Effects of Sound-Pressure Change on the 40 Hz Auditory Steady-State Response and Change-Related Cerebral Response


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Sound Stimuli
2.3. MEG Recording
2.4. Dipole Source Modeling
2.5. Data Analysis
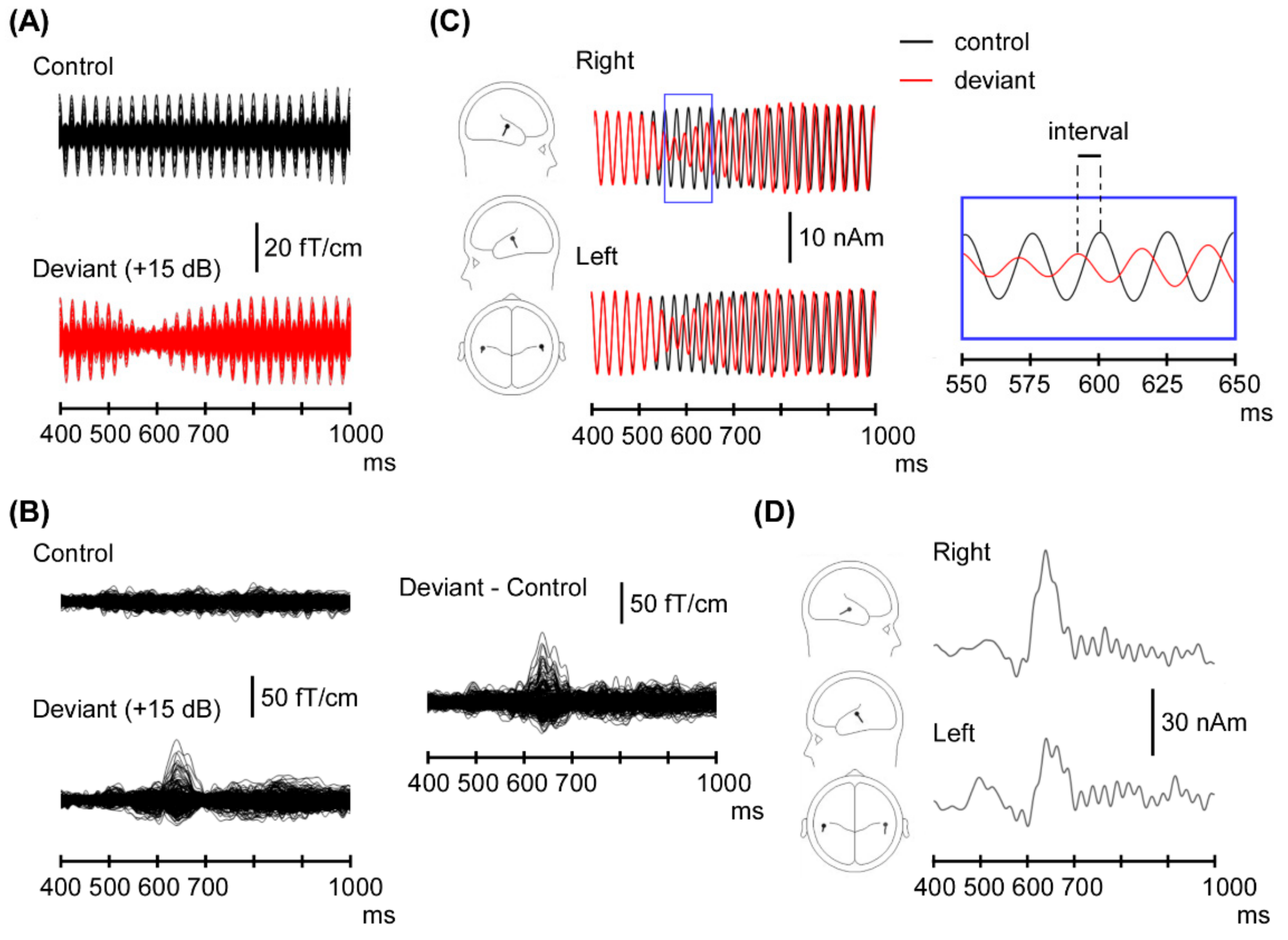
3. Results
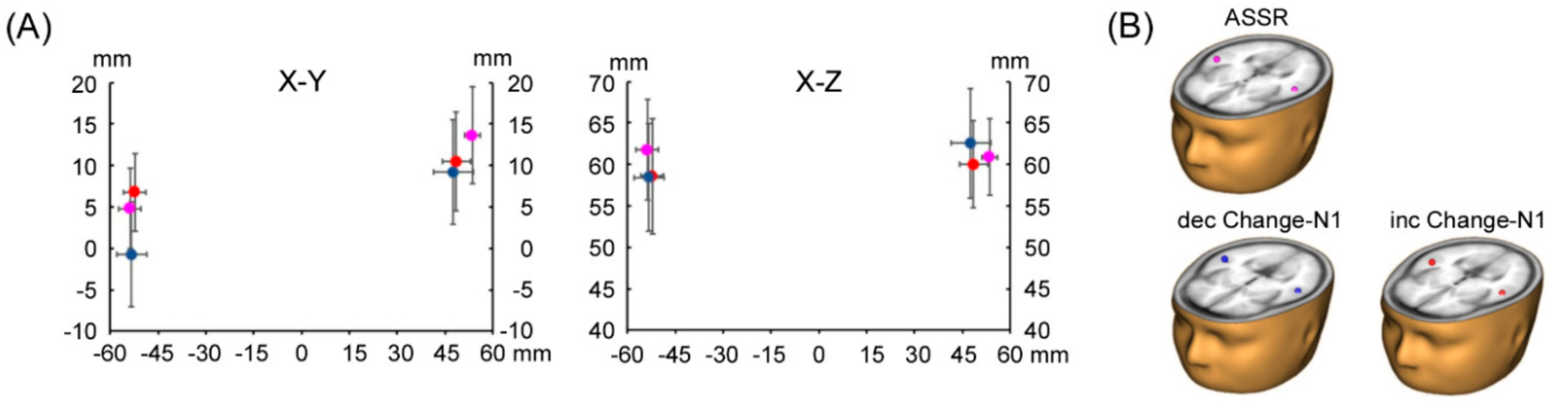
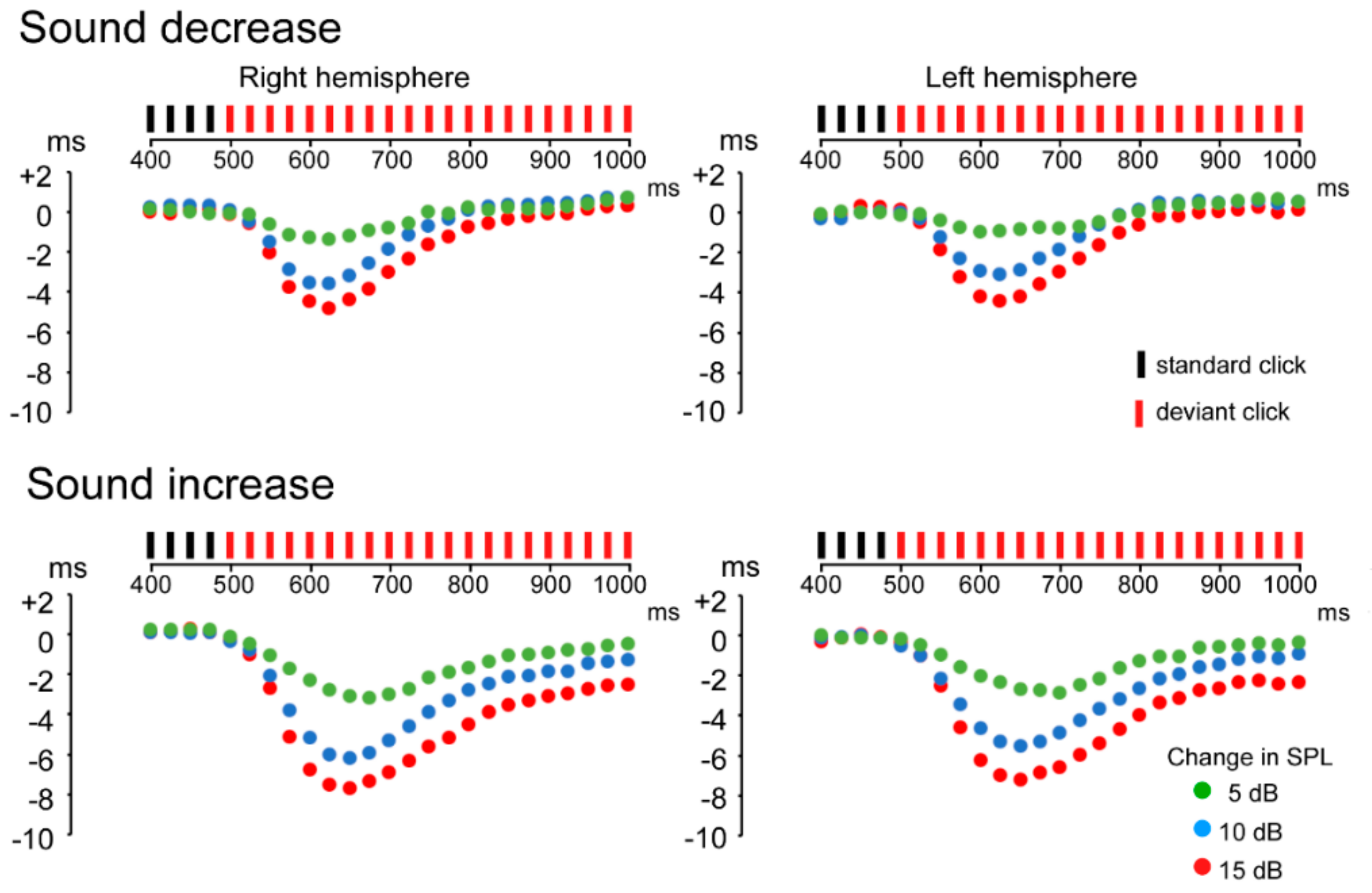
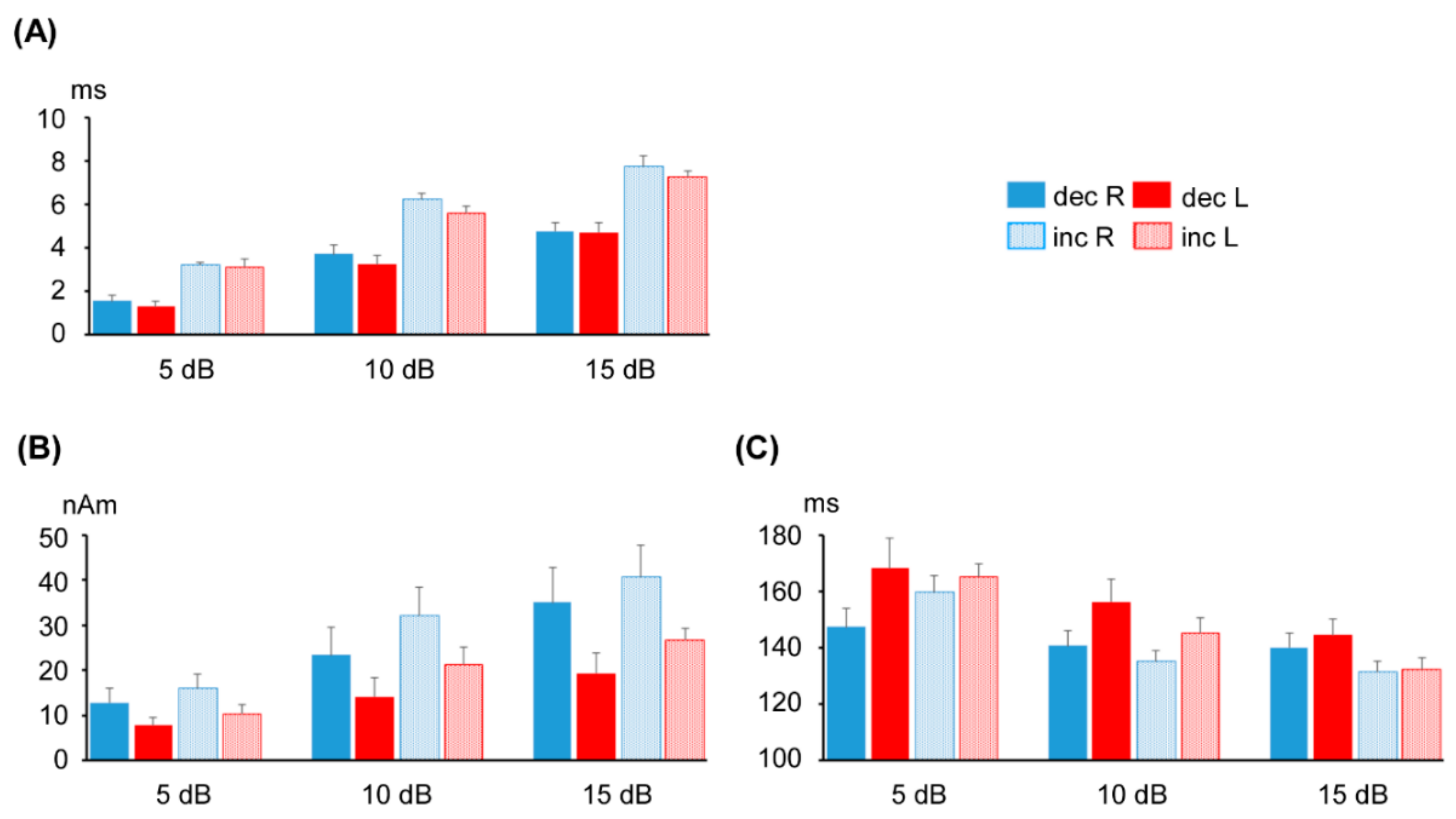
3.1. ASSR Phase Deviation
3.2. Change-N1m Values
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Inui, K.; Urakawa, T.; Yamashiro, K.; Otsuru, N.; Nishihara, M.; Takeshima, Y.; Keceli, S.; Kakigi, R. Non-linear laws of echoic memory and auditory change detection in humans. BMC Neurosci. 2010, 11, 80. [Google Scholar] [CrossRef]
- Yamashiro, K.; Inui, K.; Otsuru, N.; Kakigi, R. Change-related responses in the human auditory cortex: An MEG study. Psychophysiology 2011, 48, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Weise, A.; Schroger, E.; Feher, B.; Folyi, T.; Horvath, J. Auditory event-related potentials reflect dedicated change detection activity for higher-order acoustic transitions. Biol. Psychol. 2012, 91, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Weise, A.; Bendixen, A.; Muller, D.; Schroger, E. Which kind of transition is important for sound representation? An event-related potential study. Brain Res. 2012, 1464, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.C.; Mills, J.H.; Dubno, J.R. Electrophysiologic correlates of intensity discrimination in cortical evoked potentials of younger and older adults. Hear. Res. 2007, 228, 58–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soeta, Y.; Nakagawa, S. Temporal integration affects intensity change detection in human auditory cortex. Neuroreport 2010, 21, 1157–1161. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, M.; Inui, K.; Motomura, E.; Otsuru, N.; Ushida, T.; Kakigi, R. Auditory N1 as a change-related automatic response. Neurosci. Res. 2011, 71, 145–148. [Google Scholar] [CrossRef]
- Otsuru, N.; Tsuruhara, A.; Motomura, E.; Tanii, H.; Nishihara, M.; Inui, K.; Kakigi, R. Effects of acute nicotine on auditory change-related cortical responses. Psychopharmacology 2012, 224, 327–335. [Google Scholar] [CrossRef]
- Soeta, Y.; Nakagawa, S. Auditory evoked responses in human auditory cortex to the variation of sound intensity in an ongoing tone. Hear. Res. 2012, 287, 67–75. [Google Scholar] [CrossRef]
- Kodaira, M.; Tsuruhara, A.; Motomura, E.; Tanii, H.; Inui, K.; Kakigi, R. Effects of acute nicotine on prepulse inhibition of auditory change-related cortical responses. Behav. Brain Res. 2013, 256, 27–35. [Google Scholar] [CrossRef]
- Akiyama, L.F.; Yamashiro, K.; Inui, K.; Kakigi, R. Automatic cortical responses to sound movement: A magnetoencephalography study. Neurosci. Lett. 2011, 488, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Ohoyama, K.; Motomura, E.; Inui, K.; Nishihara, M.; Otsuru, N.; Oi, M.; Kakigi, R.; Okada, M. Memory-based pre-attentive auditory N1 elicited by sound movement. Neurosci. Res. 2012, 73, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K.; Otsuru, N.; Inui, K.; Kakigi, R. Change-related auditory P50: A MEG study. NeuroImage 2014, 86, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.J.; Longe, O.; Vaz Pato, M. Auditory evoked potentials to abrupt pitch and timbre change of complex tones: Electrophysiological evidence of ‘streaming’? Electroencephalogr. Clin. Neurophysiol. 1998, 108, 131–142. [Google Scholar] [CrossRef]
- Inui, K.; Urakawa, T.; Yamashiro, K.; Otsuru, N.; Takeshima, Y.; Nishihara, M.; Motomura, E.; Kida, T.; Kakigi, R. Echoic memory of a single pure tone indexed by change-related brain activity. BMC Neurosci. 2010, 11, 135. [Google Scholar] [CrossRef] [PubMed]
- Tanahashi, M.; Motomura, E.; Inui, K.; Ohoyama, K.; Tanii, H.; Konishi, Y.; Shiroyama, T.; Nishihara, M.; Kakigi, R.; Okada, M. Auditory change-related cerebral responses and personality traits. Neurosci. Res. 2016, 103, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Ross, B.; Herdman, A.T.; Pantev, C. Stimulus induced desynchronization of human auditory 40-Hz steady-state responses. J. Neurophysiol. 2005, 94, 4082–4093. [Google Scholar] [CrossRef]
- Ross, B.; Herdman, A.T.; Pantev, C. Right hemispheric laterality of human 40 Hz auditory steady-state responses. Cereb. Cortex 2005, 15, 2029–2039. [Google Scholar] [CrossRef]
- Ross, B. A novel type of auditory responses: Temporal dynamics of 40-Hz steady-state responses induced by changes in sound localization. J. Neurophysiol. 2008, 100, 1265–1277. [Google Scholar] [CrossRef]
- Kuriki, S.; Kobayashi, Y.; Kobayashi, T.; Tanaka, K.; Uchikawa, Y. Steady-state MEG responses elicited by a sequence of amplitude-modulated short tones of different carrier frequencies. Hear. Res. 2013, 296, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Ross, B.; Fujioka, T. 40-Hz oscillations underlying perceptual binding in young and older adults. Psychophysiology 2016, 53, 974–990. [Google Scholar] [CrossRef] [PubMed]
- Woods, D.L.; Stecker, G.C.; Rinne, T.; Herron, T.J.; Cate, A.D.; Yund, E.W.; Liao, I.; Kang, X. Functional maps of human auditory cortex: Effects of acoustic features and attention. PLoS ONE 2009, 4, e5183. [Google Scholar] [CrossRef] [PubMed]
- Bohorquez, J.; Ozdamar, O. Generation of the 40-Hz auditory steady-state response (ASSR) explained using convolution. Clin. Neurophysiol. 2008, 119, 2598–2607. [Google Scholar] [CrossRef] [PubMed]
- Althen, H.; Grimm, S.; Escera, C. Fast detection of unexpected sound intensity decrements as revealed by human evoked potentials. PLoS ONE 2011, 6, e28522. [Google Scholar] [CrossRef] [PubMed]
- Tiitinen, H.; May, P.; Reinikainen, K.; Naatanen, R. Attentive novelty detection in humans is governed by pre-attentive sensory memory. Nature 1994, 372, 90–92. [Google Scholar] [CrossRef] [PubMed]
- Takegata, R.; Heikkila, R.; Naatanen, R. Sound energy and the magnitude of change: Effects on mismatch negativity. Neuroreport 2011, 22, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Naatanen, R.; Pakarinen, S.; Rinne, T.; Takegata, R. The mismatch negativity (MMN): Towards the optimal paradigm. Clin. Neurophysiol. 2004, 115, 140–144. [Google Scholar] [CrossRef]
- Jankowiak, S.; Berti, S. Behavioral and event-related potential distraction effects with regularly occurring auditory deviants. Psychophysiology 2007, 44, 79–85. [Google Scholar] [CrossRef]
- Pakarinen, S.; Huotilainen, M.; Naatanen, R. The mismatch negativity (MMN) with no standard stimulus. Clin. Neurophysiol. 2010, 121, 1043–1050. [Google Scholar] [CrossRef]
- Tan, H.R.; Gross, J.; Uhlhaas, P.J. MEG-measured auditory steady-state oscillations show high test-retest reliability: A sensor and source-space analysis. NeuroImage 2015, 122, 417–426. [Google Scholar] [CrossRef]
- Kwon, J.S.; O’Donnell, B.F.; Wallenstein, G.V.; Greene, R.W.; Hirayasu, Y.; Nestor, P.G.; Hasselmo, M.E.; Potts, G.F.; Shenton, M.E.; McCarley, R.W. Gamma frequency-range abnormalities to auditory stimulation in schizophrenia. Arch. Gen. Psychiatry 1999, 56, 1001–1005. [Google Scholar] [CrossRef]
- Onitsuka, T.; Oribe, N.; Nakamura, I.; Kanba, S. Review of neurophysiological findings in patients with schizophrenia. Psychiatry Clin. Neurosci. 2013, 67, 461–470. [Google Scholar] [CrossRef]
- O’Donnell, B.F.; Vohs, J.L.; Krishnan, G.P.; Rass, O.; Hetrick, W.P.; Morzorati, S.L. Chapter 6-The auditory steady-state response (ASSR): A translational biomarker for schizophrenia. Suppl. Clin. Neurophysiol. 2013, 62, 101–112. [Google Scholar]
- Koerner, T.K.; Zhang, Y. Effects of background noise on inter-trial phase coherence and auditory N1-P2 responses to speech stimuli. Hear. Res. 2015, 328, 113–119. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Motomura, E.; Inui, K.; Kawano, Y.; Nishihara, M.; Okada, M. Effects of Sound-Pressure Change on the 40 Hz Auditory Steady-State Response and Change-Related Cerebral Response. Brain Sci. 2019, 9, 203. https://doi.org/10.3390/brainsci9080203
Motomura E, Inui K, Kawano Y, Nishihara M, Okada M. Effects of Sound-Pressure Change on the 40 Hz Auditory Steady-State Response and Change-Related Cerebral Response. Brain Sciences. 2019; 9(8):203. https://doi.org/10.3390/brainsci9080203
Chicago/Turabian StyleMotomura, Eishi, Koji Inui, Yasuhiro Kawano, Makoto Nishihara, and Motohiro Okada. 2019. "Effects of Sound-Pressure Change on the 40 Hz Auditory Steady-State Response and Change-Related Cerebral Response" Brain Sciences 9, no. 8: 203. https://doi.org/10.3390/brainsci9080203
APA StyleMotomura, E., Inui, K., Kawano, Y., Nishihara, M., & Okada, M. (2019). Effects of Sound-Pressure Change on the 40 Hz Auditory Steady-State Response and Change-Related Cerebral Response. Brain Sciences, 9(8), 203. https://doi.org/10.3390/brainsci9080203