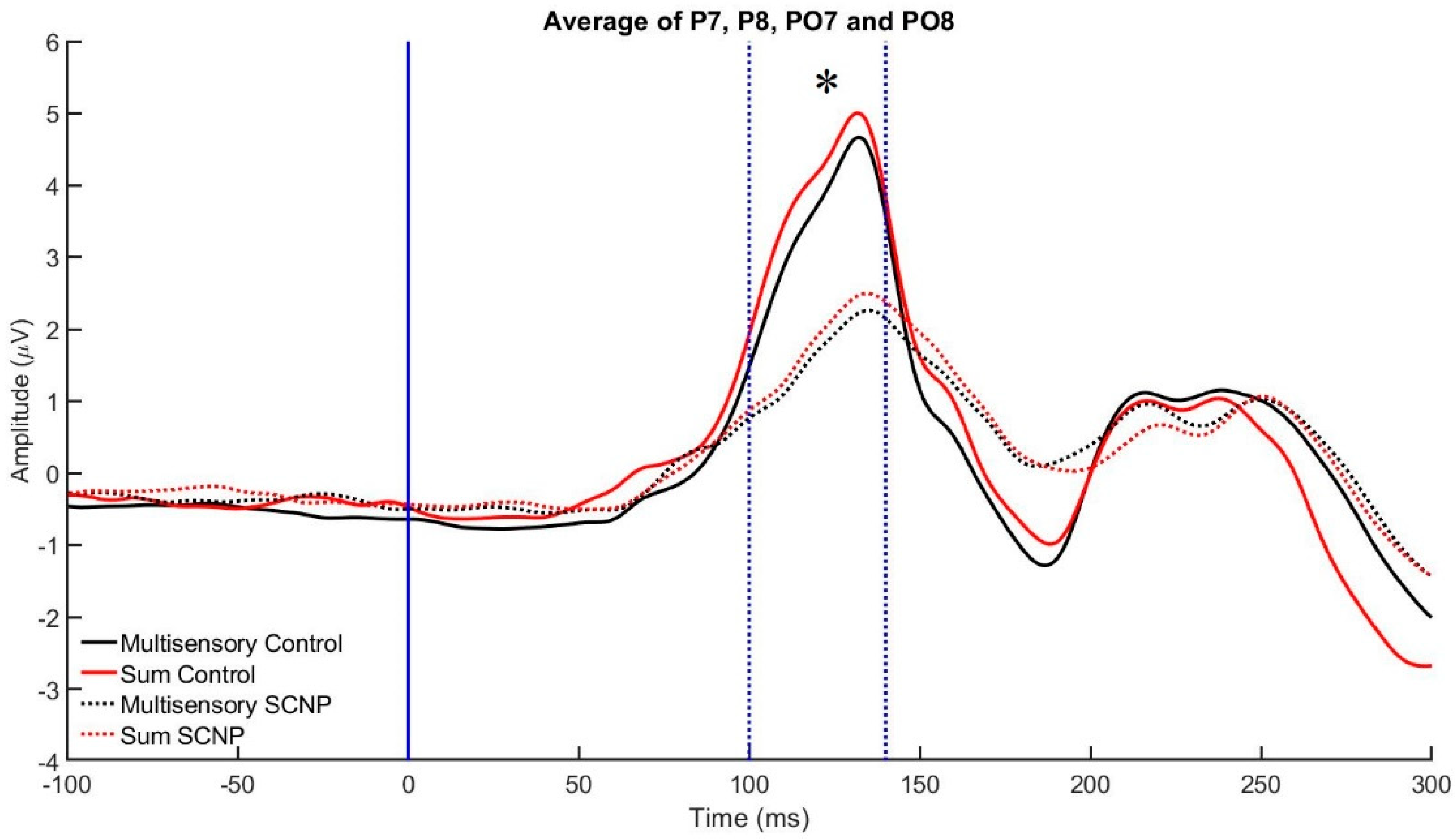
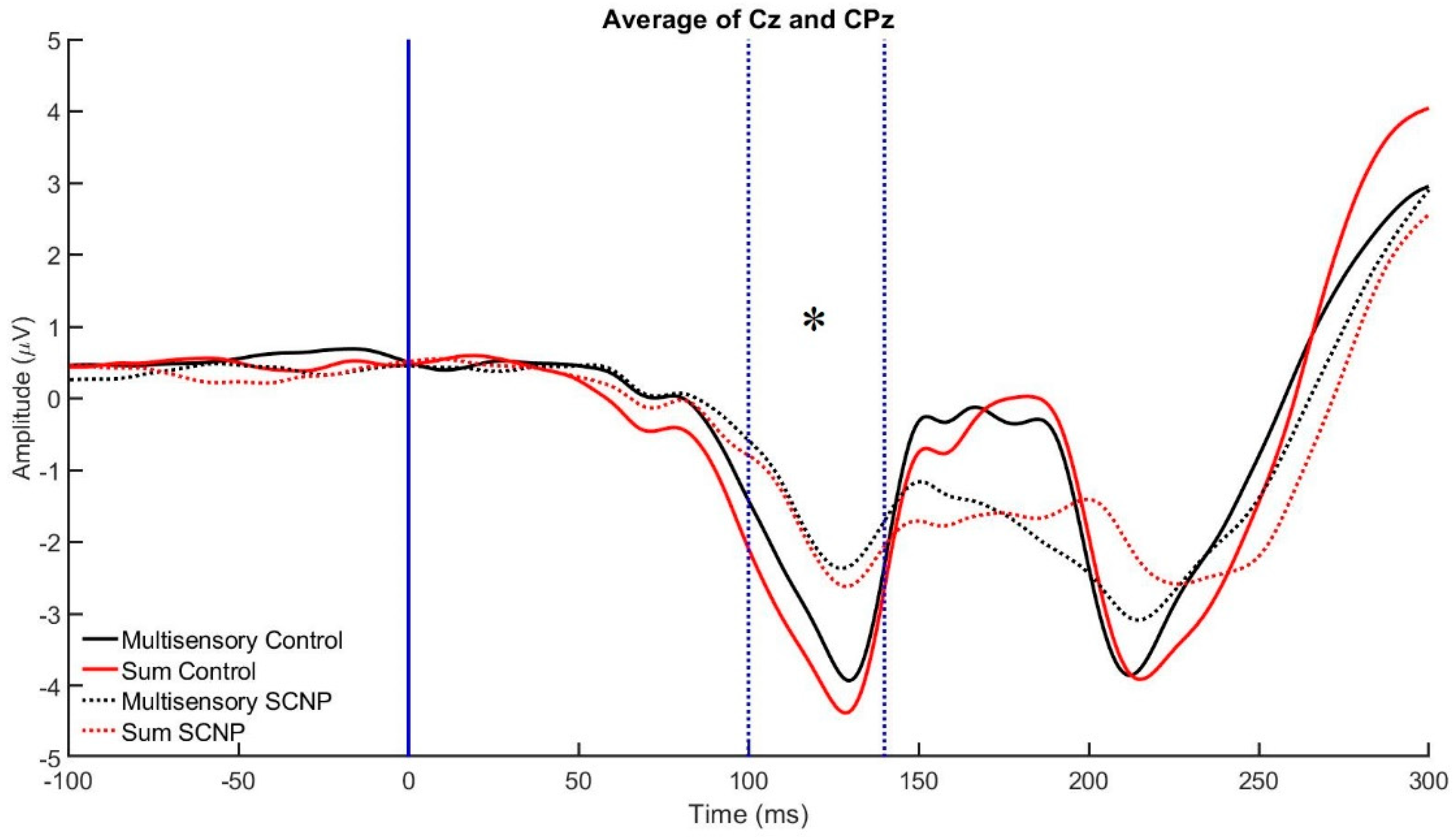
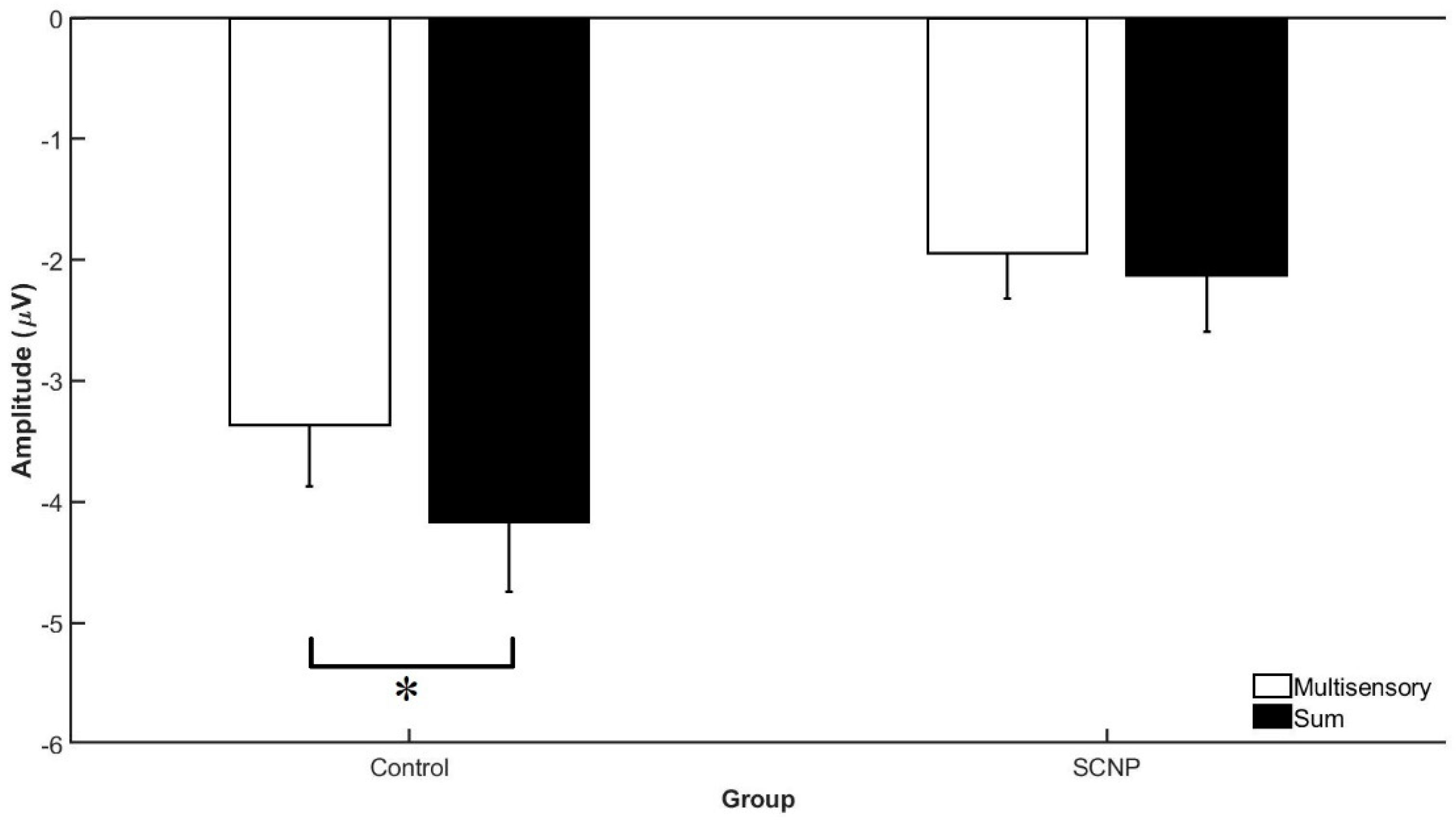
3.2. Electrophysiological Data
4. Discussion
4.1. Behavioral Findings
4.2. Electrophysiological Findings
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ariëns, G.A.M.; van Mechelen, W.; Bongers, P.M.; Bouter, L.M.; van der Wal, G. Physical risk factors for neck pain. Scand. J. Work. Environ. Health 2000, 26, 7–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariëns, G.A.M.; Bongers, P.M.; Douwes, M.; Miedema, M.C.; Hoogendoorn, W.E.; van der Wal, G.; Bouter, L.M.; van Mechelen, W. Are neck flexion, neck rotation, and sitting at work risk factors for neck pain? Results of a prospective cohort study. Occup. Environ. Med. 2001, 58, 200–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogg-Johnson, S.; van der Velde, G.; Carroll, L.J.; Holm, L.W.; Cassidy, J.D.; Guzman, J.; Côté, P.; Haldeman, S.; Ammendolia, C.; Carragee, E.; et al. The Burden and Determinants of Neck Pain in the General Population. Eur. Spine J. 2008, 17, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Nicholson, L.L.; Adams, R.D.; Bae, S.S. Proprioception and rotation range sensitization associated with subclinical neck pain. Spine 2005, 30, E60–E67. [Google Scholar] [CrossRef] [PubMed]
- Zabihhosseinian, M.; Holmes, M.W.R.; Ferguson, B.; Murphy, B. Neck muscle fatigue alters the cervical flexion relaxation ratio in sub-clinical neck pain patients. Clin. Biomech. 2015, 30, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Haavik, H.; Murphy, B. Subclinical neck pain and the effects of cervical manipulation on elbow joint position sense. J. Manip. Physiol. Ther. 2011, 34, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Abbruzzese, G.; Berardelli, A. Sensorimotor integration in movement disorders. Mov. Disord. 2003, 18, 231–240. [Google Scholar] [CrossRef]
- Haavik, H.; Murphy, B. The role of spinal manipulation in addressing disordered sensorimotor integration and altered motor control. J. Electromyogr. Kinesiol. 2012, 22, 768–776. [Google Scholar] [CrossRef]
- Haavik, T.; Murphy, B.A. Altered cortical integration of dual somatosensory input following the cessation of a 20 min period of repetitive muscle activity. Exp. Brain Res. 2007, 178, 488–498. [Google Scholar] [CrossRef]
- Dancey, E.; Murphy, B.; Andrew, D.; Yielder, P. The interactive effect of acute pain and motor learning acquisition on sensorimotor integration and motor learning outcomes. J. Neurophysiol. 2016, 116, 2210–2220. [Google Scholar] [CrossRef] [Green Version]
- Boudreau, S.; Romaniello, A.; Wang, K.; Svensson, P.; Sessle, B.J.; Arendt-Nielsen, L. The effects of intra-oral pain on motor cortex neuroplasticity associated with short-term novel tongue-protrusion training in humans. Pain 2007, 132, 169. [Google Scholar] [CrossRef] [PubMed]
- Andrew, D.; Yielder, P.; Haavik, H.; Murphy, B. The effects of subclinical neck pain on sensorimotor integration following a complex motor pursuit task. Exp. Brain Res. 2018, 236, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Baarbe, J.K.; Holmes, M.W.R.; Murphy, H.E.; Haavik, H.; Murphy, B.A. Influence of subclinical neck pain on the ability to perform a mental rotation task: A 4-week longitudinal study with a healthy control group comparison. J. Manip. Physiol. Ther. 2016, 39, 23–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiba, R.; Takakusaki, K.; Ota, J.; Yozu, A.; Haga, N. Human upright posture control models based on multisensory inputs; in fast and slow dynamics. Neurosci. Res. 2016, 104, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Murray, M.M.; Foxe, J.J.; Higgins, B.A.; Javitt, D.C.; Schroeder, C.E. Visuo-spatial neural response interactions in early cortical processing during a simple reaction time task: A high-density electrical mapping study. Neuropsychologia 2001, 39, 828–844. [Google Scholar] [CrossRef]
- Sugihara, T.; Diltz, M.D.; Averbeck, B.B.; Romanski, L.M. Integration of Auditory and Visual Communication Information in the Primate Ventrolateral Prefrontal Cortex. J. Neurosci. 2006, 26, 11138. [Google Scholar] [CrossRef] [Green Version]
- Horak, F.B. Postural orientation and equilibrium: What do we need to know about neural control of balance to prevent falls? Age Ageing 2006, 35, ii7–ii11. [Google Scholar] [CrossRef] [Green Version]
- Meredith, M.A.; Stein, B.E. Interactions among converging sensory inputs in the superior colliculus. Science 1983, 221, 389. [Google Scholar] [CrossRef] [Green Version]
- Murray, M.M.; Wallace, M.T. The Neural Bases of Multisensory Processes; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Stein, B.E.; Stanford, T.R. Multisensory integration: Current issues from the perspective of the single neuron. Nat. Rev. Neurosci. 2008, 9, 255–266. [Google Scholar] [CrossRef]
- King, A.J. The superior colliculus. Curr. Biol. 2004, 14, R335–R338. [Google Scholar] [CrossRef] [Green Version]
- Perrault, J.; Vaughan, J.W.; Stein, B.E.; Wallace, M.T. Superior Colliculus Neurons Use Distinct Operational Modes in the Integration of Multisensory Stimuli. J. Neurophysiol. 2005, 93, 2575–2586. [Google Scholar] [CrossRef] [PubMed]
- Brandwein, A.B.; Foxe, J.J.; Butler, J.S.; Russo, N.N.; Altschuler, T.S.; Gomes, H.; Molholm, S. The Development of Multisensory Integration in High-Functioning Autism: High-Density Electrical Mapping and Psychophysical Measures Reveal Impairments in the Processing of Audiovisual Inputs. Cereb. Cortex 2013, 23, 1329–1341. [Google Scholar] [CrossRef] [PubMed]
- Brandwein, A.B.; Foxe, J.J.; Russo, N.N.; Altschuler, T.S.; Gomes, H.; Molholm, S. The Development of Audiovisual Multisensory Integration Across Childhood and Early Adolescence: A High-Density Electrical Mapping Study. Cereb. Cortex 2011, 21, 1042–1055. [Google Scholar] [CrossRef] [PubMed]
- Molholm, S.; Sehatpour, P.; Mehta, A.D.; Shpaner, M.; Gomez-Ramirez, M.; Ortigue, S.; Dyke, J.P.; Schwartz, T.H.; Foxe, J.J. Audio-Visual Multisensory Integration in Superior Parietal Lobule Revealed by Human Intracranial Recordings. J. Neurophysiol. 2006, 96, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Eikema, D.J.A.; Hatzitaki, V.; Tzovaras, D.; Papaxanthis, C. Application of intermittent galvanic vestibular stimulation reveals age-related constraints in the multisensory reweighting of posture. Neurosci. Lett. 2014, 561, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Hairston, W.D.; Laurienti, P.J.; Mishra, G.; Burdette, J.H.; Wallace, M.T. Multisensory enhancement of localization under conditions of induced myopia. Exp. Brain Res. 2003, 152, 404–408. [Google Scholar] [CrossRef]
- Farid, B.; Yielder, P.; Holmes, M.; Haavik, H.; Murphy, B.A. Association of Subclinical Neck Pain With Altered Multisensory Integration at Baseline and 4-Week Follow-up Relative to Asymptomatic Controls. J. Manip. Physiol. Ther. 2018, 41, 81–91. [Google Scholar] [CrossRef]
- Holt, K.R.; Haavik, H.; Lee, A.C.L.; Murphy, B.; Elley, C.R. Effectiveness of chiropractic care to improve sensorimotor function associated with falls risk in older people: A randomized controlled trial. J. Manip. Physiol. Ther. 2016, 39, 267–278. [Google Scholar] [CrossRef] [Green Version]
- Laurienti, P.J.; Burdette, J.H.; Maldjian, J.A.; Wallace, M.T. Enhanced multisensory integration in older adults. Neurobiol. Aging 2004, 27, 1155–1163. [Google Scholar] [CrossRef]
- Miller, J. Divided attention: Evidence for coactivation with redundant signals. Cogn. Psychol. 1982, 14, 247–279. [Google Scholar] [CrossRef]
- Narjarian, K.; Splinter, R. Biomedical Signal and Image Processing, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- McCracken, H.S.; Murphy, B.A.; Glazebrook, C.M.; Burkitt, J.J.; Karellas, A.M.; Yielder, P.C. Audiovisual multisensory integration and evoked potentials in young adults with and without Attention-Deficit/Hyperactivity Disorder. Front. Hum. Neurosci. 2019, 13, 95. [Google Scholar] [CrossRef] [PubMed]
- Foxe, J.J.; Morocz, I.A.; Murray, M.M.; Higgins, B.A.; Javitt, D.C.; Schroeder, C.E. Multisensory auditory–somatosensory interactions in early cortical processing revealed by high-density electrical mapping. Cogn. Brain Res. 2000, 10, 77–83. [Google Scholar] [CrossRef]
- Giard, M.H.; Peronnet, F. Auditory-Visual Integration during Multimodal Object Recognition in Humans: A Behavioral and Electrophysiological Study. J. Cogn. Neurosci. 1999, 11, 473–490. [Google Scholar] [CrossRef] [PubMed]
- Molholm, S.; Ritter, W.; Murray, M.M.; Javitt, D.C.; Schroeder, C.E.; Foxe, J.J. Multisensory auditory–visual interactions during early sensory processing in humans: A high-density electrical mapping study. Cogn. Brain Res. 2002, 14, 115–128. [Google Scholar] [CrossRef]
- Stevenson, R.A.; Ghose, D.; Fister, J.K.; Sarko, D.K.; Altieri, N.A.; Nidiffer, A.R.; Kurela, L.R.; Siemann, J.K.; James, T.W.; Wallace, M.T. Identifying and Quantifying Multisensory Integration: A Tutorial Review. Brain Topogr. 2014, 27, 707–730. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, T.F.; Peterson, D.H. Chiropractic Technique: Principles and Procedures; Mosby Incorporated: Maryland, MO, USA, 2011. [Google Scholar]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Vernon, H.; Mior, S. The Neck Disability Index: A study of reliability and validity. J. Manip. Physiol. Ther. 1991, 14, 409–415. [Google Scholar]
- Von Korff, M.; Ormel, J.; Keefe, F.J.; Dworkin, S.F. Grading the severity of chronic pain. Pain 1992, 50, 133–149. [Google Scholar] [CrossRef]
- Teplan, M. Fundamentals of EEG measurement. Meas. Sci. Rev. 2002, 2, 1–11. [Google Scholar]
- Mantri, S.; Dukare, V.; Yeole, S.; Patil, D.; Wadhai, V.M. A Survey: Fundamental of EEG. Int. J. Adv. Res. Comput. Sci. Manag. Stud. 2013, 1, 83–89. [Google Scholar]
- Teder-Sälejärvi, W.A.; McDonald, J.J.; Di Russo, F.; Hillyard, S.A. An analysis of audio-visual crossmodal integration by means of event-related potential (ERP) recordings. Cogn. Brain Res. 2002, 14, 106–114. [Google Scholar] [CrossRef]
- Ulrich, R.; Miller, J.; Schröter, H. Testing the race model inequality: An algorithm and computer programs. Behav. Res. Methods 2007, 39, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Natalie, R.; John, F.J.; Brandwein, B.A.; Ted, A.; Hilary, G.; Sophie, M. Multisensory processing in children with autism: High-density electrical mapping of auditory—Somatosensory integration. Autism Res. 2010, 3, 253–267. [Google Scholar] [CrossRef]
- Guthrie, D.; Buchwald, J.S. Significance Testing of Difference Potentials. Psychophysiology 1991, 28, 240–244. [Google Scholar] [CrossRef] [PubMed]
- McCracken, H.; Murphy, B.; Burkitt, J.; Glazebrook, C.; Yielder, P. Audiovisual Multisensory Processing in Young Adults with Attention-Deficit/Hyperactivity Disorder. Multisens. Res. 2019, in press. [Google Scholar]
- Talsma, D.; Doty, T.J.; Woldorff, M.G. Selective Attention and Audiovisual Integration: Is Attending to Both Modalities a Prerequisite for Early Integration? Cereb. Cortex 2007, 17, 679–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lelic, D.; Niazi, I.K.; Holt, K.; Jochumsen, M.; Dremstrup, K.; Yielder, P.; Murphy, B.; Drewes, A.M.; Haavik, H. Manipulation of Dysfunctional Spinal Joints Affects Sensorimotor Integration in the Prefrontal Cortex: A Brain Source Localization Study. Neural. Plast. 2016, 9. [Google Scholar] [CrossRef] [Green Version]
- Benjamin, S.; Pandya, D.N. Parietal, temporal, and occipita projections to cortex of the superior temporal sulcus in the rhesus monkey: A retrograde tracer study. J. Comp. Neurol. 1994, 343, 445–463. [Google Scholar] [CrossRef]
- Fogassi, L.; Gallese, V.; Fadiga, L.; Luppino, G.; Matelli, M.; Rizzolatti, G. Coding of peripersonal space in inferior premotor cortex (area F4). J. Neurophysiol. 1996, 76, 141–157. [Google Scholar] [CrossRef]
- Gaffan, D.; Harrison, S. Auditory-visual associations, hemispheric specialization and temporal-frontal interaction in the rhesus monkey. Brain 1991, 114, 2133–2144. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Bin (ms) | Positive Voltage (μV) | Negative Voltage (μV) |
|---|---|---|
| 70–90 | N/A | POz, Oz |
| 100–140 | P7, PO7, P8, PO8 | CPz, Cz |
| 150–150 | Oz | N/A |
| 180–200 | Oz | N/A |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karellas, A.M.; Yielder, P.; Burkitt, J.J.; McCracken, H.S.; Murphy, B.A. The Influence of Subclinical Neck Pain on Neurophysiological and Behavioral Measures of Multisensory Integration. Brain Sci. 2019, 9, 362. https://doi.org/10.3390/brainsci9120362
Karellas AM, Yielder P, Burkitt JJ, McCracken HS, Murphy BA. The Influence of Subclinical Neck Pain on Neurophysiological and Behavioral Measures of Multisensory Integration. Brain Sciences. 2019; 9(12):362. https://doi.org/10.3390/brainsci9120362
Chicago/Turabian StyleKarellas, Antonia M., Paul Yielder, James J. Burkitt, Heather S. McCracken, and Bernadette A. Murphy. 2019. "The Influence of Subclinical Neck Pain on Neurophysiological and Behavioral Measures of Multisensory Integration" Brain Sciences 9, no. 12: 362. https://doi.org/10.3390/brainsci9120362
APA StyleKarellas, A. M., Yielder, P., Burkitt, J. J., McCracken, H. S., & Murphy, B. A. (2019). The Influence of Subclinical Neck Pain on Neurophysiological and Behavioral Measures of Multisensory Integration. Brain Sciences, 9(12), 362. https://doi.org/10.3390/brainsci9120362