
Physiological Roles of Red Carrot Methanolic Extract and Vitamin E to Abrogate Cadmium-Induced Oxidative Challenge and Apoptosis in Rat Testes: Involvement of the Bax/Bcl-2 Ratio

 , , , ,
, , , ,  ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Preparation of Red Carrot Methanolic Extract
2.3. Phytochemical Analysis and the Assay of In Vitro Antioxidant Activities of Red Carrot Methanolic Extract (RCME)
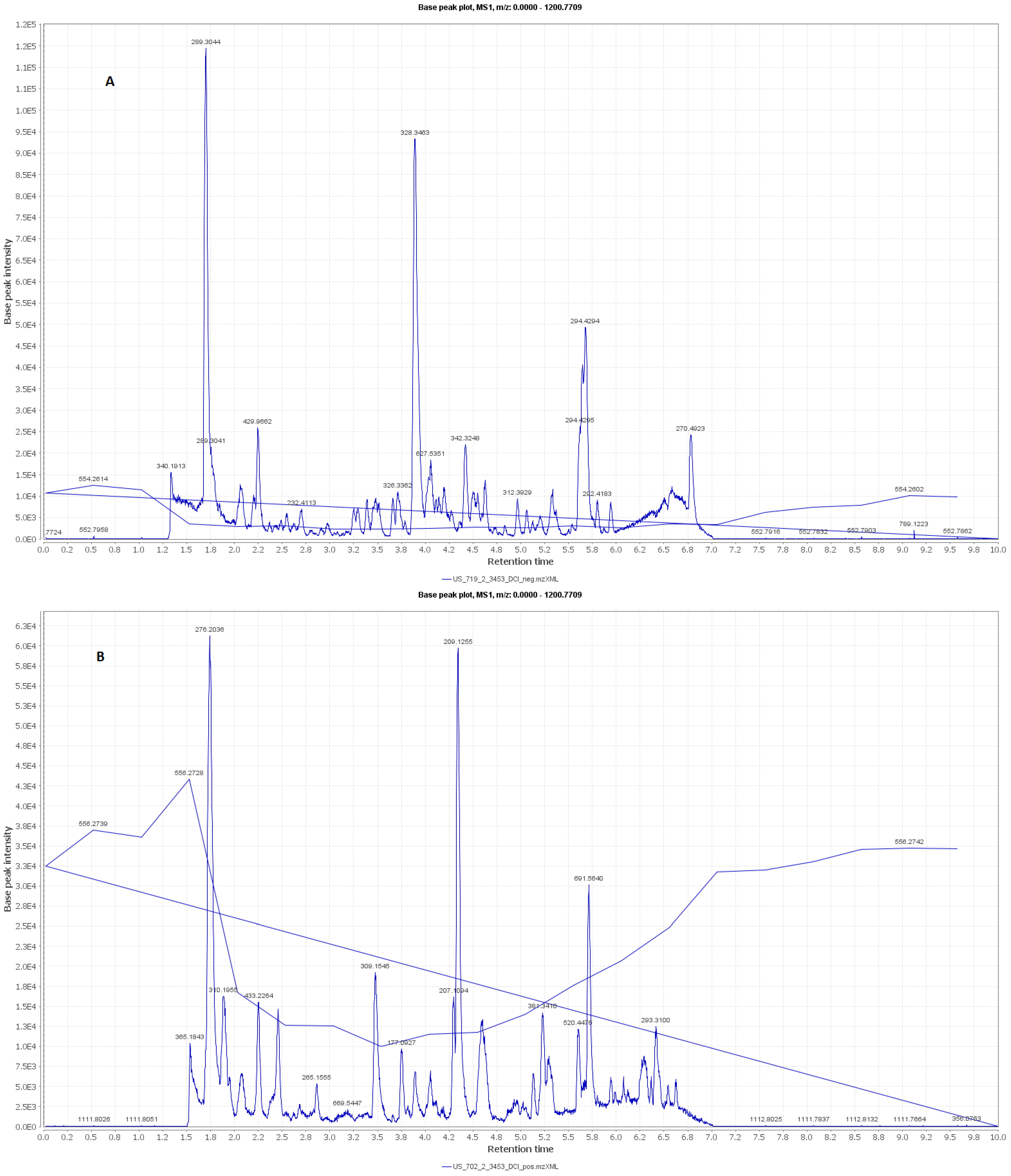
2.4. LC–MS Analysis
2.5. Animals and Experimental Design
2.6. Body Weight Gain, Testes Weights and Gonadosomatic Index
2.7. Serum and Tissue Samples
2.8. Epididymal Semen Samples Collection and Evaluation
2.9. Hormonal Assay
2.10. Determination of Oxidative Stress Markers
2.11. Determination of mRNA Relative Expression Levels of Bcl-2 and Bax Genes in Testicular Tissues by Real Time Quantitative PCR (RT-qPCR):
2.12. Histopathological Examination
2.13. Statistical Analysis
3. Results
3.1. Phytochemical Analysis and Assay of In Vitro Antioxidant Activities of the Red Carrot Methanolic Extract
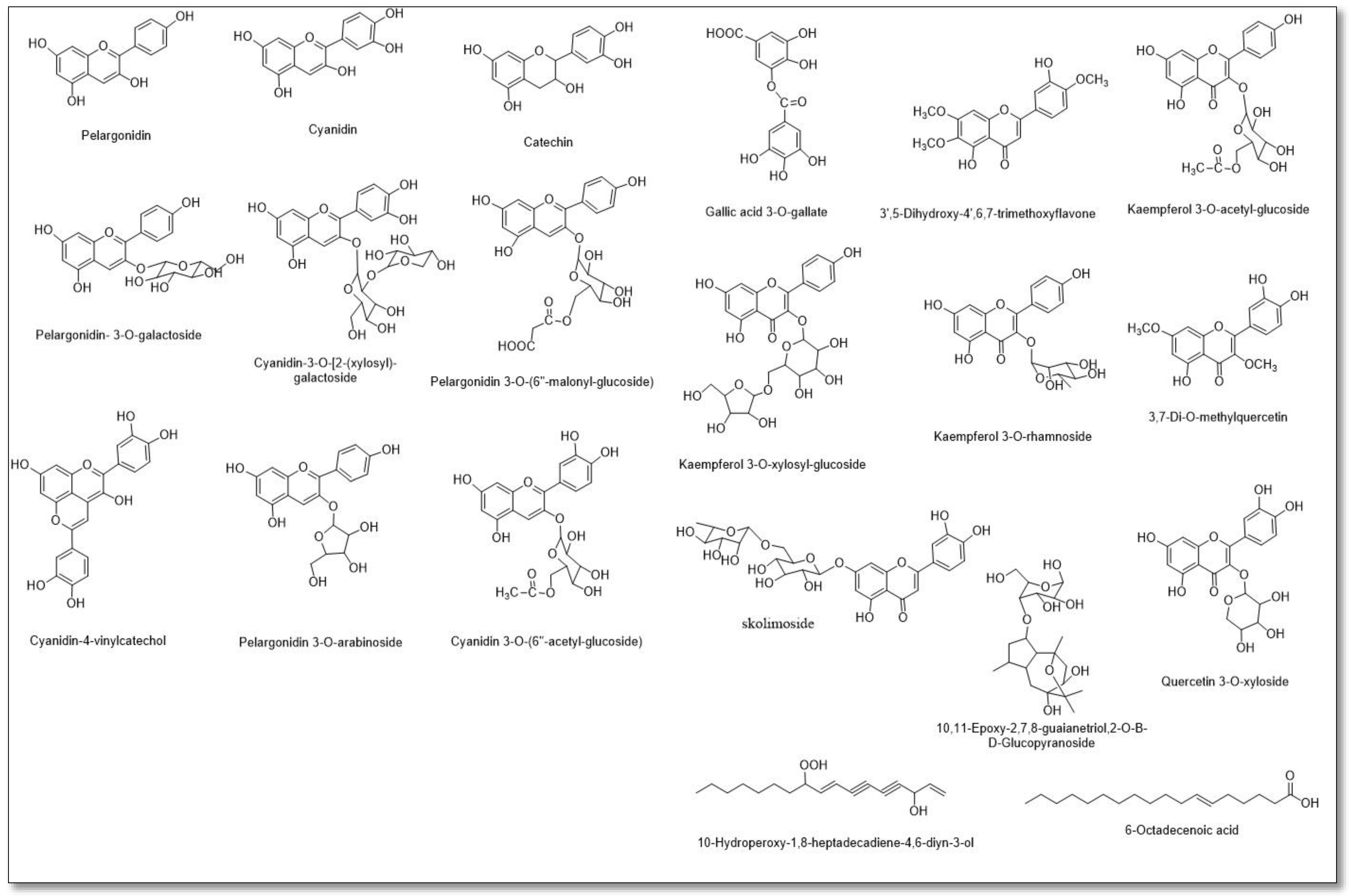
3.2. LC–MS Metabolomic Analysis of the Red Carrot Methanolic Extract
3.3. Means of Body Weight Gain, Testes Weights and Gonadosomatic Index in Control and Treated Rats
3.4. The Status of Semen Quality in Control and Treated Rats
3.5. Serum Levels of Male Reproductive Hormones (FSH, LH and Testosterone) in Control and Treated Rats
3.6. Testicular Oxidant/Antioxidant Status in Control and Treated Rats
3.7. The Testicular mRNA Relative Expression Levels of Bax and Bcl-2 in Control and Treated Rats
3.8. Histopathological Findings in Testicular, Prostatic and Semen Vesicle Glandular Structures in Control and Treated Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cavallini, G. Environmental Pollution and Infertility. In Clinical Management of Male Infertility; Springer: Berlin/Heidelberg, Germany, 2015; pp. 165–171. [Google Scholar]
- Lovaković, B.T. Cadmium, arsenic, and lead: Elements affecting male reproductive health. Curr. Opin. Toxicol. 2020, 19, 7–14. [Google Scholar] [CrossRef]
- Faroon, O.; Ashizawa, A.; Wright, S.; Tucker, P.; Jenkins, K.; Ingerman, L.; Rudisill, C. Toxicological profile of cadmium, agency for toxic substances and disease registry. Atlanta 2012, 273–274. [Google Scholar]
- Li, H.; Wallin, M.; Barregard, L.; Sallsten, G.; Lundh, T.; Ohlsson, C.; Mellström, D.; Andersson, E.M. Smoking-induced risk of osteoporosis is partly mediated by cadmium from tobacco smoke: The MrOS Sweden Study. J. Bone Miner. Res. 2020, 35, 1424–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satarug, S. Dietary cadmium intake and its effects on kidneys. Toxics 2018, 6, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chunhabundit, R. Cadmium exposure and potential health risk from foods in contaminated area, Thailand. Toxicol. Res. 2016, 32, 65–72. [Google Scholar] [CrossRef]
- WHO. Air Quality Guidelines for Europe; WHO Regional Office for Europe: Copenhagen, Denmark, 2000; ISBN 9289013583. [Google Scholar]
- Genchi, G.; Sinicropi, M.S.; Lauria, G.; Carocci, A.; Catalano, A. The effects of cadmium toxicity. Int. J. Environ. Res. Public Health 2020, 17, 3782. [Google Scholar] [CrossRef]
- Stukenborg, J.-B.; Kjartansdóttir, K.R.; Reda, A.; Colon, E.; Albersmeier, J.P.; Söder, O. Male germ cell development in humans. Horm. Res. Paediatr. 2014, 81, 2–12. [Google Scholar] [CrossRef]
- Hikim, A.P.S.; Swerdloff, R.S. Hormonal and genetic control of germ cell apoptosis in the testis. Rev. Reprod. 1999, 4, 38–47. [Google Scholar] [CrossRef]
- Amanpour, P.; Khodarahmi, P.; Salehipour, M. Protective effects of vitamin E on cadmium-induced apoptosis in rat testes. Naunyn. Schmiedebergs. Arch. Pharmacol. 2020, 393, 349–358. [Google Scholar] [CrossRef]
- Yang, Q.; Zhu, J.; Luo, X.; Li, F.; Cong, L.; Wang, Y.; Sun, Y. Melatonin attenuates cadmium-induced ovulatory dysfunction by suppressing endoplasmic reticulum stress and cell apoptosis. Reprod. Biol. Endocrinol. 2019, 17, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Angelis, C.; Galdiero, M.; Pivonello, C.; Salzano, C.; Gianfrilli, D.; Piscitelli, P.; Lenzi, A.; Colao, A.; Pivonello, R. The environment and male reproduction: The effect of cadmium exposure on reproductive function and its implication in fertility. Reprod. Toxicol. 2017, 73, 105–127. [Google Scholar] [CrossRef]
- Babaknejad, N.; Bahrami, S.; Moshtaghie, A.A.; Nayeri, H.; Rajabi, P.; Iranpour, F.G. Cadmium testicular toxicity in male Wistar rats: Protective roles of zinc and magnesium. Biol. Trace Elem. Res. 2018, 185, 106–115. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, H.; Lin, S.; Wang, K.; Wang, H.; Liu, Z. Protective effect of naringenin against cadmium-induced testicular toxicity in male SD rats. J. Inorg. Biochem. 2021, 214, 111310. [Google Scholar] [CrossRef]
- Chandel, M.; Jain, G.C. Toxic effects of transition metals on male reproductive system: A review. J. Env. Occup. Sci. Oct.-Dec. 2014, 3, 205. [Google Scholar] [CrossRef] [Green Version]
- Almeer, R.S.; Soliman, D.; Kassab, R.B.; AlBasher, G.I.; Alarifi, S.; Alkahtani, S.; Ali, D.; Metwally, D.; Abdel Moneim, A.E. Royal jelly abrogates cadmium-induced oxidative challenge in mouse testes: Involvement of the Nrf2 pathway. Int. J. Mol. Sci. 2018, 19, 3979. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.-H.; He, J.-B.; Yu, L.-H.; Li, L.; Long, M.; Liu, M.-D.; Li, P. Protective role of curcumin in cadmium-induced testicular injury in mice by attenuating oxidative stress via Nrf2/ARE pathway. Environ. Sci. Pollut. Res. 2019, 26, 34575–34583. [Google Scholar] [CrossRef] [PubMed]
- Owumi, S.E.; Adedara, I.A.; Duro-Ladipo, A.; Farombi, E.O. Acute diethyl nitrosamine and cadmium co-exposure exacerbates deficits in endocrine balance, sperm characteristics and antioxidant defence mechanisms in testes of pubertal rats. Andrologia 2019, 51, e13230. [Google Scholar] [CrossRef]
- Bjørneboe, A.; Bjørneboe, G.-E.A.; Drevon, C.A. Absorption, transport and distribution of vitamin E. J. Nutr. 1990, 120, 233–242. [Google Scholar] [CrossRef]
- Khorramabadi, K.M.; Talebi, A.R.; Sarcheshmeh, A.A.; Mirjalili, A. Protective effect of vitamin E on oxidative stress and sperm apoptosis in diabetic Mice. Int. J. Reprod. Biomed. 2019, 17, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Jelodar, G.; Akbari, A.; Parvaeei, P.; Nazifi, S. Vitamin E protects rat testis, eye and erythrocyte from oxidative stress during exposure to radiofrequency wave generated by a BTS antenna model. Int. J. Radiat. Res. 2018, 16, 217–224. [Google Scholar]
- Paunović, M.G.; Matić, M.M.; Ognjanović, B.I.; Saičić, Z.S. Antioxidative and haematoprotective activity of coenzyme Q10 and vitamin E against cadmium-induced oxidative stress in Wistar rats. Toxicol. Ind. Health 2017, 33, 746–756. [Google Scholar] [CrossRef]
- Kumar, A.; Pandey, R.; Siddiqi, N.J. Oxidative stress biomarkers of cadmium toxicity in mammalian systems and their distinct ameliorative strategy. J. Appl. Biotechnol. Bioeng. 2019, 6, 126–135. [Google Scholar]
- Sun, T.; Simon, P.W.; Tanumihardjo, S.A. Antioxidant phytochemicals and antioxidant capacity of biofortified carrots (Daucus carota L.) of various colors. J. Agric. Food Chem. 2009, 57, 4142–4147. [Google Scholar] [CrossRef]
- Bystrická, J.; Kavalcová, P.; Musilová, J.; VOLLMANNOVÁ, A.; Tomáš, T.; LENKOVÁ, M. Carrot (Daucus carota L. ssp. sativus (Hoffm.) Arcang.) as source of antioxidants. Acta Agric. Slov. 2015, 105, 303–311. [Google Scholar] [CrossRef]
- da Silva Dias, J.C. Nutritional and health benefits of carrots and their seed extracts. Food Nutr. Sci. 2014, 5, 2147. [Google Scholar] [CrossRef] [Green Version]
- Leja, M.; Kamińska, I.; Kramer, M.; Maksylewicz-Kaul, A.; Kammerer, D.; Carle, R.; Baranski, R. The content of phenolic compounds and radical scavenging activity varies with carrot origin and root color. Plant Foods Hum. Nutr. 2013, 68, 163–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zykevičiūtė-Laugks, J.; Česonienė, L.; Karklelienė, R. The influence of the sample preparation of carrots (Daucus carota L. Neptun) on the antioxidant activity and phenolic compounds. Biologija 2013, 59, 2. [Google Scholar] [CrossRef] [Green Version]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1999; Volume 299, pp. 152–178. ISBN 0076-6879. [Google Scholar]
- Benariba, N.; Djaziri, R.; Bellakhdar, W.; Belkacem, N.; Kadiata, M.; Malaisse, W.J.; Sener, A. Phytochemical screening and free radical scavenging activity of Citrullus colocynthis seeds extracts. Asian Pac. J. Trop. Biomed. 2013, 3, 35–40. [Google Scholar] [CrossRef] [Green Version]
- Jain, A.; Roy, S.; Joshi, A.; Joshi, N. Evaluation of in-vitro cytotoxic and antioxidant activity of methanolic extracts of Ipomoea carnea and Alternanthera sessilis. Int. J. Bioassays 2016, 5, 4763–4766. [Google Scholar] [CrossRef]
- Jan, S.; Khan, M.R.; Rashid, U.; Bokhari, J. Assessment of antioxidant potential, total phenolics and flavonoids of different solvent fractions of Monotheca buxifolia fruit. Osong Public Health Res. Perspect. 2013, 4, 246–254. [Google Scholar] [CrossRef] [Green Version]
- Hassanin, K.M.A.; Safwat, G.M. Role of Nigella Sativa oil in alleviating increased reproductive hormones and some inflammatory mediators caused by cadmium toxicity in rat testes. Int. J. Adv. Res. 2014, 2, 606–611. [Google Scholar]
- Layachi, N.; Kechrid, Z. Combined protective effect of vitamins C and E on cadmium induced oxidative liver injury in rats. Afr. J. Biotechnol. 2012, 11, 16013–16020. [Google Scholar]
- Sodimbaku, V.; Pujari, L.; Mullangi, R.; Marri, S. Carrot (Daucus carota L.): Nephroprotective against gentamicin-induced nephrotoxicity in rats. Indian J. Pharmacol. 2016, 48, 122. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Xie, S.; Chen, Z.; Wang, F.; Chen, K.; Zuo, Z.; Cui, H.; Guo, H.; Ouyang, P.; Chen, Z. Protective Effect of Vitamin E on Cadmium-Induced Renal Oxidative Damage and Apoptosis in Rats. Biol. Trace Elem. Res. 2021, 199, 4675–4687. [Google Scholar]
- Nouri, H.S.; Azarmi, Y.; Movahedin, M. Effect of growth hormone on testicular dysfunction induced by methotrexate in rats. Andrologia 2009, 41, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Abdoli, A.; Dalimi, A.; Movahedin, M. Impaired reproductive function of male rats infected with Toxoplasma gondii. Andrologia 2012, 44, 679–687. [Google Scholar] [CrossRef]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Elsevier Health Sciences: Amsterdam, The Netherlands, 2008; ISBN 0443102791. [Google Scholar]
- Soto-Vaca, A.; Gutierrez, A.; Losso, J.N.; Xu, Z.; Finley, J.W. Evolution of phenolic compounds from color and flavor problems to health benefits. J. Agric. Food Chem. 2012, 60, 6658–6677. [Google Scholar] [CrossRef]
- Montilla, E.C.; Arzaba, M.R.; Hillebrand, S.; Winterhalter, P. Anthocyanin composition of black carrot (Daucus carota ssp. sativus var. atrorubens Alef.) cultivars Antonina, Beta Sweet, Deep Purple, and Purple Haze. J. Agric. Food Chem. 2011, 59, 3385–3390. [Google Scholar] [CrossRef] [PubMed]
- Harborne, J.B. The Flavonoids: Advances in Research Since 1980; Springer: Berlin/Heidelberg, Germany, 2013; ISBN 1489929134. [Google Scholar]
- Poulin, M.J.; Bel-Rhlid, R.; Piché, Y.; Chenevert, R. Flavonoids released by carrot (Daucus carota) seedlings stimulate hyphal development of vesicular-arbuscular mycorrhizal fungi in the presence of optimal CO 2 enrichment. J. Chem. Ecol. 1993, 19, 2317–2327. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Zhang, L.; Yi, T.; Feng, Y.; Tian, J. Guaiane type sesquiterpenes and other constituents from Daucus carota L. Biochem. Syst. Ecol. 2010, 38, 309–312. [Google Scholar] [CrossRef]
- Pferschy-Wenzig, E.-M.; Getzinger, V.; Kunert, O.; Woelkart, K.; Zahrl, J.; Bauer, R. Determination of falcarinol in carrot (Daucus carota L.) genotypes using liquid chromatography/mass spectrometry. Food Chem. 2009, 114, 1083–1090. [Google Scholar] [CrossRef]
- Gregor, H.-D. Lipid composition of Daucus carota roots. Phytochemistry 1977, 16, 953–955. [Google Scholar] [CrossRef]
- Mouro, V.G.S.; Siman, V.A.; da Silva, J.; Dias, F.C.R.; Damasceno, E.M.; do Carmo Cupertino, M.; de Melo, F.C.S.A.; da Matta, S.L.P. Cadmium-induced testicular toxicity in mice: Subacute and subchronic route-dependent effects. Biol. Trace Elem. Res. 2020, 193, 466–482. [Google Scholar] [CrossRef]
- Austin, D.F. Medicinal plants of the world. An illustrated scientific guide to important medicinal plants and their uses. Econ. Bot. 2004, 58, 505. [Google Scholar] [CrossRef]
- Koley, T.K.; Srivastava, S.; Tripathi, Y.B.; Banerjee, K.; Oulkar, D.; Goon, A.; Tripathi, A.; Singh, B. High-resolution LCMS profiling of phenolic compounds of Indian black carrot and evaluation of its effect on antioxidant defense and glucose metabolism in animal model. Agric. Res. 2019, 8, 481–489. [Google Scholar] [CrossRef]
- Noda, Y.; Kaneyuki, T.; Mori, A.; Packer, L. Antioxidant activities of pomegranate fruit extract and its anthocyanidins: Delphinidin, cyanidin, and pelargonidin. J. Agric. Food Chem. 2002, 50, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Claudio, S.R.; Gollucke, A.P.B.; Yamamura, H.; Morais, D.R.; Bataglion, G.A.; Eberlin, M.N.; Peres, R.C.; Oshima, C.T.F.; Ribeiro, D.A. Purple carrot extract protects against cadmium intoxication in multiple organs of rats: Genotoxicity, oxidative stress and tissue morphology analyses. J. Trace Elem. Med. Biol. 2016, 33, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Abarikwu, S.O.; Simple, G.; Onuoha, C.S. Morphometric Evaluation of the Seminiferous Tubules and the Antioxidant Protective Effects of Gallic Acid and Quercetin in the Testis and Liver of Butyl Phthalate Treated Rats. Indian J. Clin. Biochem. 2020, 35, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Nna, V.U.; Ujah, G.A.; Mohamed, M.; Etim, K.B.; Igba, B.O.; Augustine, E.R.; Osim, E.E. Cadmium chloride–induced testicular toxicity in male wistar rats; prophylactic effect of quercetin, and assessment of testicular recovery following cadmium chloride withdrawal. Biomed. Pharmacother. 2017, 94, 109–123. [Google Scholar] [CrossRef]
- Bisht, S.; Faiq, M.; Tolahunase, M.; Dada, R. Oxidative stress and male infertility. Nat. Rev. Urol. 2017, 14, 470. [Google Scholar] [CrossRef] [PubMed]
- Henkel, R.; Samanta, L.; Agarwal, A. Oxidants, Antioxidants, and Impact of the Oxidative Status in Male Reproduction; Academic Press: Cambridge, MA, USA, 2018; ISBN 0128125675. [Google Scholar]
- Faraji, T.; Momeni, H.R.; Malmir, M. Protective effects of silymarin on testis histopathology, oxidative stress indicators, antioxidant defence enzymes and serum testosterone in cadmium-treated mice. Andrologia 2019, 51, e13242. [Google Scholar] [CrossRef] [PubMed]
- Samadder, A.; Tarafdar, D.; Das, R.; Khuda-Bukhsh, A.R.; Abraham, S.K. Efficacy of nanoencapsulated pelargonidin in ameliorating pesticide toxicity in fish and L6 cells: Modulation of oxidative stress and signalling cascade. Sci. Total Environ. 2019, 671, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yao, Z.; Yang, D.; Jiang, X.; Sun, J.; Tian, L.; Hu, J.; Wu, B.; Bai, W. Cyanidin-3-O-glucoside restores spermatogenic dysfunction in cadmium-exposed pubertal mice via histone ubiquitination and mitigating oxidative damage. J. Hazard. Mater. 2020, 387, 121706. [Google Scholar] [CrossRef] [PubMed]
- Acquaviva, R.; Russo, A.; Galvano, F.; Galvano, G.; Barcellona, M.L.; Volti, G.L.; Vanella, A. Cyanidin and cyanidin 3-O-β-D-glucoside as DNA cleavage protectors and antioxidants. Cell Biol. Toxicol. 2003, 19, 243–252. [Google Scholar] [CrossRef]
- Sharath Babu, G.R.; Anand, T.; Ilaiyaraja, N.; Khanum, F.; Gopalan, N. Pelargonidin modulates Keap1/Nrf2 pathway gene expression and ameliorates citrinin-induced oxidative stress in HepG2 cells. Front. Pharmacol. 2017, 8, 868. [Google Scholar] [CrossRef]
- Kalender, Y.; Kaya, S.; Durak, D.; Uzun, F.G.; Demir, F. Protective effects of catechin and quercetin on antioxidant status, lipid peroxidation and testis-histoarchitecture induced by chlorpyrifos in male rats. Environ. Toxicol. Pharmacol. 2012, 33, 141–148. [Google Scholar] [CrossRef]
- Blanco, A.; Moyano, R.; Vivo, J.; Flores-Acuña, R.; Molina, A.; Blanco, C.; Agüera, E.; Monterde, J.G. Quantitative changes in the testicular structure in mice exposed to low doses of cadmium. Environ. Toxicol. Pharmacol. 2007, 23, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Atere, T.G.; Akinloye, O.A. High dose of standardised extract of Costus afer leaves potentiates cadmium reproductive toxicity in Wistar rats. Andrologia 2019, 51, e13360. [Google Scholar] [CrossRef]
- Ahmed, M.M.; El-Shazly, S.A.; Alkafafy, M.E.; Mohamed, A.A.; Mousa, A.A. Protective potential of royal jelly against cadmium-induced infertility in male rats. Andrologia 2018, 50, e12996. [Google Scholar] [CrossRef]
- Siu, E.R.; Mruk, D.D.; Porto, C.S.; Cheng, C.Y. Cadmium-induced testicular injury. Toxicol. Appl. Pharmacol. 2009, 238, 240–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kini, R.D.; Tripathi, Y.; Raghuveer, C.V.; Pai, S.R.; Ramswamy, C.; Nayanatara, A.K.; Vinodhini, N.A.; Ranade, A. Protective role of vitamin E against cadmium chloride induced testicular damage in rats. J. Physiol. Biomed. Sci. 2009, 22, 12–16. [Google Scholar]
- Wang, J.; Zhu, H.; Wang, K.; Yang, Z.; Liu, Z. Protective effect of quercetin on rat testes against cadmium toxicity by alleviating oxidative stress and autophagy. Environ. Sci. Pollut. Res. Int. 2020, 27, 25278–25286. [Google Scholar] [CrossRef] [PubMed]
- Saha, R.; Roychoudhury, S.; Kar, K.; Varghese, A.C.; Nandi, P.; Sharma, G.D.; Formicki, G.; Slama, P.; Kolesarova, A. Coenzyme Q10 ameliorates cadmium induced reproductive toxicity in male rats. Physiol. Res. 2019, 68, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Marettová, E.; Maretta, M.; Legáth, J. Toxic effects of cadmium on testis of birds and mammals: A review. Anim. Reprod. Sci. 2015, 155, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Vernet, P.; Aitken, R.J.; Drevet, J.R. Antioxidant strategies in the epididymis. Mol. Cell. Endocrinol. 2004, 216, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Acharya, U.R.; Mishra, M.; Patro, J.; Panda, M.K. Effect of vitamins C and E on spermatogenesis in mice exposed to cadmium. Reprod. Toxicol. 2008, 25, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Agdam, H.R.; Razi, M.; Amniattalab, A.; Malekinejad, H.; Molavi, M. Co-Administration of vitamin E and testosterone attenuates the atrazine-induced toxic effects on sperm quality and testes in rats. Cell J. 2017, 19, 292. [Google Scholar]
- Alizadeh, H.; Khaki, A.; Farzadi, L.; Nouri, M.; Ahmadi, A.Y.; Seyedghiasi, G.; Shahnazi, V. The therapeutic effects of a medicinal plant mixture in capsule form on catalase levels in the semen of men with oligospermia. Crescent J. Med. Biol. Sci. 2015, 2, 6–9. [Google Scholar]
- Jiang, X.; Zhu, C.; Li, X.; Sun, J.; Tian, L.; Bai, W. Cyanidin-3-O-glucoside at low doses protected against 3-Chloro-1, 2-propanediol induced testis injury and improved spermatogenesis in male rats. J. Agric. Food Chem. 2018, 66, 12675–12684. [Google Scholar] [CrossRef]
- Boonsorn, T.; Kongbuntad, W.; Narkkong, N.A.; Aengwanich, W. Effects of catechin addition to extender on sperm quality and lipid peroxidation in boar semen. Am. J. Agric. Environ. Sci. 2010, 7, 283–288. [Google Scholar]
- Mao, T.; Han, C.; Wei, B.; Zhao, L.; Zhang, Q.; Deng, R.; Liu, J.; Luo, Y.; Zhang, Y. Protective effects of quercetin against cadmium chloride-induced oxidative injury in goat sperm and zygotes. Biol. Trace Elem. Res. 2018, 185, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Imafidon, C.E.; Olatoye, T.R.; Bamidele, F.S.; Ojo, O.E.; Ademoye, K.A. Cadmium-induced testicular toxicity, oxidative stress and histopathology in Wistar rats: Sustained effects of polyphenol-rich extract of Vernonia amygdalina (Del.) leaf. J. Interdiscipl. Histopathol. 2016, 4, 54–62. [Google Scholar] [CrossRef]
- Wu, X.; Guo, X.; Wang, H.; Zhou, S.; Li, L.; Chen, X.; Wang, G.; Liu, J.; Ge, H.-S.; Ge, R.-S. A brief exposure to cadmium impairs Leydig cell regeneration in the adult rat testis. Sci. Rep. 2017, 7, 6337. [Google Scholar] [CrossRef] [Green Version]
- Gunnarsson, D.; Nordberg, G.; Selstam, G. Differential effects of cadmium on the gene expression of seven-transmembrane-spanning receptors and GAPDH in the rat testis. Toxicol. Lett. 2007, 168, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Tena-Sempere, M. Kisspeptin/GPR54 system as potential target for endocrine disruption of reproductive development and function. Int. J. Androl. 2010, 33, 360–368. [Google Scholar] [CrossRef] [PubMed]
- de Souza Predes, F.; Diamante, M.A.S.; Dolder, H. Testis response to low doses of cadmium in Wistar rats. Int. J. Exp. Pathol. 2010, 91, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Anahara, R.; Toyama, Y.; Mori, C. Review of the histological effects of the anti-androgen, flutamide, on mouse testis. Reprod. Toxicol. 2008, 25, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Guo, J.; Jiang, X.; Sun, J.; Tian, L.; Jiao, R.; Tang, Y.; Bai, W. Cyanidin-3-O-glucoside protects against cadmium-induced dysfunction of sex hormone secretion via the regulation of hypothalamus-pituitary-gonadal axis in male pubertal mice. Food Chem. Toxicol. 2019, 129, 13–21. [Google Scholar] [CrossRef]
- Ujah, G.A.; Nna, V.U.; Agah, M.I.; Omue, L.O.; Leku, C.B.; Osim, E.E. Effect of quercetin on cadmium chloride-induced impairments in sexual behaviour and steroidogenesis in male Wistar rats. Andrologia 2018, 50, e12866. [Google Scholar] [CrossRef]
- Yu, P.; Pu, H.; Chen, S.; Wang, S.; Wang, P.S. Effects of catechin, epicatechin and epigallocatechin gallate on testosterone production in rat leydig cells. J. Cell. Biochem. 2010, 110, 333–342. [Google Scholar] [CrossRef]
- Rossé, T.; Olivier, R.; Monney, L.; Rager, M.; Conus, S.; Fellay, I.; Jansen, B.; Borner, C. Bcl-2 prolongs cell survival after Bax-induced release of cytochrome c. Nature 1998, 391, 496–499. [Google Scholar] [CrossRef]
- Eleawa, S.M.; Alkhateeb, M.A.; Alhashem, F.H.; Bin-Jaliah, I.; Sakr, H.F.; Elrefaey, H.M.; Elkarib, A.O.; Alessa, R.M.; Haidara, M.A.; Shatoor, A.S. Resveratrol reverses cadmium chloride-induced testicular damage and subfertility by downregulating p53 and Bax and upregulating gonadotropins and Bcl-2 gene expression. J. Reprod. Dev. 2014, 60, 115–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, R.; Zheng, S.; Wang, X. Selenium protects against cadmium-induced kidney apoptosis in chickens by activating the PI3K/AKT/Bcl-2 signaling pathway. Environ. Sci. Pollut. Res. 2017, 24, 20342–20353. [Google Scholar] [CrossRef] [PubMed]
- Brüne, B.; von Knethen, A.; Sandau, K.B. Nitric oxide (NO): An effector of apoptosis. Cell Death Differ. 1999, 6, 969–975. [Google Scholar] [CrossRef] [Green Version]
- Jahan, S.; Zahra, A.; Irum, U.; Iftikhar, N.; Ullah, H. Protective effects of different antioxidants against cadmium induced oxidative damage in rat testis and prostate tissues. Syst. Biol. Reprod. Med. 2014, 60, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jiang, X.; Sun, J.; Zhu, C.; Li, X.; Tian, L.; Liu, L.; Bai, W. Cytoprotective effects of dietary flavonoids against cadmium-induced toxicity. Ann. N. Y. Acad. Sci. 2017, 1398, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Jayaprakasha, G.K.; Murthy, K.N.C.; Pellati, F.; Patil, B.S. BetaSweet carrot extracts have antioxidant activity and in vitro antiproliferative effects against breast cancer cells. J. Funct. Foods 2019, 62, 103552. [Google Scholar] [CrossRef]
- Kurilich, A.C.; Clevidence, B.A.; Britz, S.J.; Simon, P.W.; Novotny, J.A. Plasma and urine responses are lower for acylated vs. nonacylated anthocyanins from raw and cooked purple carrots. J. Agric. Food Chem. 2005, 53, 6537–6542. [Google Scholar] [CrossRef]
- Potter, A.S.; Foroudi, S.; Stamatikos, A.; Patil, B.S.; Deyhim, F. Drinking carrot juice increases total antioxidant status and decreases lipid peroxidation in adults. Nutr. J. 2011, 10, 96. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polarity | m/z | Rt | Formula | Name | Source |
|---|---|---|---|---|---|
| [M − H]− | 270.4925 | 6.8127 | C15H11O5 | Pelargonidin | [41] |
| [M + H]+ | 288.1700 | 2.103 | C15H11O6 | Cyanidin | [41,42,43] |
| [M + H]+ | 291.1863 | 6.100 | C15H14O6 | Catechin | [41] |
| [M + H]+ | 434.2948 | 3.018 | C21H21O10 | Pelargonidin- 3-O-galactoside | [42] |
| [M − H]− | 580.6404 | 9.84 | C26H29O15 | Cyanidin-3-O-[2-(xylosyl)-galactoside | [43] |
| [M + H]+ | 520.4487 | 5.719 | C24H23O13 | Pelargonidin 3-O-(6″-malonyl-glucoside) | [42] |
| [M − H]− | 490.858 | 4.571 | C23H23O12 | Cyanidin 3-O-(6″-acetylglucoside) | [42] |
| [M + H]+ | 404.2404 | 2.299 | C20H19O9 | Pelargonidin 3-O-arabinoside | [42] |
| [M + H]+ | 420.2739 | 2.734 | C23H15O8 | Cyanidin-4-vinylcatechol | [42] |
| [M − H]− | 323.1974 | 4.671 | C14H10O9 | Gallic acid 3-O-gallate | [28] |
| [M − H]− | 343.3254 | 4.604 | C18H16O7 | 3′,5-Dihydroxy-4′,6,7-trimethoxyflavone | [44] |
| [M + H]+ | 433.2267 | 2.474 | C21H20O10 | Kaempferol 3-O-rhamnoside | [44] |
| [M − H]− | 489.8591 | 4.429 | C23H22O12 | Kaempferol 3-O-acetyl-glucoside | [44] |
| [M − H]− | 579.6392 | 1.679 | C26H28O15 | Kaempferol 3-O-xylosyl-glucoside | [44] |
| [M + H]+ | 595.5058 | 6.241 | C27H30O15 | Skolimoside | [45] |
| [M − H]− | 329.3467 | 4.075 | C17H14O7 | 3,7-Di-O-methylquercetin | [44] |
| [M + H]+ | 435.4387 | 6.361 | C20H18O11 | Quercetin 3-O-xyloside | [44] |
| [M + H]+ | 433.2267 | 2.474 | C21H36O9 | 10,11-Epoxy-2,7,8-guaianetriol,2-O-β-D-glucopyranoside | [45] |
| [M + H]+ | 277.2078 | 1.6447 | C17H24O3 | 10-Hydroperoxy-1,8-heptadecadiene-4,6-diyn-3-ol | [46] |
| [M − H]− | 281.3715 | 4.023 | C18H34O2 | 6-Octadecenoic acid | [47] |
| Normal Control | CdCl2 | CdCl2 & Vit. E | CdCl2 & RCME | |
|---|---|---|---|---|
| Testosterone (ng/mL) | 3.89 ± 0.08 | 2.84 ± 0.13 a | 3.30 ± 0.12 a | 3.94 ± 0.2 b, c |
| LH (ng/mL) | 2.47 ± 0.22 | 1.22 ± 0.13 a | 1.81 ± 0.07 a, b | 2.28 ± 0.17 b |
| FSH (ng/mL) | 5.52 ± 0.23 | 8.21 ± 0.25 a | 6.15 ± 0.12 b | 5.80 ± 0.16 b |
| Glutathione (mg/g testicular tissues) | 4.59 ± 0.30 | 2.21 ± 0.16 a | 2.81 ± 0.07 a, b | 2.92 ± 0.06 a, b |
| Catalase (U/g testicular tissues) | 1.69 ± 0.10 | 1.25 ± 0.08 a | 1.53 ± 0.05 b | 1.51 ± 0.06 b |
| Superoxide dismutase (U/g testicular tissues) | 1.46 ± 0.06 | 1.11 ± 0.05 a | 1.44 ± 0.07 b | 1.44 ± 0.07 b |
| TAC (mmol/g testicular tissues) | 1.87 ± 0.12 | 1.09 ± 0.08 a | 1.85 ± 0.10 b | 1.58 ± 0.08 b |
| MDA (nmol/g testicular tissues) | 0.88 ± 0.04 | 2.12 ± 0.18 a | 1.61 ± 0.14 a, b | 1.5 ± 0.13 a, b |
| Nitric oxide (µmol/g testicular tissues) | 8.82 ± 0.23 | 12.26 ± 0.56 a | 10.64 ± 0.46 | 10.77 ± 0.50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel-Wahab, A.; Hassanin, K.M.A.; Mahmoud, A.A.; Abdel-Badeea, W.I.E.; Abdel-Razik, A.-R.H.; Attia, E.Z.; Abdelmohsen, U.R.; Abdel Aziz, R.L.; Najda, A.; Alanazi, I.S.; et al. Physiological Roles of Red Carrot Methanolic Extract and Vitamin E to Abrogate Cadmium-Induced Oxidative Challenge and Apoptosis in Rat Testes: Involvement of the Bax/Bcl-2 Ratio. Antioxidants 2021, 10, 1653. https://doi.org/10.3390/antiox10111653
Abdel-Wahab A, Hassanin KMA, Mahmoud AA, Abdel-Badeea WIE, Abdel-Razik A-RH, Attia EZ, Abdelmohsen UR, Abdel Aziz RL, Najda A, Alanazi IS, et al. Physiological Roles of Red Carrot Methanolic Extract and Vitamin E to Abrogate Cadmium-Induced Oxidative Challenge and Apoptosis in Rat Testes: Involvement of the Bax/Bcl-2 Ratio. Antioxidants. 2021; 10(11):1653. https://doi.org/10.3390/antiox10111653
Chicago/Turabian StyleAbdel-Wahab, Ahmed, Kamel M. A. Hassanin, Ahmed A. Mahmoud, Walaa I. E. Abdel-Badeea, Abdel-Razik H. Abdel-Razik, Eman Zekry Attia, Usama Ramadan Abdelmohsen, Rabie L. Abdel Aziz, Agnieszka Najda, Ibtesam S. Alanazi, and et al. 2021. "Physiological Roles of Red Carrot Methanolic Extract and Vitamin E to Abrogate Cadmium-Induced Oxidative Challenge and Apoptosis in Rat Testes: Involvement of the Bax/Bcl-2 Ratio" Antioxidants 10, no. 11: 1653. https://doi.org/10.3390/antiox10111653
APA StyleAbdel-Wahab, A., Hassanin, K. M. A., Mahmoud, A. A., Abdel-Badeea, W. I. E., Abdel-Razik, A. -R. H., Attia, E. Z., Abdelmohsen, U. R., Abdel Aziz, R. L., Najda, A., Alanazi, I. S., Alsharif, K. F., Abdel-Daim, M. M., & Mahmoud, M. O. (2021). Physiological Roles of Red Carrot Methanolic Extract and Vitamin E to Abrogate Cadmium-Induced Oxidative Challenge and Apoptosis in Rat Testes: Involvement of the Bax/Bcl-2 Ratio. Antioxidants, 10(11), 1653. https://doi.org/10.3390/antiox10111653