
Phenolic Compounds of Reynoutria sp. as Modulators of Oral Cavity Lactoperoxidase System

 ,
,  , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Preparation of Extracts
2.3. Assessment of the Effect of Extracts and Selected Polyphenols on the Production of OSCN– by LPO
2.4. Analysis of Kinetics via Stopped-Flow Method
2.5. Assessment of the Effect of Extracts on the Inhibition of S. mutans Growth by LPO
2.6. Determination of the MIC and MBC
2.7. Statistical Analysis
3. Results and Discussion
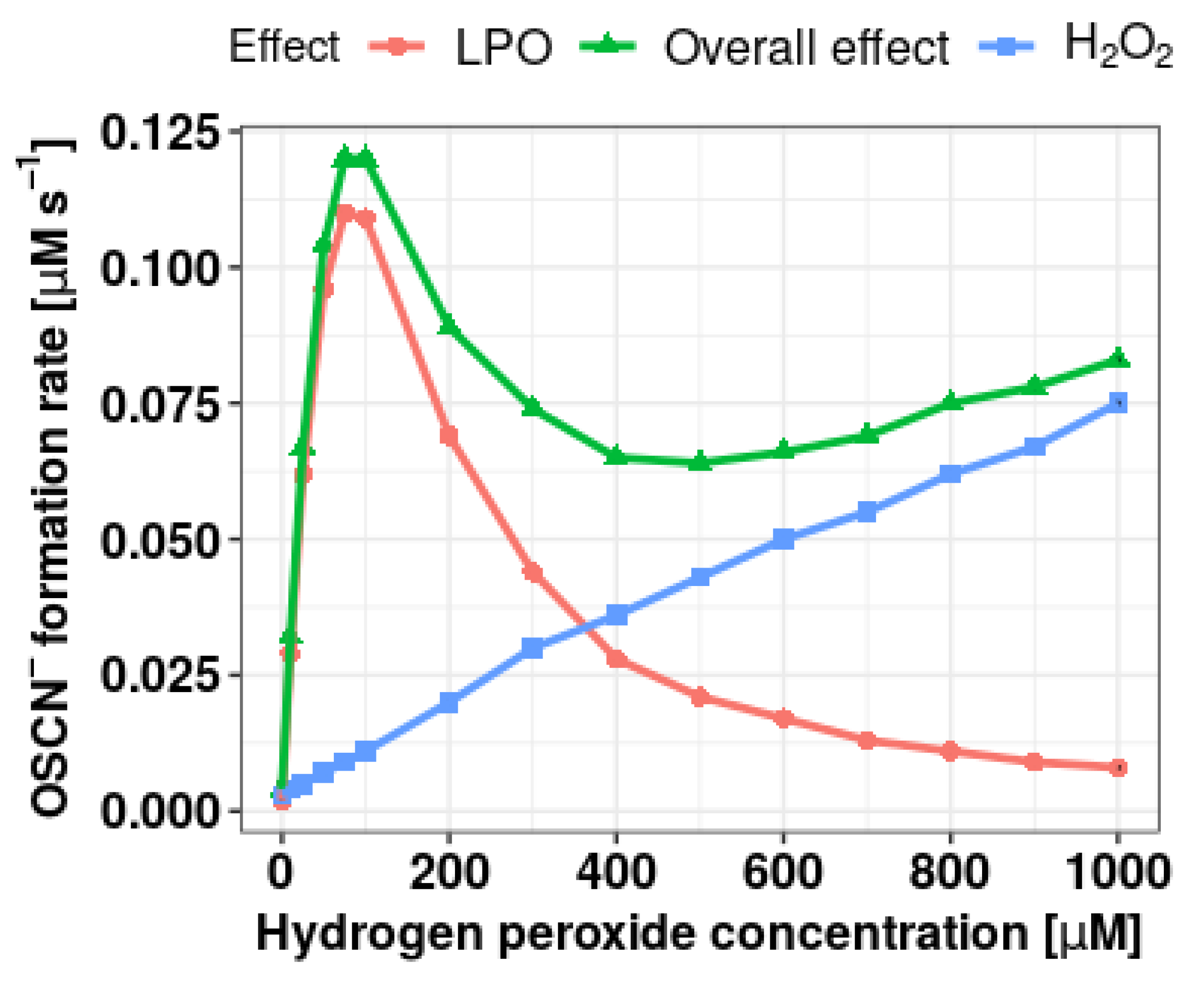
3.1. Selection of Conditions for Determining the Effect of Extracts on the Formation of OSCN– in the LPO Halogenation Cycle
3.2. Effect of Individual Extracts and Polyphenols on the Formation of OSCN– in the LPO Halogenation Cycle
3.3. Analysis of the Kinetics of the Reactions of Extracts, Fractions and Selected Polyphenols with Compound I and Compound II
3.4. Assessment of the Ability to Increase the Antimicrobial Activity of the LPO System
3.5. Antimicrobial Activity of Extracts from Reynoutria sp.
3.6. Physiological Relevance
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Magacz, M.; Kędziora, K.; Sapa, J.; Krzyściak, W. The Significance of Lactoperoxidase System in Oral Health: Application and Efficacy in Oral Hygiene Products. Int. J. Mol. Sci. 2019, 20, 1443. [Google Scholar] [CrossRef] [Green Version]
- Dua, S.; Maclean, M.J.; Fitzgerald, M.; McAnoy, A.M.; Bowie, J.H. Is the hypothiocyanite anion (OSCN)- the major product in the peroxidase catalyzed oxidation of the thiocyanate anion (SCN)-? A joint experimental and theoretical study. J. Phys. Chem. A 2006, 110, 4930–4936. [Google Scholar] [CrossRef]
- Ihalin, R.; Loimaranta, V.; Tenovuo, J. Origin, structure, and biological activities of peroxidases in human saliva. Arch. Biochem. Biophys. 2006, 445, 261–268. [Google Scholar] [CrossRef]
- Bafort, F.; Parisi, O.; Perraudin, J.P.; Jijakli, M.H. Mode of Action of Lactoperoxidase as Related to Its Antimicrobial Activity: A Review. Enzyme Res. 2014, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, D.T.; Barrett, T.J.; White, M.Y.; Cordwell, S.J.; Davies, M.J.; Hawkins, C.L. Cellular targets of the myeloperoxidase-derived oxidant hypothiocyanous acid (HOSCN) and its role in the inhibition of glycolysis in macrophages. Free Radic. Biol. Med. 2016, 94, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.L.; Aune, T.M. Lactoperoxidase, peroxide, thiocyanate antimicrobial system: Correlation of sulfhydryl oxidation with antimicrobial action. Infect. Immun. 1978, 20, 456–463. [Google Scholar] [CrossRef] [Green Version]
- Furtmüller, P.G.; Burner, U.; Jantschko, W.; Regelsberger, G.; Obinger, C. Two-electron reduction and one-electron oxidation of organic hydroperoxides by human myeloperoxidase. FEBS Lett. 2000, 484, 139–143. [Google Scholar] [CrossRef] [Green Version]
- Kussendrager, K.D.; van Hooijdonk, A.C. Lactoperoxidase: Physico-chemical properties, occurrence, mechanism of action and applications. Br. J. Nutr. 2000, 84 (Suppl. S1), S19–S25. [Google Scholar] [CrossRef] [Green Version]
- Sheikh, I.A.; Jiffri, E.H.; Ashraf, G.M.; Kamal, M.A.; Beg, M.A. Structural studies on inhibitory mechanisms of antibiotic, corticosteroid and catecholamine molecules on lactoperoxidase. Life Sci. 2018, 207, 412–419. [Google Scholar] [CrossRef]
- Köksal, Z.; Alim, Z. Lactoperoxidase, an antimicrobial enzyme, is inhibited by some indazoles. Drug Chem. Toxicol. 2020, 43, 22–26. [Google Scholar] [CrossRef]
- Sarikaya, S.B.O.; Sisecioglu, M.; Cankaya, M.; Gulcin, I.; Ozdemir, H. Inhibition profile of a series of phenolic acids on bovine lactoperoxidase enzyme. J. Enzyme Inhib. Med. Chem. 2015, 30, 479–483. [Google Scholar] [CrossRef]
- El-Benna, J.; Dang, P.M.-C.; Gougerot-Pocidalo, M.-A. Role of the NADPH oxidase systems Nox and Duox in host defense and inflammation. Expert Rev. Clin. Immunol. 2007, 3, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Geiszt, M.; Witta, J.; Baffi, J.; Lekstrom, K.; Leto, T.L. Dual oxidases represent novel hydrogen peroxide sources supporting mucosal surface host defense. FASEB J. 2003, 17, 1502–1504. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ge, X.; Dou, Y.; Wang, X.; Patel, J.R.; Xu, P. Identification of hydrogen peroxide production-related genes in Streptococcus sanguinis and their functional relationship with pyruvate oxidase. Microbiology 2011, 157, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Høiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flemming, H.-C.; Wuertz, S. Bacteria and archaea on Earth and their abundance in biofilms. Nat. Rev. Microbiol. 2019, 17, 247–260. [Google Scholar] [CrossRef]
- Folayan, M.O.; El Tantawi, M.; Ramos-Gomez, F.; Sabbah, W. Editorial: Country Profile of the Epidemiology and Clinical Management of Early Childhood Caries. Front. Public Health 2020, 8, 141. [Google Scholar] [CrossRef]
- Pitts, N.B.; Zero, D.T.; Marsh, P.D.; Ekstrand, K.; Weintraub, J.A.; Ramos-Gomez, F.; Tagami, J.; Twetman, S.; Tsakos, G.; Ismail, A. Dental caries. Nat. Rev. Dis. Prim. 2017, 3, 17030. [Google Scholar] [CrossRef] [Green Version]
- Gau, J.; Prévost, M.; Van Antwerpen, P.; Sarosi, M.-B.; Rodewald, S.; Arnhold, J.; Flemmig, J. Tannins and Tannin-Related Derivatives Enhance the (Pseudo-)Halogenating Activity of Lactoperoxidase. J. Nat. Prod. 2017, 80, 1328–1338. [Google Scholar] [CrossRef]
- Gau, J.; Arnhold, J.; Flemmig, J. Reactivation of peroxidase activity in human saliva samples by polyphenols. Arch. Oral Biol. 2018, 85, 70–78. [Google Scholar] [CrossRef]
- Upadhyay, S.; Dixit, M. Role of polyphenols and other phytochemicals on molecular signaling. Oxid. Med. Cell. Longev. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Shen, T.; Lou, H. Dietary Polyphenols and Their Biological Significance. Int. J. Mol. Sci. 2007, 8, 950–988. [Google Scholar] [CrossRef] [Green Version]
- Flemmig, J.; Rusch, D.; Czerwińska, M.E.; Rauwald, H.W.; Arnhold, J. Components of a standardised olive leaf dry extract (Ph. Eur.) promote hypothiocyanite production by lactoperoxidase. Arch. Biochem. Biophys. 2014, 549, 17–25. [Google Scholar] [CrossRef]
- Gau, J.; Furtmüller, P.G.; Obinger, C.; Prévost, M.; Van Antwerpen, P.; Arnhold, J.; Flemmig, J. Flavonoids as promoters of the (pseudo-)halogenating activity of lactoperoxidase and myeloperoxidase. Free Radic. Biol. Med. 2016, 97, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Gau, J.; Furtmüller, P.G.; Obinger, C.; Arnhold, J.; Flemmig, J. Enhancing hypothiocyanite production by lactoperoxidase—mechanism and chemical properties of promotors. Biochem. Biophys. Rep. 2015, 4, 257–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jug, U.; Naumoska, K.; Vovk, I. (-)-Epicatechin-An Important Contributor to the Antioxidant Activity of Japanese Knotweed Rhizome Bark Extract as Determined by Antioxidant Activity-Guided Fractionation. Antioxidants 2021, 10, 133. [Google Scholar] [CrossRef] [PubMed]
- Nawrot-Hadzik, I.; Granica, S.; Domaradzki, K.; Pecio, Ł.; Matkowski, A. Isolation and Determination of Phenolic Glycosides and Anthraquinones from Rhizomes of Various Reynoutria Species. Planta Med. 2018, 84, 1118–1126. [Google Scholar] [CrossRef] [Green Version]
- Song, J.-H.; Kim, S.-K.; Chang, K.-W.; Han, S.-K.; Yi, H.-K.; Jeon, J.-G. In vitro inhibitory effects of Polygonum cuspidatum on bacterial viability and virulence factors of Streptococcus mutans and Streptococcus sobrinus. Arch. Oral Biol. 2006, 51, 1131–1140. [Google Scholar] [CrossRef]
- Nawrot-Hadzik, I.; Slusarczyk, S.; Granica, S.; Hadzik, J.; Matkowski, A. Phytochemical diversity in rhizomes of three Reynoutria species and their antioxidant activity correlations elucidated by LC-ESI-MS/MS analysis. Molecules 2019, 24, 1136. [Google Scholar] [CrossRef] [Green Version]
- Nawrot-Hadzik, I.; Hadzik, J.; Fleischer, M.; Choromańska, A.; Sterczała, B.; Kubasiewicz-Ross, P.; Saczko, J.; Gałczyńska-Rusin, M.; Gedrange, T.; Matkowski, A. Chemical composition of east Asian invasive knotweeds, their cytotoxicity and antimicrobial efficacy against cariogenic pathogens: An in-vitro study. Med. Sci. Monit. 2019, 25, 3279–3287. [Google Scholar] [CrossRef]
- Du, Q.; Fu, M.; Zhou, Y.; Cao, Y.; Guo, T.; Zhou, Z.; Li, M.; Peng, X.; Zheng, X.; Li, Y.; et al. Sucrose promotes caries progression by disrupting the microecological balance in oral biofilms: An in vitro study. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Schlorke, D.; Flemmig, J.; Birkemeyer, C.; Arnhold, J. Formation of cyanogen iodide by lactoperoxidase. J. Inorg. Biochem. 2016, 154, 35–41. [Google Scholar] [CrossRef]
- Nagy, P.; Jameson, G.N.L.; Winterbourn, C.C. Kinetics and mechanisms of the reaction of hypothiocyanous acid with 5-thio-2-nitrobenzoic acid and reduced glutathione. Chem. Res. Toxicol. 2009, 22, 1833–1840. [Google Scholar] [CrossRef] [PubMed]
- Koksal, Z.; Kalin, R.; Kalin, P.; Karaman, M.; Gulcin, İ.; Ozdemir, H. Lactoperoxidase inhibition of some natural phenolic compounds: Kinetics and molecular docking studies. J. Food Biochem. 2020, 44, e13132. [Google Scholar] [CrossRef] [PubMed]
- Kilmister, R.L.; Faulkner, P.; Downey, M.O.; Darby, S.J.; Falconer, R.J. The complexity of condensed tannin binding to bovine serum albumin—An isothermal titration calorimetry study. Food Chem. 2016, 190, 173–178. [Google Scholar] [CrossRef]
- Prigent, S.V.E.; Voragen, A.G.J.; van Koningsveld, G.A.; Baron, A.; Renard, C.M.G.C.; Gruppen, H. Interactions between globular proteins and procyanidins of different degrees of polymerization. J. Dairy Sci. 2009, 92, 5843–5853. [Google Scholar] [CrossRef] [Green Version]
- Monzani, E.; Gatti, A.L.; Profumo, A.; Casella, L.; Gullotti, M. Oxidation of phenolic compounds by lactoperoxidase. Evidence for the presence of a low-potential compound II during catalytic turnover. Biochemistry 1997, 36, 1918–1926. [Google Scholar] [CrossRef] [PubMed]
- Brück, T.B.; Harvey, P.J. Oxidation of mitoxantrone by lactoperoxidase. Biochim. Biophys. Acta 2003, 1649, 154–163. [Google Scholar] [CrossRef]
- Smullen, J.; Koutsou, G.A.; Foster, H.A.; Zumbé, A.; Storey, D.M. The antibacterial activity of plant extracts containing polyphenols against Streptococcus mutans. Caries Res. 2007, 41, 342–349. [Google Scholar] [CrossRef]
- Xiang, H.; Cao, F.; Ming, D.; Zheng, Y.; Dong, X.; Zhong, X.; Mu, D.; Li, B.; Zhong, L.; Cao, J.; et al. Aloe-emodin inhibits Staphylococcus aureus biofilms and extracellular protein production at the initial adhesion stage of biofilm development. Appl. Microbiol. Biotechnol. 2017, 101, 6671–6681. [Google Scholar] [CrossRef]
- Vaillancourt, K.; LeBel, G.; Pellerin, G.; Ben Lagha, A.; Grenier, D. Effects of the Licorice Isoflavans Licoricidin and Glabridin on the Growth, Adherence Properties, and Acid Production of Streptococcus mutans, and Assessment of Their Biocompatibility. Antibiotics 2021, 10, 163. [Google Scholar] [CrossRef] [PubMed]
- Selwitz, R.H.; Ismail, A.I.; Pitts, N.B. Dental caries. Lancet 2007, 369, 51–59. [Google Scholar] [CrossRef]
- Whelton, H.P.; Spencer, A.J.; Do, L.G.; Rugg-Gunn, A.J. Fluoride Revolution and Dental Caries: Evolution of Policies for Global Use. J. Dent. Res. 2019, 98, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Autio-Gold, J. The role of chlorhexidine in caries prevention. Oper. Dent. 2008, 33, 710–716. [Google Scholar] [CrossRef]
- Adams, S.E.; Arnold, D.; Murphy, B.; Carroll, P.; Green, A.K.; Smith, A.M.; Marsh, P.D.; Chen, T.; Marriott, R.E.; Brading, M.G. A randomised clinical study to determine the effect of a toothpaste containing enzymes and proteins on plaque oral microbiome ecology. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Söderling, E.M. Xylitol, mutans streptococci, and dental plaque. Adv. Dent. Res. 2009, 21, 74–78. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract or Fraction | Species | No | IC50 (µg/mL) | EC50 (µg/mL) | Activation (%) |
|---|---|---|---|---|---|
| Acetone | R. x bohemica | 1 | 3.203 ± 0.334 10,11,12,13 | - | - |
| R. japonica | 2 | 2.806 ± 0.253 10,11,12,13 | - | - | |
| R. sachalinensis | 3 | 2.657 ± 0.416 10,11,12 | - | - | |
| DCM | R. x bohemica | 4 | 2.525 ± 0.176 7,8,9,16,17,18 | 0.034 ± 0.001 7,8,9 | 18.71 |
| R. japonica | 5 | 3.266 ± 0.226 6,10,11 | - | - | |
| R. sachalinensis | 6 | 5.486 ± 0.759 7,8,9,17 | - | - | |
| Diethyl eter | R. x bohemica | 7 | 1.049 ± 0.044 4,6,9,10,11,13 | 0.009 ± 0.002 4 | 15.56 |
| R. japonica | 8 | 0.794 ± 0.067 4,6,9,10,11,12,13 | 0.006 ± 0.001 4 | 25.51 | |
| R. sachalinensis | 9 | 1.155 ± 0.059 4,6,7,8,10,11,12,13,14,15,18 | 0.008 ± 0.003 4 | 10.69 | |
| Ethyl acetate | R. x bohemica | 10 | 1.199 ± 0.100 1,2,3,5,7,8,9,16,17,18 | - | - |
| R. japonica | 11 | 1.771 ± 0.096 1,2,3,5,7,8,9,16,17,18 | - | - | |
| R. sachalinensis | 12 | 1.904 ± 0.240 8,9,16,17,18 | - | - | |
| Butanol | R. x bohemica | 13 | 2.663 ± 0.316 1,2,7,8,9,16,17,18 | - | - |
| R. japonica | 14 | 2.267 ± 0.315 9,17 | - | - | |
| R. sachalinensis | 15 | 1.056 ± 0.169 9,17 | - | - | |
| Water | R. x bohemica | 16 | 3.062 ± 0.674 4,10,11,12,13 | - | - |
| R. japonica | 17 | 10.039 ± 3.659 4,6,10,11,12,13,14,15 | - | - | |
| R. sachalinensis | 18 | 2.240 ± 0.316 4,9,10,11,12,13 | - | - | |
| Compound | IC50 (µM) | EC50 (µM) | Activation (%) | ||
| (-)-Epicatechin | a | 74.081 ± 3.624 b,d | 1.314 ± 0.239 b | 46.31 | |
| Resveratrol | b | 0.423 ± 0.018 a,d | 0.009 ± 0.002 a | 12.58 | |
| Emodin | c | - | - | - | |
| Vanicoside B | d | 1.000 ± 0.074 a,b | - | - |
| Species | Solvent | No | kobs1 (s−1) | kobs2 (s−1) | kobs1 (s−1) | kobs2 (s−1) |
|---|---|---|---|---|---|---|
| Compound I -> Compound II | Compound II -> Native LPO | |||||
| R. x bohemica | acetone | 1 | 18.71 ± 4.15 2,3,4,6 | - | 2.35 ± 0.30 2,3,5,6 | - |
| DCM | 2 | 87.36 ± 11.82 1,3,5,6 | 3.36 ± 0.50 | 18.72 ± 6.23 1,3,5,6 | 0.85 ± 0.18 3,4,6 | |
| Et2O | 3 | 485.60 ± 19.86 1,2,4,5,6 | - | 97.65 ± 11.14 1,2,4,5,6 | 6.24 ± 1.08 2,4,5,6 | |
| EtAc | 4 | 98.87 ± 20.85 1,3,5,6 | - | 14.57 ± 1.22 3,5,6 | 3.86 ± 0.88 2,3,5,6 | |
| butanol | 5 | 15.64 ± 2.80 2,3,4,6 | 2.62 ± 0.62 | 0.38 ± 0.07 1,2,3,4 | 0.10 ± 0.04 3,4 | |
| Water | 6 | 2.01 ± 0.22 1,2,3,4,5 | - | 0.23 ± 0.08 1,2,3,4 | 0.01 ± 0.01 2,3,4 | |
| R. japonica | acetone | 7 | 55.38 ± 2.31 8,11,12 | - | 5.81 ± 0.87 8,9,10,11,12 | - |
| DCM | 8 | 130.90 ± 26.22 7,11,12 | 2.63 ± 0.52 | 28.76 ± 6.71 7,9,10,11,12 | 3.43 ± 1.13 11,12 | |
| Et2O | 9 | 98.87 ± 20.85 11,12 | - | 14.57 ± 1.22 7,8,11,12 | 3.86 ± 0.88 11,12 | |
| EtAc | 10 | 79.04 ± 13.57 11,12 | - | 14.29 ± 0.61 7,8,11,12 | 4.20 ± 0.87 11,12 | |
| butanol | 11 | 20.03 ± 2.56 7,8,9,10,12 | 2.97 ± 0.54 | 0.66 ± 0.16 7,8,9,10 | 0.27 ± 0.108,9,10 | |
| Water | 12 | 2.11 ± 0.35 7,8,9,10,11 | - | 0.33 ± 0.05 7,8,9,10 | 0.02 ± 0.01 8,9,10 | |
| R. sachalinensis | acetone | 13 | 33.55 ± 8.40 15,17,18 | - | 0.84 ± 0.22 15,16 | - |
| DCM | 14 | 16.21 ± 2.21 15,16,18 | 2.11 ± 0.21 | 1.46 ± 1.17 15 | 0.41 ± 0.22 15,18 | |
| Et2O | 15 | 156.43 ± 16.57 13,14,16,17,18 | - | 22.38 ± 1.73 13,14,16,17,18 | 8.46 ± 1.7914,16,17,18 | |
| EtAc | 16 | 48.62 ± 5.06 14,15,17,18 | - | 4.18 ± 0.79 13,15,17,18 | 1.52 ± 0.31 15,18 | |
| butanol | 17 | 19.70 ± 3.90 13,15,16,18 | 3.37 ± 0.88 | 0.40 ± 0.07 15,16 | 0.11 ± 0.07 15 | |
| Water | 18 | 1.77 ± 0.21 13,14,15,16,17 | - | 0.42 ± 0.03 15,16 | 0.06 ± 0.01 14,15,16 | |
| Acetone Extract | k (ml µg−1 s−1) Compound I -> Compound II | k (ml µg−1 s−1) Compound II -> Native LPO |
| R. sachalinensis | 1.07 ± 0.10 | 0.07 ± 0.01 |
| R. japonica | 13.18 ± 0.31 | 1.38 ± 0.10 |
| R. x bohemica | 12.44 ± 0.55 | 0.49 ± 0.03 |
| Substrate | k (M−1 s−1) Compound I -> Compound II | k (M−1 s−1) Compound II -> Native LPO |
| (-)-Epicatechin | (4.3 ± 0.3) × 106 | (2.6 ± 0,4) × 106 |
| Resveratrol | N/D * | (1.71 ± 0.1) × 107 ** |
| Emodin | N/D *** | (2.5± 0.2) × 105 |
| Vanicoside B | (1.9 ± 0.3) × 106 | (3.6 ± 0.5) × 103 |
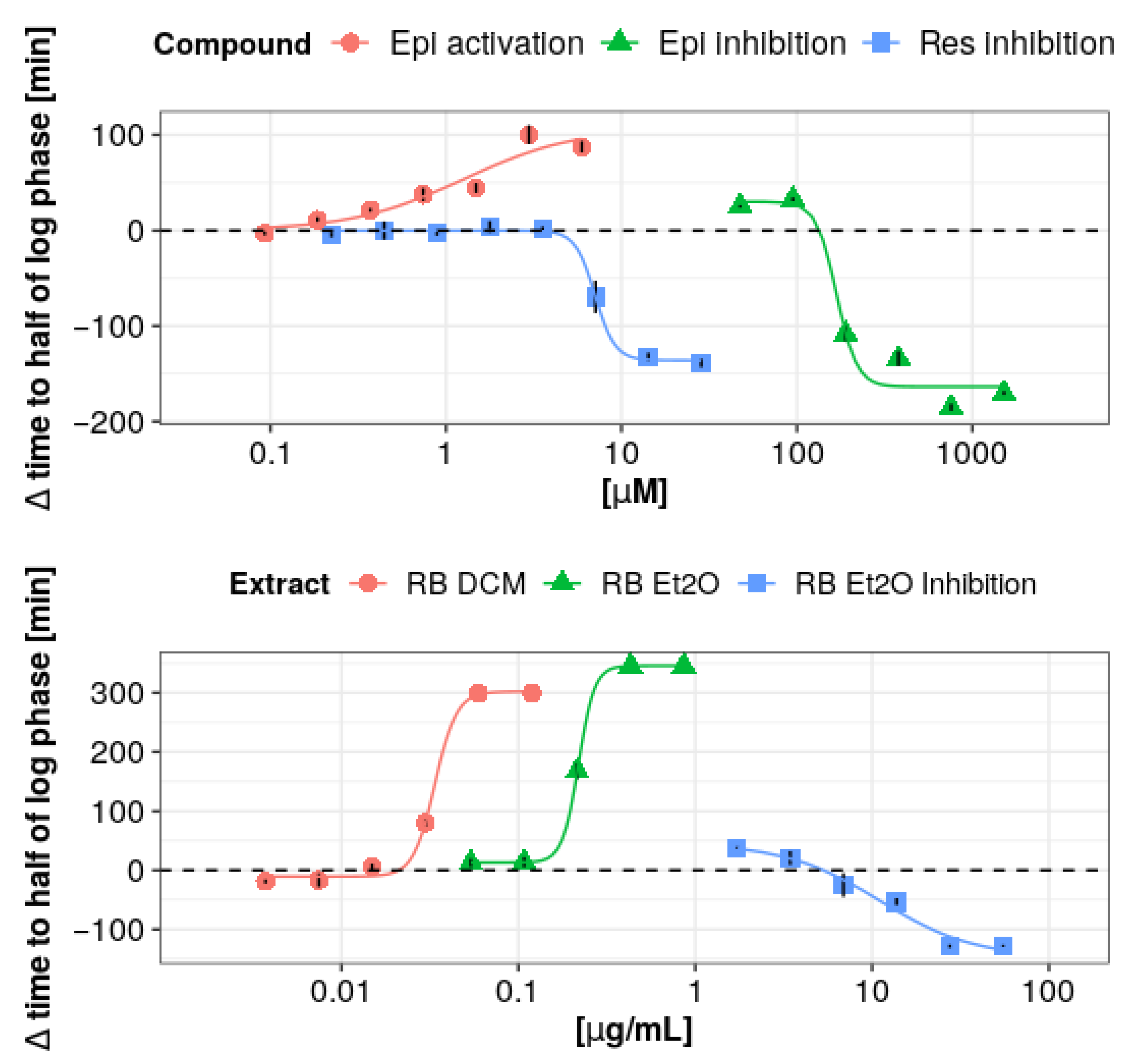
| Compound | Effect Type | ED10/IC10 (µM) | ED50/IC50 (µM) | ED90/IC90 (µM) | Δtmax (min) |
| (-)-Epicatechin | Activation | 0.13 ± 0.33 | 2.07 ± 1.01 | 14.57 ± 4.04 | 111 ± 11 |
| (-)-Epicatechin | Inhibition | 106.96 ± 9.58 | 161.106 ± 16.14 | 494.29 ± 168.37 | −185 ± 4 |
| Resveratrol | Inhibition | 5.17 ± 2.37 | 7.07 ± 0.27 | 9.67 ± 4.44 | −138 ± 4 |
| Extract | Effect Type | ED10/IC10 (µg/mL) | ED50/IC50 (µg/mL) | ED90/IC90 (µg/mL) | Δtmax (min) |
| R. x bohemica (DCM) | Activation | 0.02 ± 0.01 | 0.03 ± 0.01 | 0.05 ± 0.01 | 299 ± 7 |
| R. x bohemica (diethyl ether) | Activation | 0.17 ± 0.10 | 0.22 ± 0.01 | 0.28 ± 0.19 | 345 ± 13 |
| R. x bohemica (diethyl ether) | Inhibition | 12.40 ± 7.72 | 15.02 ± 7.42 | 18.17 ± 7.81 | −128 ± 3 |
| R. japonica (diethyl ether) | no effect | - | - | - | - |
| R. sachalinensis (diethyl ether) | no effect | - | - | - | - |
| Fraction | Species | MIC (mg/mL) | MBC (mg/mL) |
|---|---|---|---|
| Acetone | R. x bohemica | 0.30 | 0.60 |
| R. japonica | 0.15 | 0.60 | |
| R. sachalinensis | 0.30 | 1.20 | |
| DCM | R. x bohemica | 0.08 | >1.20 |
| R. japonica | 0.30 | >1.20 | |
| R. sachalinensis | 0.15 | >1.20 | |
| Etyl acetate | R. x bohemica | 0.30 | 0.60 |
| R. japonica | 0.15 | 0.60 | |
| R. sachalinensis | 0.30 | 0.60 | |
| Diethyl eter | R. x bohemica | 0.30 | >1.20 |
| R. japonica | 0.15 | >1.20 | |
| R. sachalinensis | 0.60 | >1.20 | |
| Butanol | R. x bohemica | 1.20 | >1.20 |
| R. japonica | 0.30 | 1.20 | |
| R. sachalinensis | 0.30 | 0.60 | |
| Water | R. x bohemica | >1.20 | >1.20 |
| R. japonica | 1.20 | >1.20 | |
| R. sachalinensis | >1.20 | >1.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magacz, M.; Oszajca, M.; Nawrot-Hadzik, I.; Drożdż, R.; Jurczak, A.; Hadzik, J.; Smakosz, A.; Krzyściak, W. Phenolic Compounds of Reynoutria sp. as Modulators of Oral Cavity Lactoperoxidase System. Antioxidants 2021, 10, 676. https://doi.org/10.3390/antiox10050676
Magacz M, Oszajca M, Nawrot-Hadzik I, Drożdż R, Jurczak A, Hadzik J, Smakosz A, Krzyściak W. Phenolic Compounds of Reynoutria sp. as Modulators of Oral Cavity Lactoperoxidase System. Antioxidants. 2021; 10(5):676. https://doi.org/10.3390/antiox10050676
Chicago/Turabian StyleMagacz, Marcin, Maria Oszajca, Izabela Nawrot-Hadzik, Ryszard Drożdż, Anna Jurczak, Jakub Hadzik, Aleksander Smakosz, and Wirginia Krzyściak. 2021. "Phenolic Compounds of Reynoutria sp. as Modulators of Oral Cavity Lactoperoxidase System" Antioxidants 10, no. 5: 676. https://doi.org/10.3390/antiox10050676
APA StyleMagacz, M., Oszajca, M., Nawrot-Hadzik, I., Drożdż, R., Jurczak, A., Hadzik, J., Smakosz, A., & Krzyściak, W. (2021). Phenolic Compounds of Reynoutria sp. as Modulators of Oral Cavity Lactoperoxidase System. Antioxidants, 10(5), 676. https://doi.org/10.3390/antiox10050676