Activation of Membrane Estrogen Receptors Attenuates NOP-Mediated Tactile Antihypersensitivity in a Rodent Model of Neuropathic Pain


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Implantation of Cannulae
2.3. Spared Nerve Injury
2.4. Paw Withdrawal Assay
2.5. Drugs
2.6. Immunoblotting
2.7. Data Analysis
3. Results
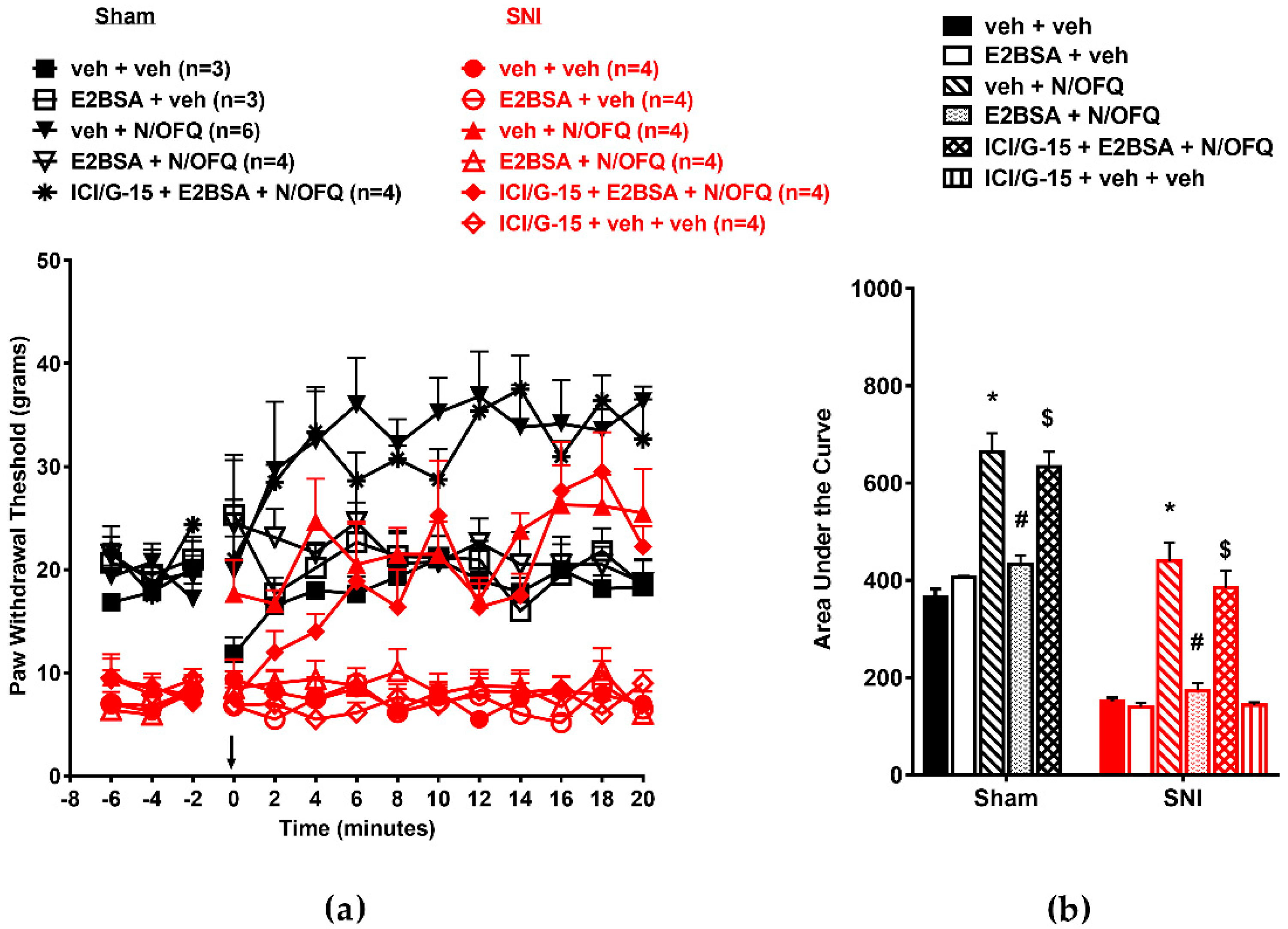
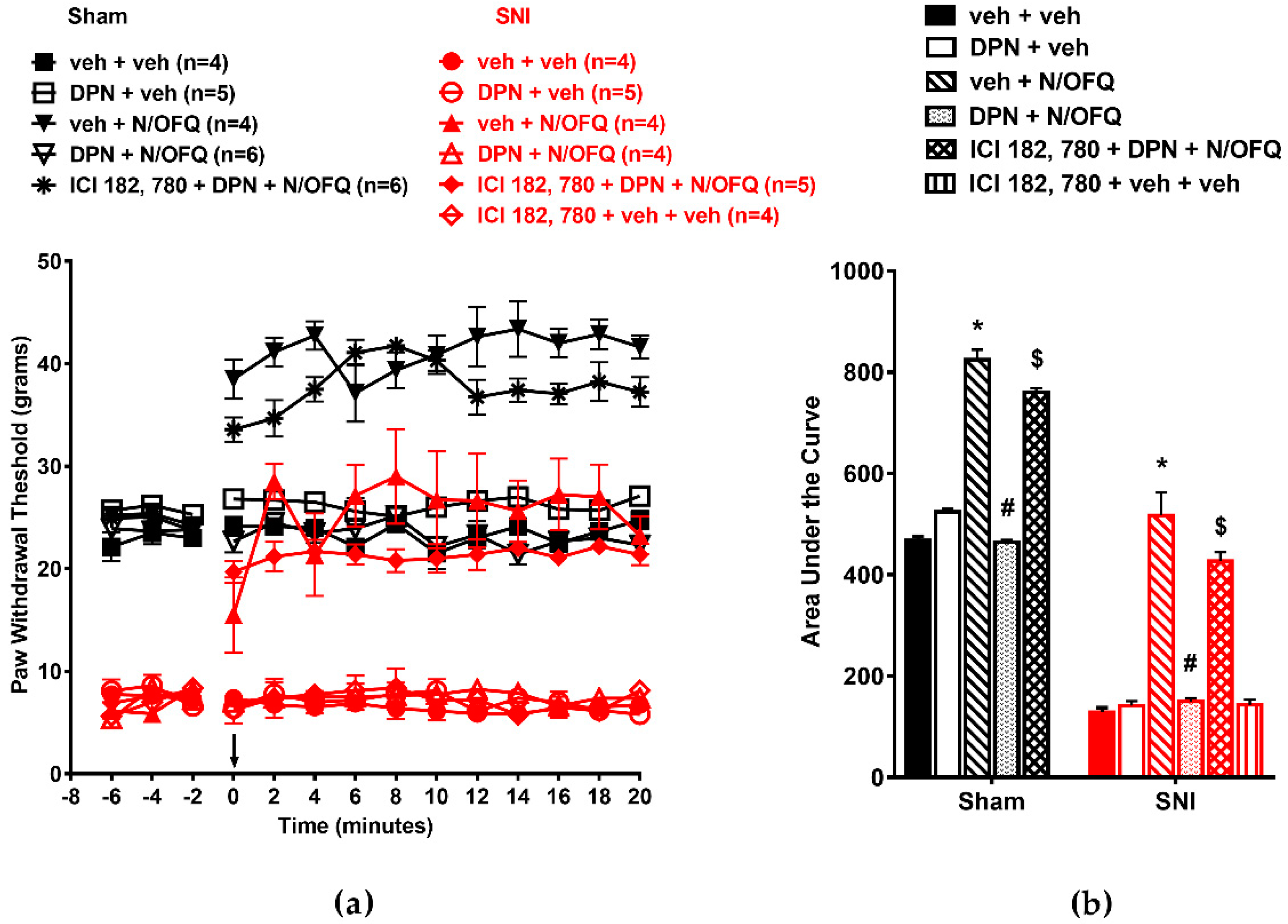
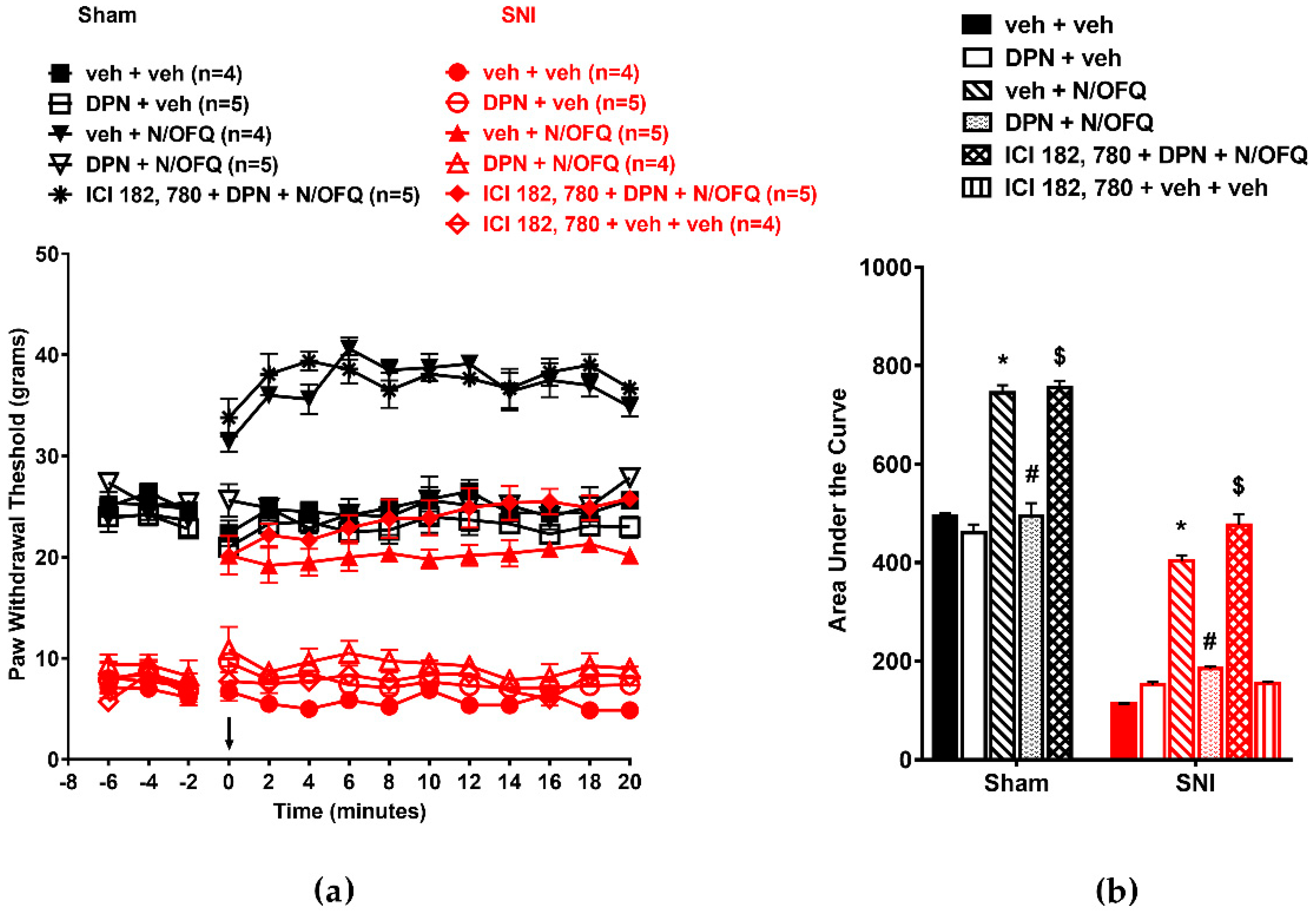
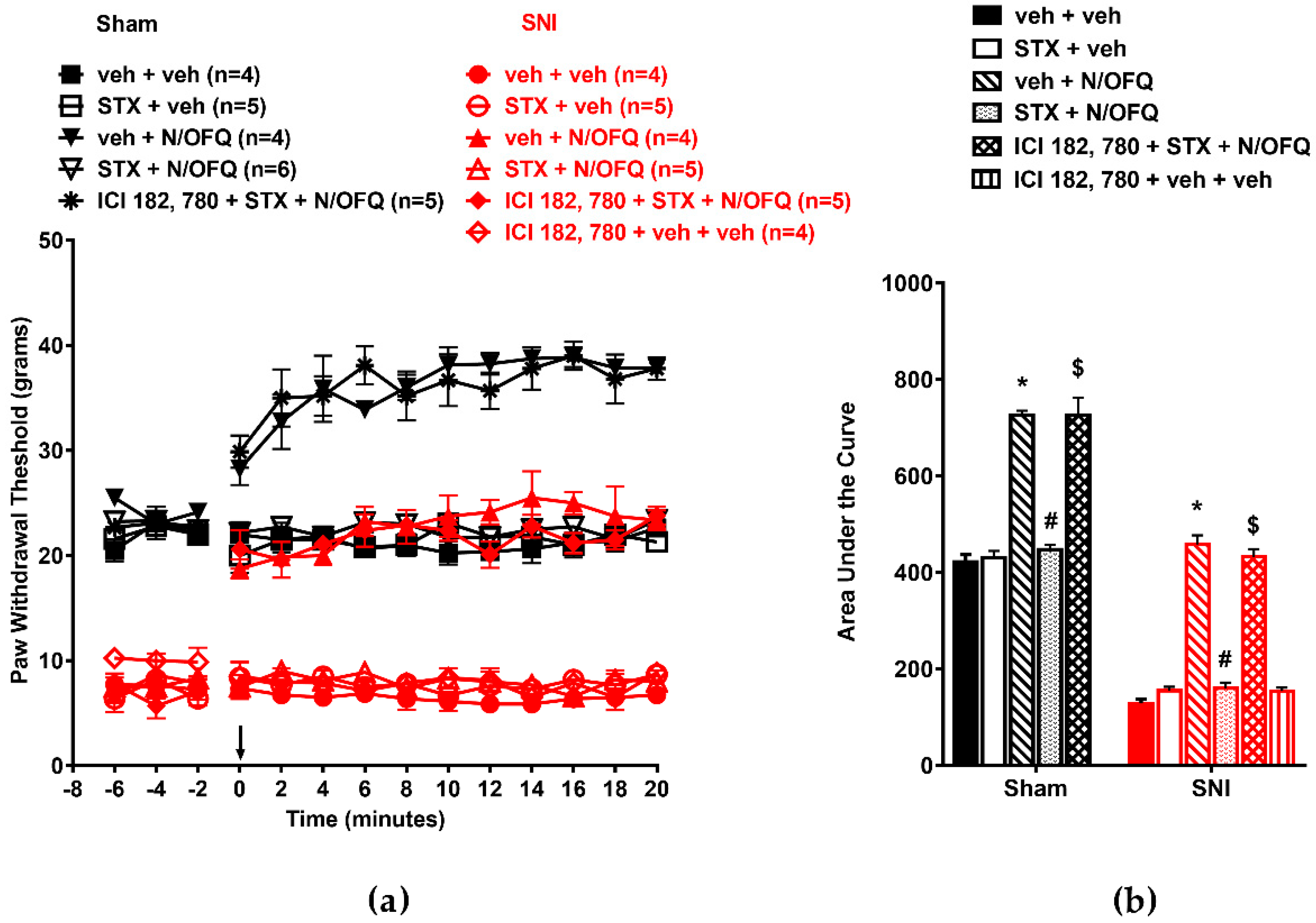
3.1. N/OFQ Reversesd Tactile Hypersensitivity following SNI, and E2-BSA Rapidly Attenuated the Effect of N/OFQ
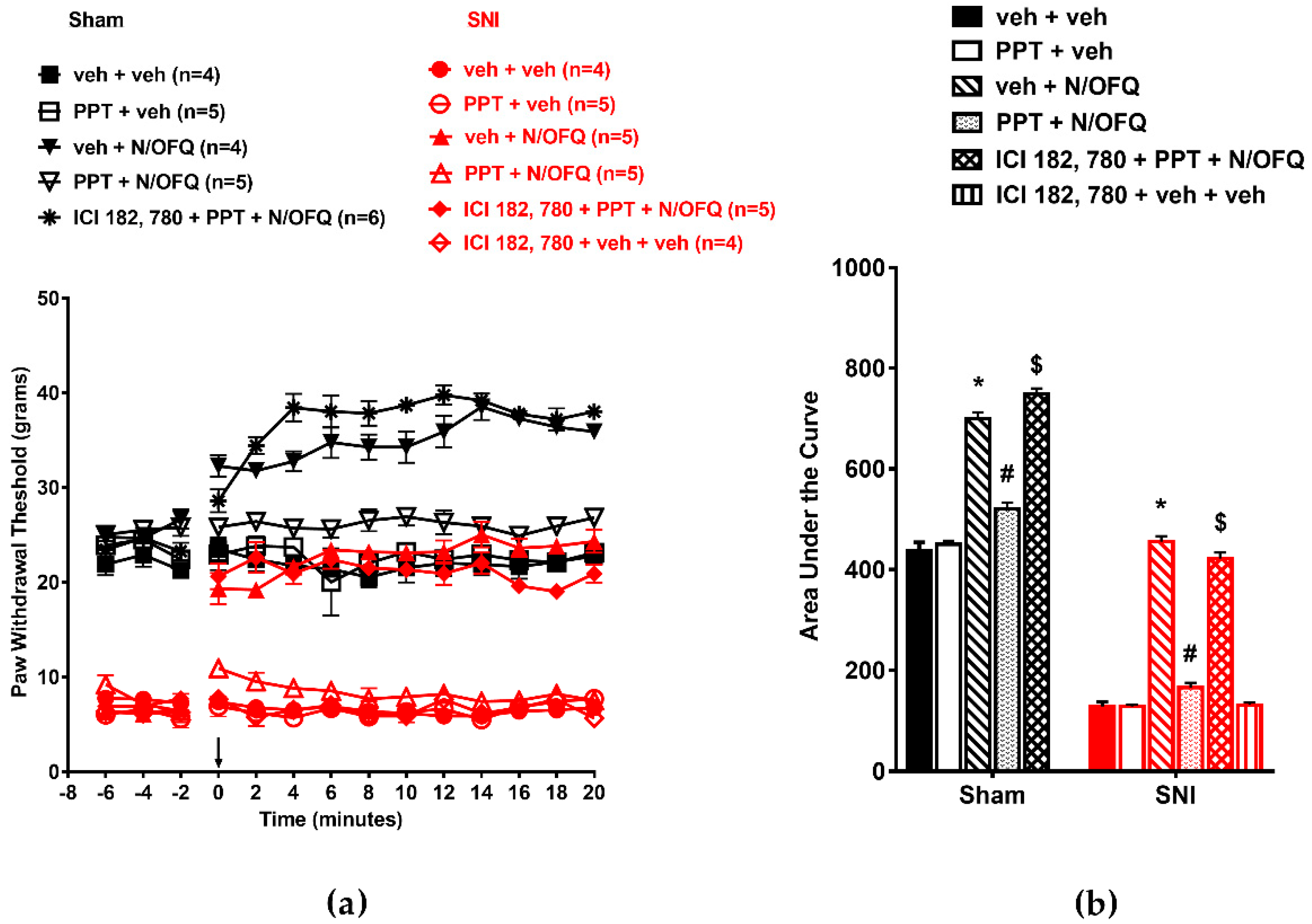
3.2. Selective Activation of ERα Rapidly Attenuated NOP-Mediated Tactile Antihypersensitivity
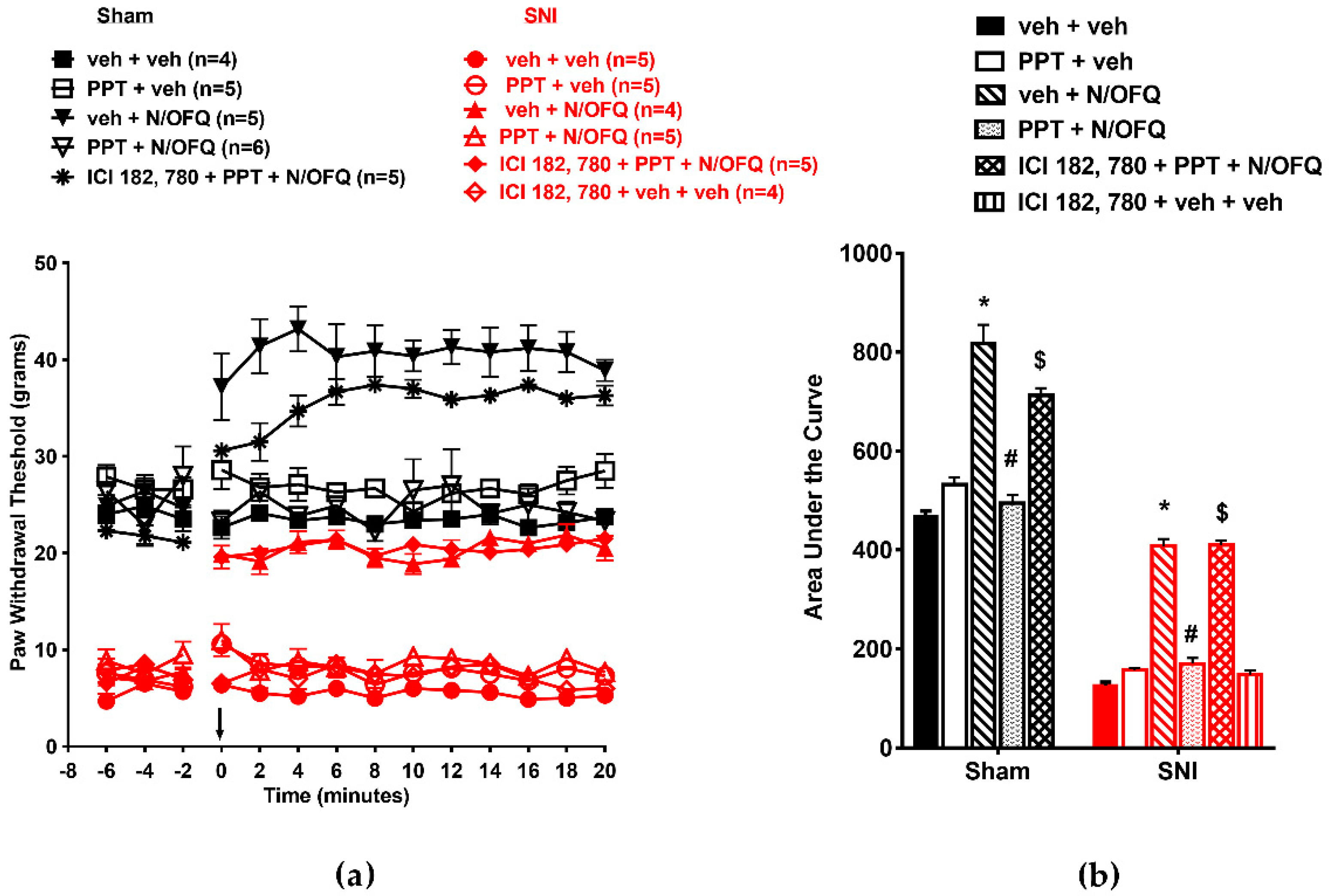
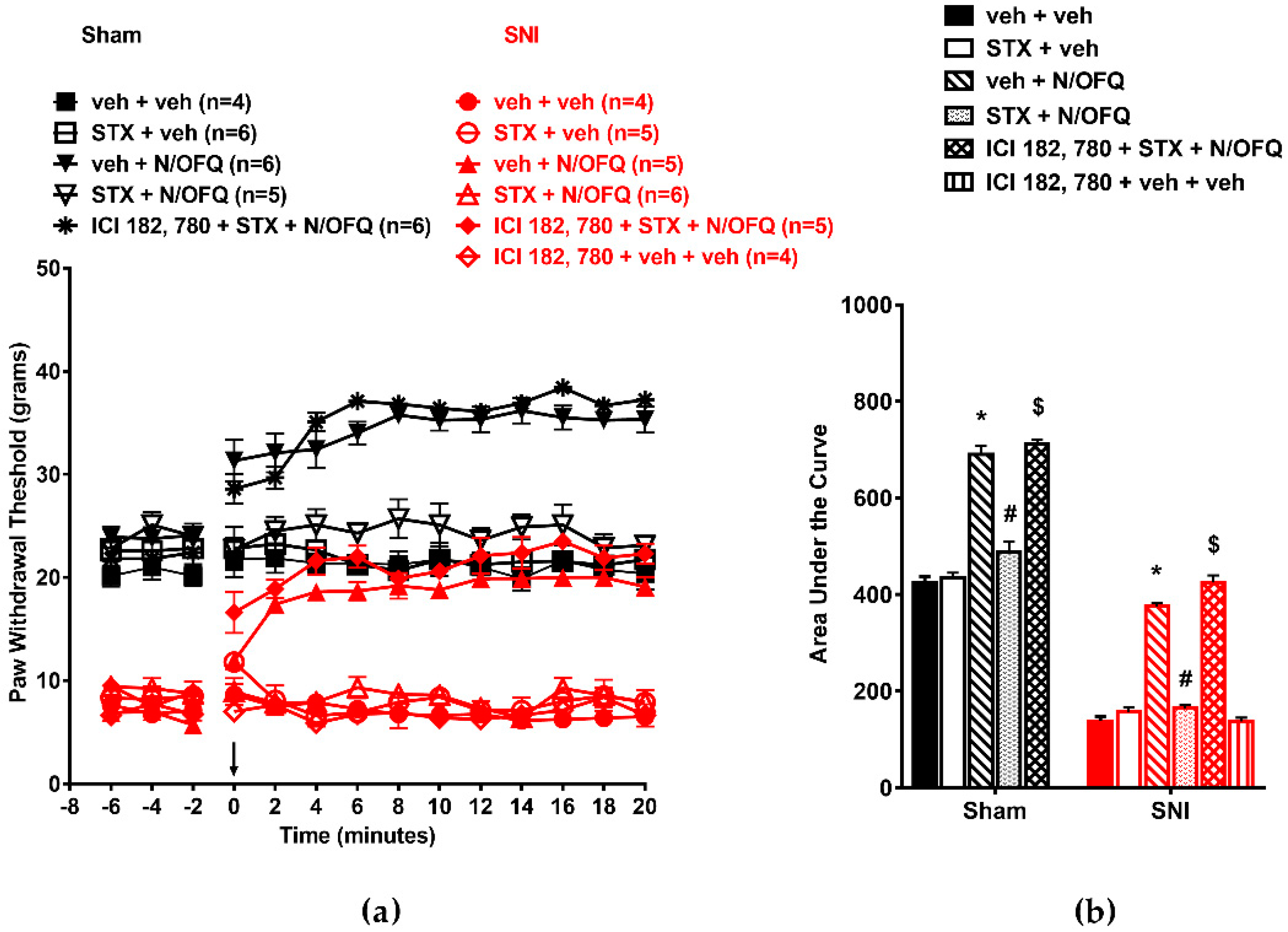
3.3. Selective Activation of ERβ Rapidly Abolished the Effect of N/OFQ
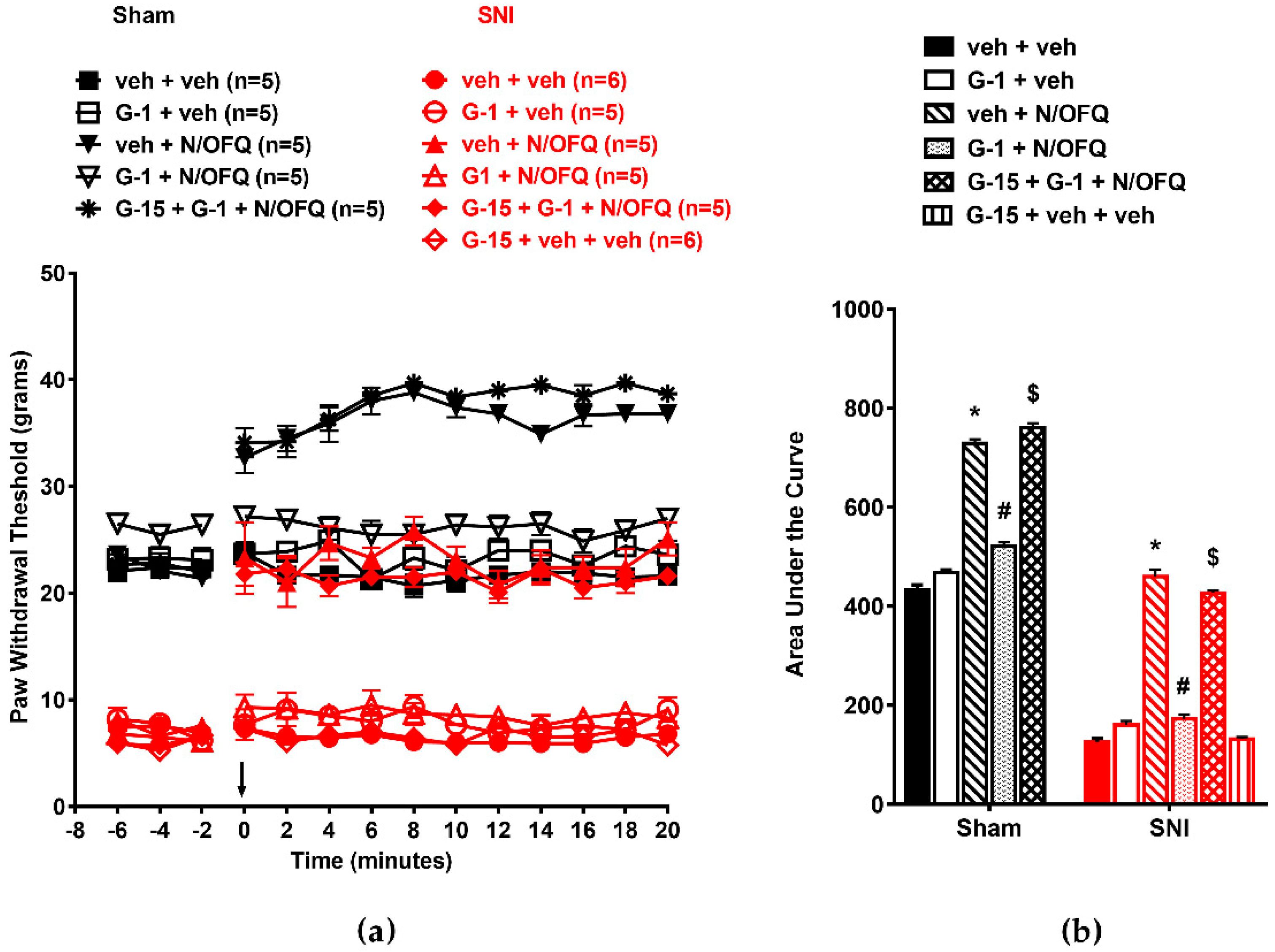
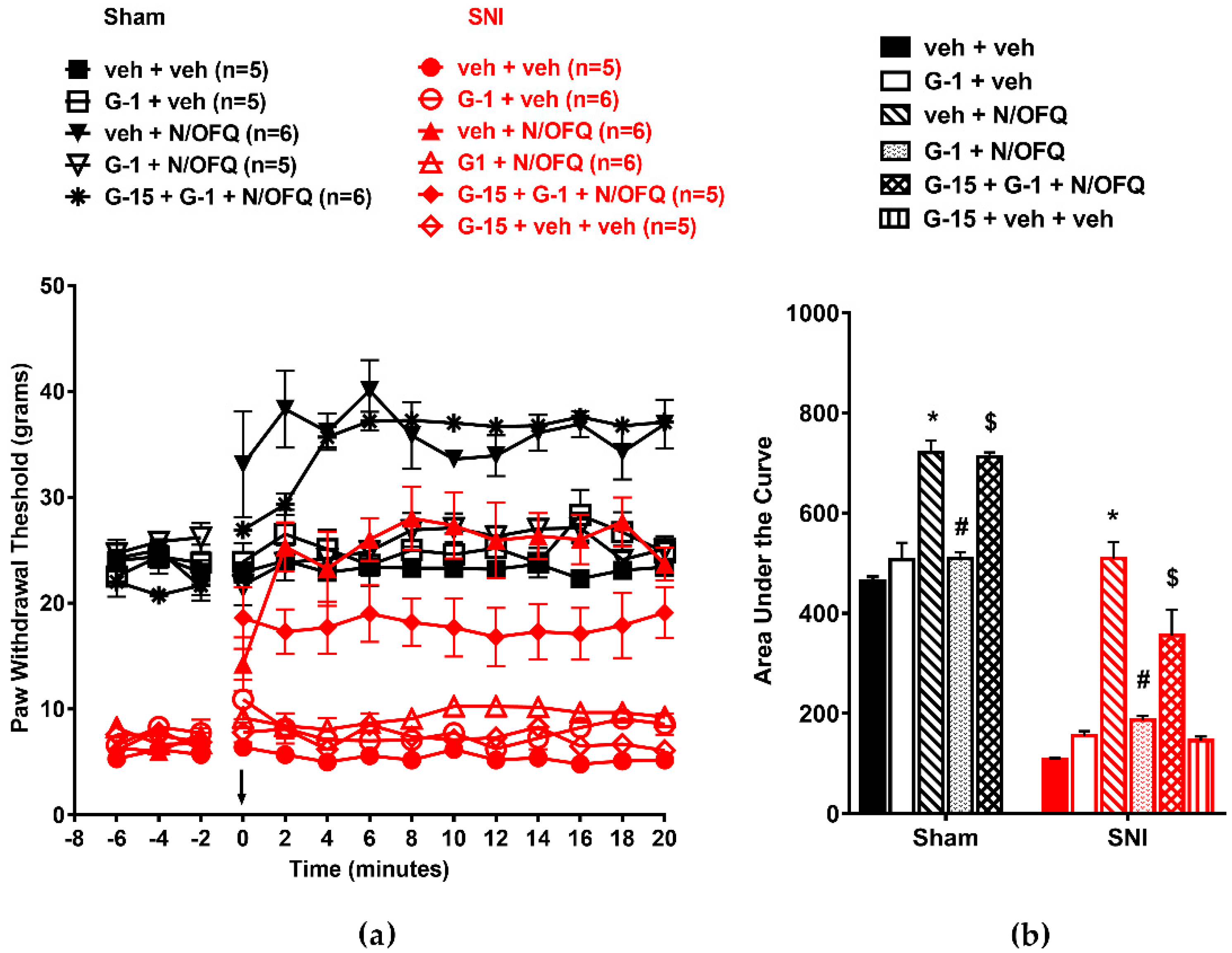
3.4. Selective Activation of GPR30 Rapidly Attenuated the Effect of N/OFQ
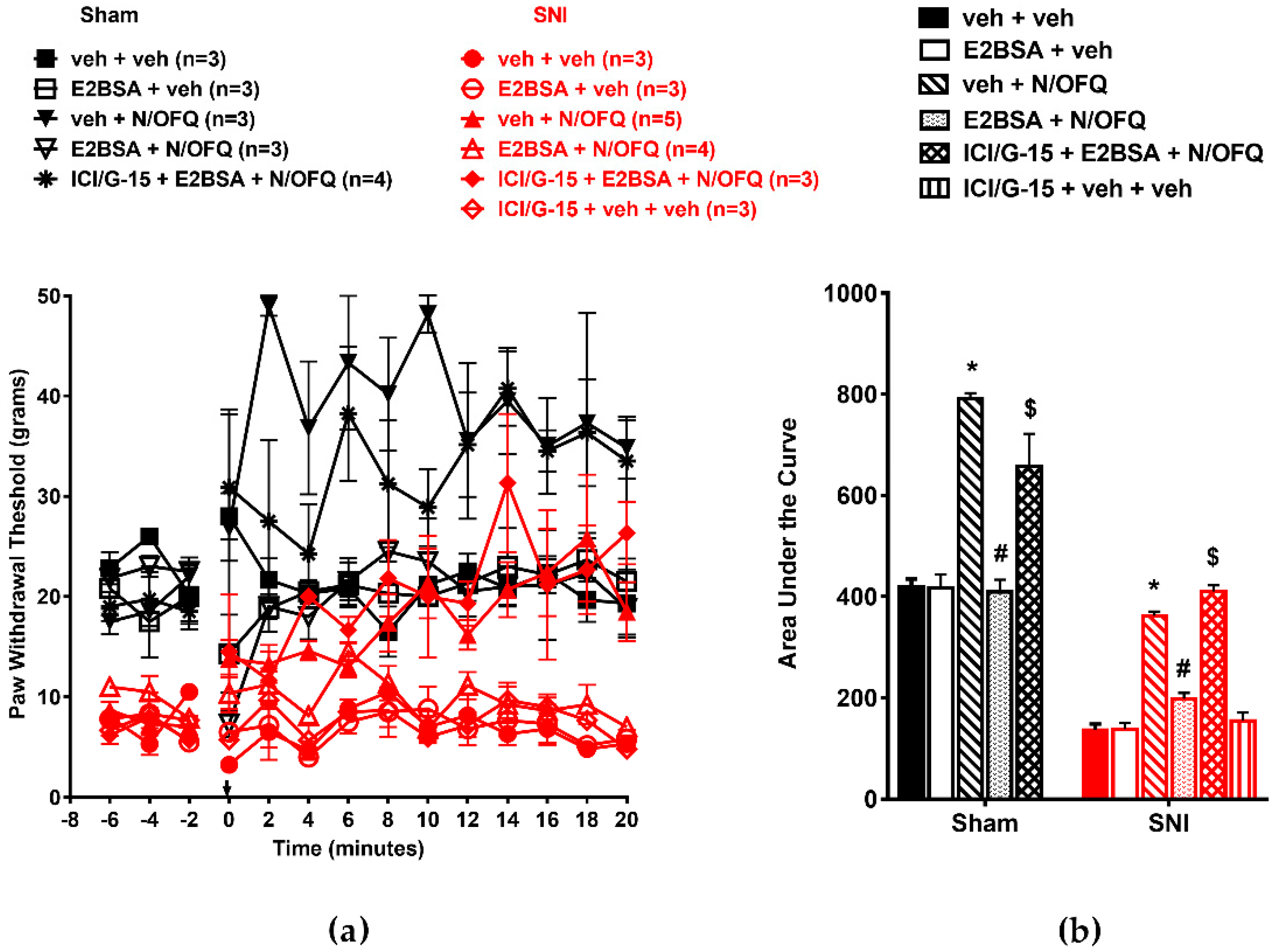
3.5. Selective Activation of Gq-mER Rapidly Abolished the Effects of N/OFQ
3.6. Activation of mERs Attenuated NOP-Mediated Tactile Antihypersensitivity via an ERK-, PKA-, PKC-, and Akt- Independent Mechanism
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Edlund, M.J.; Martin, B.C.; Fan, M.-Y.; Devries, A.; Braden, J.B.; Sullivan, M.D. Risks for Opioid Abuse and Dependence Among Recipients of Chronic Opioid Therapy: Results from the TROUP Study. Drug Alcohol Depend. 2010, 112, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Bunzow, J.R.; Saez, C.; Mortrud, M.; Bouvier, C.; Williams, J.T.; Low, M.; Grandy, D.K. Molecular cloning and tissue distribution of a putative member of the rat opioid receptor gene family that is not a mu, delta or kappa opioid receptor type. FEBS Lett. 1994, 347, 284–288. [Google Scholar] [CrossRef]
- Mollereau, C.; Parmentier, M.; Mailleux, P.; Butour, J.-L.; Moisand, C.; Chalon, P.; Caput, D.; Vassart, G.; Meunier, J.-C. ORL1, a novel member of the opioid receptor family. FEBS Lett. 1994, 341, 33–38. [Google Scholar] [CrossRef]
- Lutfy, K.; Cowan, A. Buprenorphine: A Unique Drug with Complex Pharmacology. Curr. Neuropharmacol. 2004, 2, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Ko, M.-C.; Naughton, N.N. Antinociceptive Effects of Nociceptin/Orphanin FQ Administered Intrathecally in Monkeys. J. Pain 2009, 10, 509–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, A.P.; Ko, M.C. The therapeutic potential of nociceptin/orphanin FQ receptor agonists as analgesics without abuse liability. ACS Chem. Neurosci. 2013, 4, 214–224. [Google Scholar] [CrossRef]
- Houtani, T.; Nishi, M.; Takeshima, H.; Sato, K.; Sakuma, S.; Kakimoto, S.; Ueyama, T.; Noda, T.; Sugimoto, T. Distribution of nociceptin/orphanin FQ precursor protein and receptor in brain and spinal cord: A study using in situ hybridization and X-gal histochemistry in receptor-deficient mice. J. Comp. Neurol. 2000, 424, 489–508. [Google Scholar] [CrossRef]
- Ikeda, K.; Kobayashi, K.; Kobayashi, T.; Ichikawa, T.; Kumanishi, T.; Kishida, H.; Yano, R.; Manabe, T. Functional coupling of the nociceptin/orphanin FQ receptor with the G-protein-activated K+ (GIRK) channel. Mol. Brain Res. 1997, 45, 117–126. [Google Scholar] [CrossRef]
- Schroeder, R.A.; Brandes, J.; Buse, D.C.; Calhoun, A.; Eikermann-Haerter, K.; Golden, K.; Halker, R.; Kempner, J.; Maleki, N.; Moriarty, M.; et al. Sex and Gender Differences in Migraine—Evaluating Knowledge Gaps. J. Womens Health (Larchmt) 2018, 27, 965–973. [Google Scholar] [CrossRef]
- Berkley, K.J. Sex differences in pain. Behav. Brain Sci. 1997, 20, 371–380. [Google Scholar] [CrossRef]
- Bi, R.-Y.; Ding, Y.; Gan, Y.-H. A new hypothesis of sex-differences in temporomandibular disorders: Estrogen enhances hyperalgesia of inflamed TMJ through modulating voltage-gated sodium channel 1.7 in trigeminal ganglion? Med. Hypotheses 2015, 84, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, F.; Ross, K.; Anderson, J.; Russell, I.J.; Hebert, L. The prevalence and characteristics of fibromyalgia in the general population. Arthritis Rheum. 1995, 38, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Claiborne, J.; Nag, S.; Mokha, S.S. Activation of Opioid Receptor Like-1 Receptor in the Spinal Cord Produces Sex-Specific Antinociception in the Rat: Estrogen Attenuates Antinociception in the Female, whereas Testosterone Is Required for the Expression of Antinociception in the Male. J. Neurosci. 2006, 26, 13048–13053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craft, R.M.; Ulibarri, C.; Leitl, M.D.; Sumner, J.E. Dose- and time-dependent estradiol modulation of morphine antinociception in adult female rats. Eur. J. Pain 2008, 12, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Lawson, K.P.; Nag, S.; Thompson, A.D.; Mokha, S.S. Sex-specificity and estrogen-dependence of kappa opioid receptor-mediated antinociception and antihyperalgesia. Pain 2010, 151, 806–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.-J.; Chakrabarti, S.; Schnell, S.; Wessendorf, M.; Gintzler, A.R. Spinal Synthesis of Estrogen and Concomitant Signaling by Membrane Estrogen Receptors Regulate Spinal κ- and μ-Opioid Receptor Heterodimerization and Female-Specific Spinal Morphine Antinociception. J. Neurosci. 2011, 31, 11836–11845. [Google Scholar] [CrossRef] [PubMed]
- Peckham, E.M.; Traynor, J.R. Comparison of the antinociceptive response to morphine and morphine-like compounds in male and female Sprague-Dawley rats. J. Pharmacol. Exp. Ther. 2006, 316, 1195–1201. [Google Scholar] [CrossRef] [PubMed]
- Small, K.M.; Nag, S.; Mokha, S.S. Activation of membrane estrogen receptors attenuates opioid receptor-like1 receptor-mediated antinociception via an ERK-dependent non-genomic mechanism. Neuroscience 2013, 255, 177–190. [Google Scholar] [CrossRef] [Green Version]
- Kelly, M.J.; Qiu, J.; Ronnekleiv, O.K. Estrogen modulation of G-protein-coupled receptor activation of potassium channels in the central nervous system. Ann. N. Y. Acad. Sci. 2003, 1007, 6–16. [Google Scholar] [CrossRef]
- Nag, S.; Mokha, S.S. Activation of a Gq-coupled membrane estrogen receptor rapidly attenuates α2-adrenoceptor-induced antinociception via an ERK I/II-dependent, non-genomic mechanism in the female rat. Neuroscience 2014, 267, 122–134. [Google Scholar] [CrossRef]
- Decosterd, I.; Woolf, C.J. Spared nerve injury: An animal model of persistent peripheral neuropathic pain. Pain 2000, 87, 149–158. [Google Scholar] [CrossRef]
- Saleh, M.C.; Connell, B.J.; Saleh, T.M. Medullary and intrathecal injections of 17beta-estradiol in male rats. Brain Res. 2000, 867, 200–209. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, X.; Zhang, X.M.; Zhao, Z.Q.; Zhang, Y.Q. Estrogen facilitates spinal cord synaptic transmission via membrane-bound estrogen receptors: implications for pain hypersensitivity. J. Biol. Chem. 2012, 287, 33268–33281. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.-Y.; Chen, G.-D.; Tung, K.-C.; Chien, Y.-W.; Lai, C.-Y.; Hsieh, M.-C.; Chiu, C.-H.; Lai, C.-H.; Lee, S.-D.; Lin, T.-B. Estrogen-dependent facilitation on spinal reflex potentiation involves the Cdk5/ERK1/2/NR2B cascade in anesthetized rats. Am. J. Physiol. Metab. 2009, 297, E416–E426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bologa, C.G.; Revankar, C.M.; Young, S.M.; Edwards, B.S.; Arterburn, J.B.; Kiselyov, A.S.; A Parker, M.; E Tkachenko, S.; Savchuck, N.P.; A Sklar, L.; et al. Virtual and biomolecular screening converge on a selective agonist for GPR30. Nat. Methods 2006, 2, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Bosch, M.A.; Tobias, S.C.; Grandy, D.K.; Scanlan, T.S.; Rønnekleiv, O.K.; Kelly, M.J. Rapid Signaling of Estrogen in Hypothalamic Neurons Involves a Novel G-Protein-Coupled Estrogen Receptor that Activates Protein Kinase C. J. Neurosci. 2003, 23, 9529–9540. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.; Bosch, M.A.; Tobias, S.C.; Krust, A.; Graham, S.M.; Murphy, S.J.; Korach, K.S.; Chambon, P.; Scanlan, T.S.; Rønnekleiv, O.K.; et al. A G-Protein-Coupled Estrogen Receptor Is Involved in Hypothalamic Control of Energy Homeostasis. J. Neurosci. 2006, 26, 5649–5655. [Google Scholar] [CrossRef] [Green Version]
- Stevis, P.E.; Deecher, D.C.; Suhadolnik, L.; Mallis, L.M.; Frail, D.E. Differential Effects of Estradiol and Estradiol-BSA Conjugates. Endocrinology 1999, 140, 5455–5458. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Boil. Chem. 1951, 193, 265–275. [Google Scholar]
- Honda, K.; Sawada, H.; Kihara, T.; Urushitani, M.; Nakamizo, T.; Akaike, A.; Shimohama, S. Phosphatidylinositol 3-kinase mediates neuroprotection by estrogen in cultured cortical neurons. J. Neurosci. Res. 2000, 60, 321–327. [Google Scholar] [CrossRef]
- Kelly, M. Rapid effects of estrogen to modulate G protein-coupled receptors via activation of protein kinase A and protein kinase C pathways. Steroids 1999, 64, 64–75. [Google Scholar] [CrossRef]
- Kelly, M.J.; Rønnekleiv, O.K. Control of CNS neuronal excitability by estrogens via membrane-initiated signaling. Mol. Cell. Endocrinol. 2009, 308, 17–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Setalo, G.; Guan, X.; Warren, M.; Toran-Allerand, C.D. Estrogen-Induced Activation of Mitogen-Activated Protein Kinase in Cerebral Cortical Explants: Convergence of Estrogen and Neurotrophin Signaling Pathways. J. Neurosci. 1999, 19, 1179–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toran-Allerand, C.D. Estrogen and the brain: beyond ER-alpha, ER-beta, and 17beta-estradiol. Ann. N. Y. Acad. Sci. 2005, 1052, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Wade, C.B.; Robinson, S.; Shapiro, R.A.; Dorsa, D.M. Estrogen Receptor (ER)α and ERβ Exhibit Unique Pharmacologic Properties When Coupled to Activation of the Mitogen-Activated Protein Kinase Pathway 1. Endocrinology 2001, 142, 2336–2342. [Google Scholar] [CrossRef] [PubMed]
- Neary, J.T. Protein kinase signaling cascades in CNS trauma. IUBMB Life 2005, 57, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M.; Mizokoshi, A.; Shigemoto-Mogami, Y.; Koizumi, S.; Inoue, K. Activation of p38 mitogen-activated protein kinase in spinal hyperactive microglia contributes to pain hypersensitivity following peripheral nerve injury. Glia 2004, 45, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhang, F.-X.; Huang, F.; Lu, Y.-J.; Li, G.-D.; Bao, L.; Xiao, H.-S.; Zhang, X. Peripheral nerve injury induces trans-synaptic modification of channels, receptors and signal pathways in rat dorsal spinal cord. Eur. J. Neurosci. 2004, 19, 871–883. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.-Q.; Liu, S.; He, D.-D.; Liu, Y.-P.; Song, X.-J. Activation of the cAMP-PKA signaling pathway in rat dorsal root ganglion and spinal cord contributes toward induction and maintenance of bone cancer pain. Behav. Pharmacol. 2014, 25, 267–276. [Google Scholar] [CrossRef]
- Calò, G.; Rizzi, A.; Marzola, G.; Guerrini, R.; Salvadori, S.; Beani, L.; Regoli, D.; Bianchi, C. Pharmacological characterization of the nociceptin receptor mediating hyperalgesia in the mouse tail withdrawal assay. Br. J. Pharmacol. 1998, 125, 373–378. [Google Scholar] [CrossRef] [Green Version]
- Reinscheid, R.K.; Nothacker, H.-P.; Bourson, A.; Ardati, A.; Henningsen, R.A.; Bunzow, J.R.; Grandy, D.K.; Langen, H.; Monsma, F.J.; Civelli, O. Orphanin FQ: A Neuropeptide That Activates an Opioidlike G Protein-Coupled Receptor. Science 1995, 270, 792–794. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.J.; Hao, J.X.; Wiesenfeld-Hallin, Z. Nociceptin or antinociceptin: potent spinal antinociceptive effect of orphanin FQ/nociceptin in the rat. NeuroReport 1996, 7, 2092–2094. [Google Scholar] [PubMed]
- Chen, Y.; Sommer, C.; Claudia, S. Activation of the nociceptin opioid system in rat sensory neurons produces antinociceptive effects in inflammatory pain: Involvement of inflammatory mediators. J. Neurosci. Res. 2007, 85, 1478–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courteix, C.; Coudoré-Civiale, M.-A.; Privat, A.-M.; Pelissier, T.; Eschalier, A.; Fialip, J. Evidence for an exclusive antinociceptive effect of nociceptin/orphanin FQ, an endogenous ligand for the ORL1 receptor, in two animal models of neuropathic pain. Pain 2004, 110, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Conde, K.; Meza, C.; Kelly, M.J.; Sinchak, K.; Wagner, E.J.; Conde, K. Estradiol rapidly attenuates ORL-1 receptor-mediated inhibition of proopiomelanocortin neurons via Gq-coupled, membrane-initiated signaling. Neuroendocrinology 2016, 103, 787–805. [Google Scholar] [CrossRef] [PubMed]
- Dun, S.L.; Brailoiu, G.C.; Gao, X.; Brailoiu, E.; Arterburn, J.B.; Prossnitz, E.R.; Oprea, T.I.; Dun, N.J. Expression of estrogen receptor GPR30 in the rat spinal cord and in autonomic and sensory ganglia. J. Neurosci. Res. 2009, 87, 1610–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappas, T.C.; Gametchu, B.; Watson, C.S. Membrane estrogen receptors identified by multiple antibody labeling and impeded-ligand binding. FASEB J. 1995, 9, 404–410. [Google Scholar] [CrossRef]
- Razandi, M.; Pedram, A.; Greene, G.L.; Levin, E.R. Cell Membrane and Nuclear Estrogen Receptors (ERs) Originate from a Single Transcript: Studies of ERα and ERβ Expressed in Chinese Hamster Ovary Cells. Mol. Endocrinol. 1999, 13, 307–319. [Google Scholar] [CrossRef]
- Thomas, P.; Pang, Y.; Filardo, E.J.; Dong, J. Identity of an Estrogen Membrane Receptor Coupled to a G Protein in Human Breast Cancer Cells. Endocrinology 2005, 146, 624–632. [Google Scholar] [CrossRef] [Green Version]
- Levin, E.R. Membrane oestrogen receptor α signalling to cell functions. J. Physiol. 2009, 587, 5019–5023. [Google Scholar] [CrossRef]
- Raz, L.; Khan, M.M.; Mahesh, V.B.; Vadlamudi, R.K.; Brann, D.W. Rapid Estrogen Signaling in the Brain. Neurosignals 2008, 16, 140–153. [Google Scholar] [CrossRef] [PubMed]
- Roepke, T.A.; Qiu, J.; Bosch, M.A.; Rønnekleiv, O.K.; Kelly, M.J. Cross-talk between membrane-initiated and nuclear-initiated oestrogen signaling in the hypothalamus. J. Neuroendocr. 2009, 21, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.J.; Qiu, J.; Wagner, E.J.; Rønnekleiv, O.K. Rapid effects of estrogen on G protein-coupled receptor activation of potassium channels in the central nervous system (CNS). J. Steroid Biochem. Mol. Boil. 2002, 83, 187–193. [Google Scholar] [CrossRef]
- Prossnitz, E.R.; Arterburn, J.B.; Smith, H.O.; Oprea, T.I.; Sklar, L.A.; Hathaway, H.J. Estrogen Signaling through the Transmembrane G Protein–Coupled Receptor GPR30. Annu. Rev. Physiol. 2008, 70, 165–190. [Google Scholar] [CrossRef] [PubMed]
- Flores, C.; Shughrue, P.; Petersen, S.; Mokha, S. Sex-related differences in the distribution of opioid receptor-like 1 receptor mRNA and colocalization with estrogen receptor mRNA in neurons of the spinal trigeminal nucleus caudalis in the rat. Neuroscience 2003, 118, 769–778. [Google Scholar] [CrossRef]
- Filardo, E.J.; Quinn, J.A.; Bland, K.I.; Frackelton, A.R. Estrogen-Induced Activation of Erk-1 and Erk-2 Requires the G Protein-Coupled Receptor Homolog, GPR30, and Occurs via Trans-Activation of the Epidermal Growth Factor Receptor through Release of HB-EGF. Mol. Endocrinol. 2000, 14, 1649–1660. [Google Scholar] [CrossRef] [PubMed]
- Filardo, E.J.; Quinn, J.A.; Frackelton, A.R.; Bland, K.I. Estrogen Action Via the G Protein-Coupled Receptor, GPR30: Stimulation of Adenylyl Cyclase and cAMP-Mediated Attenuation of the Epidermal Growth Factor Receptor-to-MAPK Signaling Axis. Mol. Endocrinol. 2002, 16, 70–84. [Google Scholar] [CrossRef] [PubMed]
- Filardo, E.J.; Thomas, P. Minireview: G Protein-Coupled Estrogen Receptor-1, GPER-1: Its Mechanism of Action and Role in Female Reproductive Cancer, Renal and Vascular Physiology. Endocrinology 2012, 153, 2953–2962. [Google Scholar] [CrossRef] [Green Version]
- Revankar, C.M. A Transmembrane Intracellular Estrogen Receptor Mediates Rapid Cell Signaling. Science 2005, 307, 1625–1630. [Google Scholar] [CrossRef] [Green Version]
- Revankar, C.M.; Mitchell, H.D.; Field, A.S.; Burai, R.; Corona, C.; Ramesh, C.; Sklar, L.A.; Arterburn, J.B.; Prossnitz, E.R. Synthetic Estrogen Derivatives Demonstrate the Functionality of Intracellular GPR30. ACS Chem. Boil. 2007, 2, 536–544. [Google Scholar] [CrossRef]
- Dickson, R.B.; Clark, C.R. Estrogen Receptors in the Male. Arch. Androl. 1981, 7, 205–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]










© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wright, D.M.; Small, K.M.; Nag, S.; Mokha, S.S. Activation of Membrane Estrogen Receptors Attenuates NOP-Mediated Tactile Antihypersensitivity in a Rodent Model of Neuropathic Pain. Brain Sci. 2019, 9, 147. https://doi.org/10.3390/brainsci9060147
Wright DM, Small KM, Nag S, Mokha SS. Activation of Membrane Estrogen Receptors Attenuates NOP-Mediated Tactile Antihypersensitivity in a Rodent Model of Neuropathic Pain. Brain Sciences. 2019; 9(6):147. https://doi.org/10.3390/brainsci9060147
Chicago/Turabian StyleWright, Danyeal M., Keri M. Small, Subodh Nag, and Sukhbir S. Mokha. 2019. "Activation of Membrane Estrogen Receptors Attenuates NOP-Mediated Tactile Antihypersensitivity in a Rodent Model of Neuropathic Pain" Brain Sciences 9, no. 6: 147. https://doi.org/10.3390/brainsci9060147
APA StyleWright, D. M., Small, K. M., Nag, S., & Mokha, S. S. (2019). Activation of Membrane Estrogen Receptors Attenuates NOP-Mediated Tactile Antihypersensitivity in a Rodent Model of Neuropathic Pain. Brain Sciences, 9(6), 147. https://doi.org/10.3390/brainsci9060147