Reliability of Fronto–Amygdala Coupling during Emotional Face Processing


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Protocol
2.3. Image Acquisition and Analysis
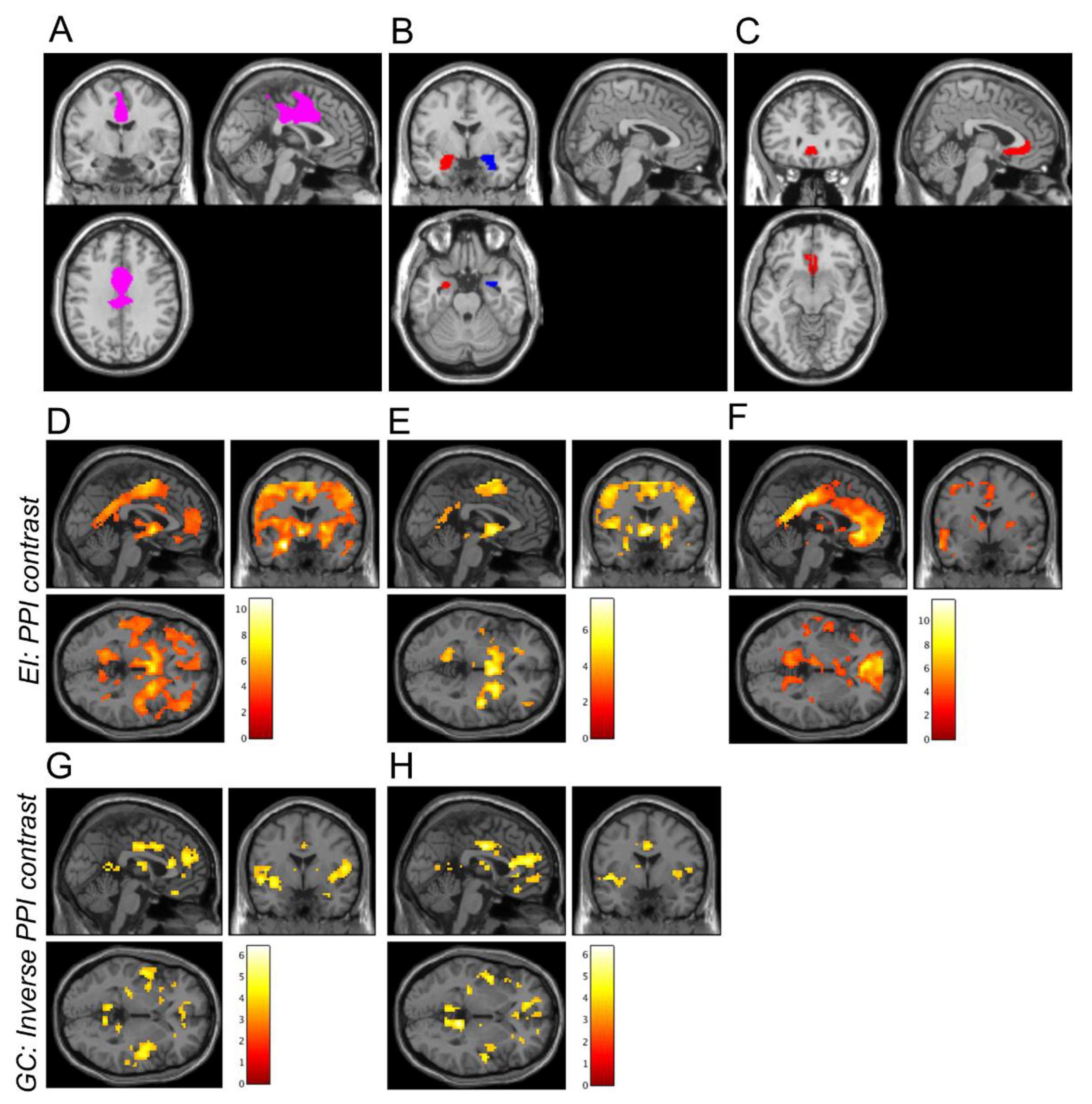
2.4. Psychophysiological Interaction Analysis
2.5. Reliability Testing
2.6. Power Analysis
3. Results
3.1. PPI Analysis
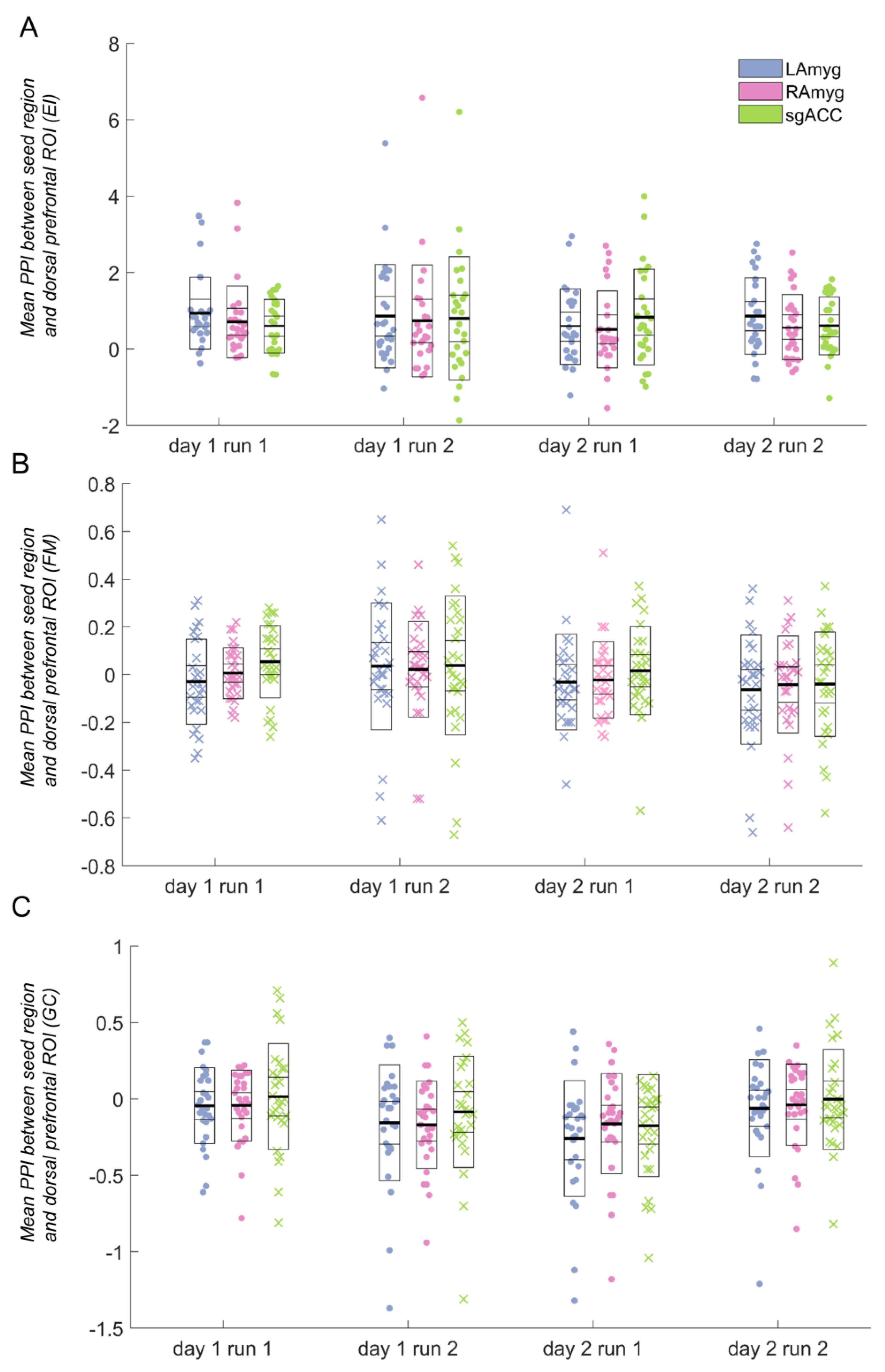
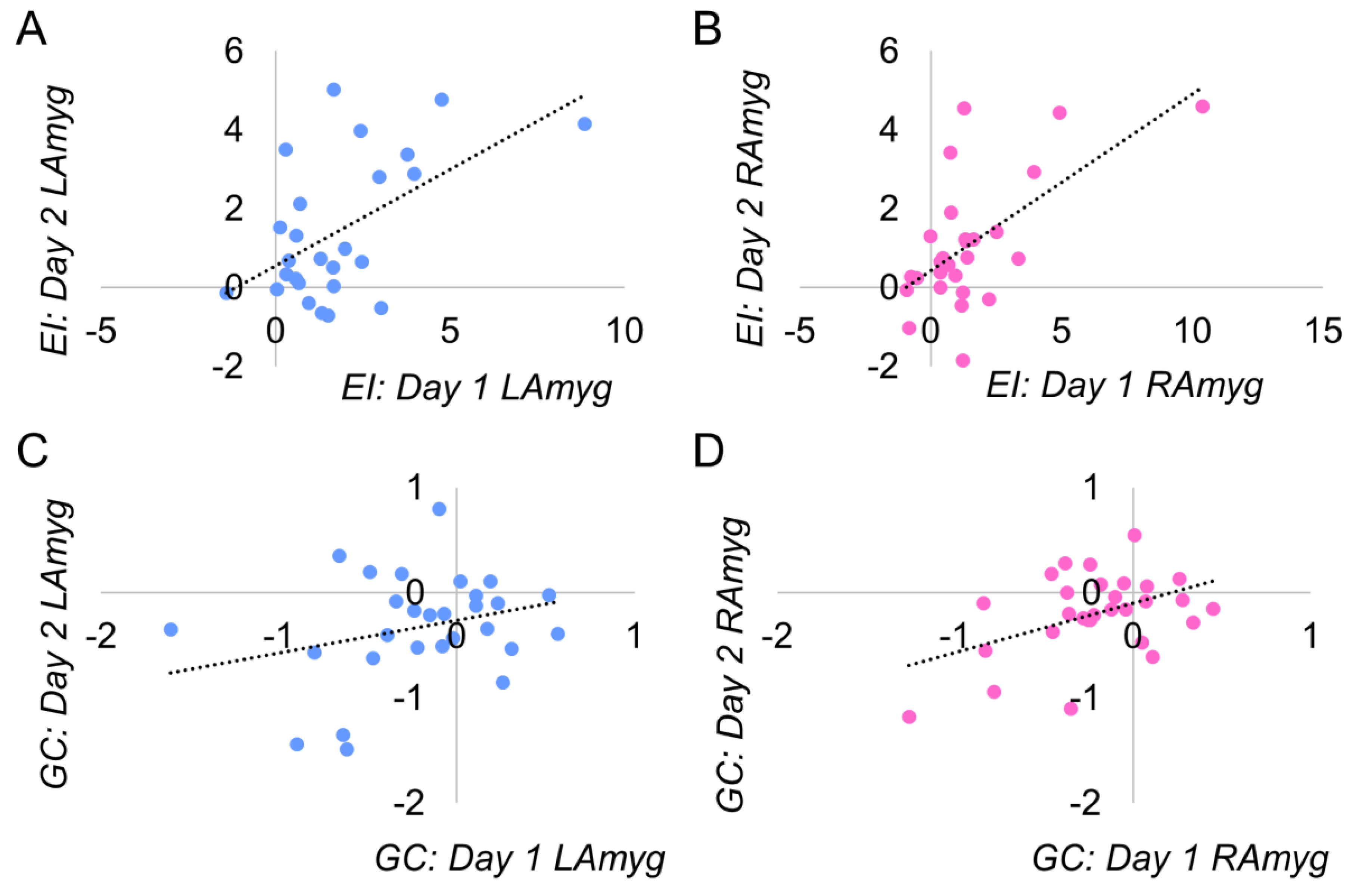
3.2. Reliability
3.3. Emotion Identification
3.4. Gender Classification
4. Discussion
Limitations and Future Directions
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nord, C.L.; Gray, A.; Charpentier, C.J.; Robinson, O.J.; Roiser, J.P. Unreliability of putative fMRI biomarkers during emotional face processing. Neuroimage 2017, 156, 119–127. [Google Scholar] [CrossRef]
- Plichta, M.M.; Schwarz, A.J.; Grimm, O.; Morgen, K.; Mier, D.; Haddad, L.; Gerdes, A.B.; Sauer, C.; Tost, H.; Esslinger, C.; et al. Test–retest reliability of evoked BOLD signals from a cognitive–emotive fMRI test battery. Neuroimage 2012, 60, 1746–1758. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, T.; Somerville, L.H.; Alexander, A.L.; Oakes, T.R.; Davidson, R.J.; Kalin, N.H.; Whalen, P.J. Stability of amygdala BOLD response to fearful faces over multiple scan sessions. Neuroimage 2005, 25, 1112–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manuck, S.B.; Brown, S.M.; Forbes, E.E.; Hariri, A.R. Temporal stability of individual differences in amygdala reactivity. Am. J. Psychiat. 2007, 164, 1613–1614. [Google Scholar] [CrossRef] [PubMed]
- Salvadore, G.; Cornwell, B.R.; Sambataro, F.; Latov, D.; Colon-Rosario, V.; Carver, F.; Holroyd, T.; DiazGranados, N.; Machado-Vieira, R.; Grillon, C.; et al. Anterior cingulate desynchronization and functional connectivity with the amygdala during a working memory task predict rapid antidepressant response to ketamine. Neuropsychopharmacology 2010, 35, 1415. [Google Scholar] [CrossRef] [PubMed]
- Sarpal, D.K.; Argyelan, M.; Robinson, D.G.; Szeszko, P.R.; Karlsgodt, K.H.; John, M.; Weissman, N.; Gallego, J.A.; Kane, J.M.; Lencz, T.; et al. Baseline striatal functional connectivity as a predictor of response to antipsychotic drug treatment. Am. J. Psychiat. 2015, 173, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Hanson, J.L.; Knodt, A.R.; Brigidi, B.D.; Hariri, A.R. Heightened connectivity between the ventral striatum and medial prefrontal cortex as a biomarker for stress-related psychopathology: understanding interactive effects of early and more recent stress. Psychol. Med. 2018, 48, 1835–1843. [Google Scholar] [CrossRef]
- Klumpp, H.; Keutmann, M.K.; Fitzgerald, D.A.; Shankman, S.A.; Phan, K.L. Resting state amygdala-prefrontal connectivity predicts symptom change after cognitive behavioral therapy in generalized social anxiety disorder. Biol. Mood Anxiety Disord. 2014, 4, 14. [Google Scholar] [CrossRef]
- Robinson, O.J.; Krimsky, M.; Lieberman, L.; Allen, P.; Vytal, K.; Grillon, C. The dorsal medial prefrontal (anterior cingulate) cortex–amygdala aversive amplification circuit in unmedicated generalised and social anxiety disorders: an observational study. Lancet Psychiatry 2014, 1, 294–302. [Google Scholar] [CrossRef]
- Feldker, K.; Heitmann, C.; Neumeister, P.; Tupak, S.; Schrammen, E.; Moeck, R.; Zwitserlood, P.; Bruchmann, M.; Straube, T. Transdiagnostic brain responses to disorder-related threat across four psychiatric disorders. Psychol. Med. 2017, 47, 730–743. [Google Scholar] [CrossRef] [PubMed]
- Robinson, O.J.; Charney, D.R.; Overstreet, C.; Vytal, K.; Grillon, C. The adaptive threat bias in anxiety: amygdala–dorsomedial prefrontal cortex coupling and aversive amplification. Neuroimage 2012, 60, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Boeke, E.A.; Moscarello, J.M.; LeDoux, J.E.; Phelps, E.A.; Hartley, C.A. Active avoidance: neural mechanisms and attenuation of Pavlovian conditioned responding. J. Neurosci. 2017, 37, 4808–4818. [Google Scholar] [CrossRef] [PubMed]
- Campese, V.D.; Gonzaga, R.; Moscarello, J.M.; LeDoux, J.E. Modulation of instrumental responding by a conditioned threat stimulus requires lateral and central amygdala. Front. Behav. Neurosci. 2015, 9, 293. [Google Scholar] [CrossRef]
- Sengupta, A.; Yau, J.O.; Jean-Richard-Dit-Bressel, P.; Liu, Y.; Millan, E.Z.; Power, J.M.; McNally, G.P. Basolateral amygdala neurons maintain aversive emotional salience. J. Neurosci. 2018, 38, 3001–3012. [Google Scholar] [CrossRef]
- Terburg, D.; Morgan, B.; Montoya, E.; Hooge, I.; Thornton, H.; Hariri, A.; Panksepp, J.; Stein, D.; Van Honk, J. Hypervigilance for fear after basolateral amygdala damage in humans. Transl. Psychiatr. 2012, 2, e115. [Google Scholar] [CrossRef]
- Carlisi, C.O.; Robinson, O.J. The role of prefrontal–subcortical circuitry in negative bias in anxiety: Translational, developmental and treatment perspectives. Brain Neurosci Adv. 2018, 2, 2398212818774223. [Google Scholar] [CrossRef] [PubMed]
- Etkin, A.; Egner, T.; Kalisch, R. Emotional processing in anterior cingulate and medial prefrontal cortex. Trends. Cogn. Sci. 2011, 15, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Kalisch, R.; Gerlicher, A.M. Making a mountain out of a molehill: On the role of the rostral dorsal anterior cingulate and dorsomedial prefrontal cortex in conscious threat appraisal, catastrophizing, and worrying. Neurosci. Biobehav. Rev. 2014, 42, 1–8. [Google Scholar] [CrossRef]
- Klavir, O.; Genud-Gabai, R.; Paz, R. Functional connectivity between amygdala and cingulate cortex for adaptive aversive learning. Neuron 2013, 80, 1290–1300. [Google Scholar] [CrossRef]
- Livneh, U.; Paz, R. Amygdala-prefrontal synchronization underlies resistance to extinction of aversive memories. Neuron 2012, 75, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Robinson, O.; Krimsky, M.; Lieberman, L.; Vytal, K.; Ernst, M.; Grillon, C. Anxiety-potentiated amygdala–medial frontal coupling and attentional control. Transl. Psychiatr. 2016, 6, e833. [Google Scholar] [CrossRef]
- Klumpp, H.; Fitzgerald, J.M.; Kinney, K.L.; Kennedy, A.E.; Shankman, S.A.; Langenecker, S.A.; Phan, K.L. Predicting cognitive behavioral therapy response in social anxiety disorder with anterior cingulate cortex and amygdala during emotion regulation. NeuroImage-Clin. 2017, 15, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, D.V.; Lecrubier, Y.; Sheehan, K.H.; Amorim, P.; Janavs, J.; Weiller, E.; Hergueta, T.; Baker, R.; Dunbar, G.C. The Mini-International Neuropsychiatric Interview (MINI): the development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J. Clin. Psychiatry 1998. [Google Scholar]
- Hariri, A.R.; Mattay, V.S.; Tessitore, A.; Kolachana, B.; Fera, F.; Goldman, D.; Egan, M.F.; Weinberger, D.R. Serotonin transporter genetic variation and the response of the human amygdala. Science 2002, 297, 400–403. [Google Scholar] [CrossRef] [PubMed]
- O’Nions, E.J.; Dolan, R.J.; Roiser, J.P. Serotonin transporter genotype modulates subgenual response to fearful faces using an incidental task. J. Cogn. Neurosci. 2011, 23, 3681–3693. [Google Scholar] [CrossRef]
- Wiggins, G.; Triantafyllou, C.; Potthast, A.; Reykowski, A.; Nittka, M.; Wald, L. 32-channel 3 tesla receive-only phased-array head coil with soccer-ball element geometry. Magn. Reson. Med. 2006, 56, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Weiskopf, N.; Hutton, C.; Josephs, O.; Deichmann, R. Optimal EPI parameters for reduction of susceptibility-induced BOLD sensitivity losses: a whole-brain analysis at 3 T and 1.5 T. Neuroimage 2006, 33, 493–504. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, J.X.; Woolrich, M.W.; Behrens, T.E.; Smith, S.M.; Johansen-Berg, H. Tools of the trade: psychophysiological interactions and functional connectivity. Soc. Cogn. Affect. Neurosci. 2012, 7, 604–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, C.M.; Miller, M.B. How reliable are the results from functional magnetic resonance imaging? Ann. N. Y. Acad. Sci. 2010, 1191, 133–155. [Google Scholar]
- Bartko, J.J. The intraclass correlation coefficient as a measure of reliability. Psychol. Rep. 1966, 19, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Shrout, P.E.; Fleiss, J.L. Intraclass Correlations: Uses in Assessing Rater Reliability. Psychol. Bull. 1979, 86, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Fleiss, J.L. Reliability of measurement. In The Design and Analysis of Clinical Experiments; John Wiley & Sons: Hoboken, NJ, USA, 1986; pp. 1–32. [Google Scholar]
- Shehzad, Z.; Kelly, A.C.; Reiss, P.T.; Gee, D.G.; Gotimer, K.; Uddin, L.Q.; Lee, S.H.; Margulies, D.S.; Roy, A.K.; Biswal, B.B.; et al. The resting brain: unconstrained yet reliable. Cereb. Cortex 2009, 19, 2209–2229. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, A.J.; McGonigle, J. Negative edges and soft thresholding in complex network analysis of resting state functional connectivity data. Neuroimage 2011, 55, 1132–1146. [Google Scholar] [CrossRef] [PubMed]
- Braun, U.; Plichta, M.M.; Esslinger, C.; Sauer, C.; Haddad, L.; Grimm, O.; Mier, D.; Mohnke, S.; Heinz, A.; Erk, S.; et al. Test–retest reliability of resting-state connectivity network characteristics using fMRI and graph theoretical measures. Neuroimage 2012, 59, 1404–1412. [Google Scholar] [CrossRef] [PubMed]
- Guijt, A.M.; Sluiter, J.K.; Frings-Dresen, M.H. Test-retest reliability of heart rate variability and respiration rate at rest and during light physical activity in normal subjects. Arch. Med. Res. 2007, 38, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, B.W.; Rajendra, J.K.; Craighead, W.E.; Kelley, M.E.; McGrath, C.L.; Choi, K.S.; Kinkead, B.; Nemeroff, C.B.; Mayberg, H.S.; et al. Functional connectivity of the subcallosal cingulate cortex and differential outcomes to treatment with cognitive-behavioral therapy or antidepressant medication for major depressive disorder. Am. J. Psychiat. 2017, 174, 533–545. [Google Scholar] [CrossRef]
- Salomons, T.V.; Dunlop, K.; Kennedy, S.H.; Flint, A.; Geraci, J.; Giacobbe, P.; Downar, J. Resting-state cortico-thalamic-striatal connectivity predicts response to dorsomedial prefrontal rTMS in major depressive disorder. Neuropsychopharmacology 2014, 39, 488–498. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | Emotion Identification (EI) | Face Matching (FM) | Gender Classification (GC) |
|---|---|---|---|
| Task duration | 4:03 | 5:55 | 6:24 |
| Task design | Event-related | Blocked | Blocked |
| Regressors of interest | Happy; fearful; neutral | Faces; shapes | Happy; fearful; neutral |
| Instruction | Explicit (match emotion) | Implicit (match face or shape to test stimuli) | Implicit (classify gender of face) |
| Regressors of no interest | 6 movement parameters | 6 movement parameters | 6 movement parameters + errors |
| Contrast | Faces > fixation | Faces > shapes | Faces > fixation |
| Task | Reliability | LAmyg ICC (95% CI) | p-value | RAmyg ICC (95% CI) | p-value | sgACC ICC (95% CI) | p-value |
|---|---|---|---|---|---|---|---|
| EI | Between-day (both runs) | 0.704 (0.340 to 0.867)* | 0.002 | 0.738 (0.427 to 0.881)* | 0.001 | 0.121 (−0.928 to 0.600) | 0.372 |
| Between-day (run 1) | 0.585 (0.074 to 0.814)* | 0.016 | 0.765 (0.485 to 0.893)* | <0.001 | 0.167 (−0.828 to 0.620) | 0.323 | |
| Between-day (run 2) | 0.573 (0.047 to 0.808) * | 0.019 | 0.555 (0.008 to 0.801)* | 0.024 | 0.017 (−1.158 to 0.552) | 0.483 | |
| Within-day (day 1) | 0.652 (0.224 to 0.844)* | 0.005 | 0.832 (0.626 to 0.925)* | <0.001 | 0.314 (−0.504 to 0.688) | 0.171 | |
| Within-day (day 2) | 0.768 (0.482 to 0.896)* | <0.001 | 0.699 (0.339 to 0.863)* | 0.002 | 0.408 (0.299 to 0.730)* | 0.094 | |
| GC | Between-day (both runs) | 0.442 (−0.206 to 0.742)* | 0.068 | 0.627 (0.206 to 0.825)* | 0.006 | Not analysed | |
| Between-day (run 1) | 0.071 (−1.01 to 0.570) | 0.425 | 0.579 (0.103 to 0.802)* | 0.013 | Not analysed | ||
| Between-day (run 2) | 0.493 (−0.095 to 0.765)* | 0.042 | 0.326 (−0.435 to 0.684) | 0.151 | Not analysed | ||
| Within-day (day 1) | 0.143 (−0.852 to 0.603) | 0.346 | 0.146 (−0.818 to 0.599) | 0.339 | Not analysed | ||
| Within-day (day 2) | 0.171 (−0.792 to 0.616) | 0.315 | −0.319 (−1.810 to 0.381) | 0.766 | Not analysed |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nord, C.L.; Gray, A.; Robinson, O.J.; Roiser, J.P. Reliability of Fronto–Amygdala Coupling during Emotional Face Processing. Brain Sci. 2019, 9, 89. https://doi.org/10.3390/brainsci9040089
Nord CL, Gray A, Robinson OJ, Roiser JP. Reliability of Fronto–Amygdala Coupling during Emotional Face Processing. Brain Sciences. 2019; 9(4):89. https://doi.org/10.3390/brainsci9040089
Chicago/Turabian StyleNord, Camilla L, Alan Gray, Oliver J Robinson, and Jonathan P Roiser. 2019. "Reliability of Fronto–Amygdala Coupling during Emotional Face Processing" Brain Sciences 9, no. 4: 89. https://doi.org/10.3390/brainsci9040089
APA StyleNord, C. L., Gray, A., Robinson, O. J., & Roiser, J. P. (2019). Reliability of Fronto–Amygdala Coupling during Emotional Face Processing. Brain Sciences, 9(4), 89. https://doi.org/10.3390/brainsci9040089