
Mitigation of Aflatoxin B1 Hepatoxicity by Dietary Hedyotis diffusa Is Associated with Activation of NRF2/ARE Signaling in Chicks


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Birds, Dietary Treatments, and Sample Collections
2.2. Serum Biochemical and Histologic Analysis
2.3. Hepatic Antioxidant Parameters and AFBO–DNA Adduct Concentrations Analysis
2.4. Real-Time qPCR and Western Blot Analyses
2.5. Statistical Analysis
3. Results
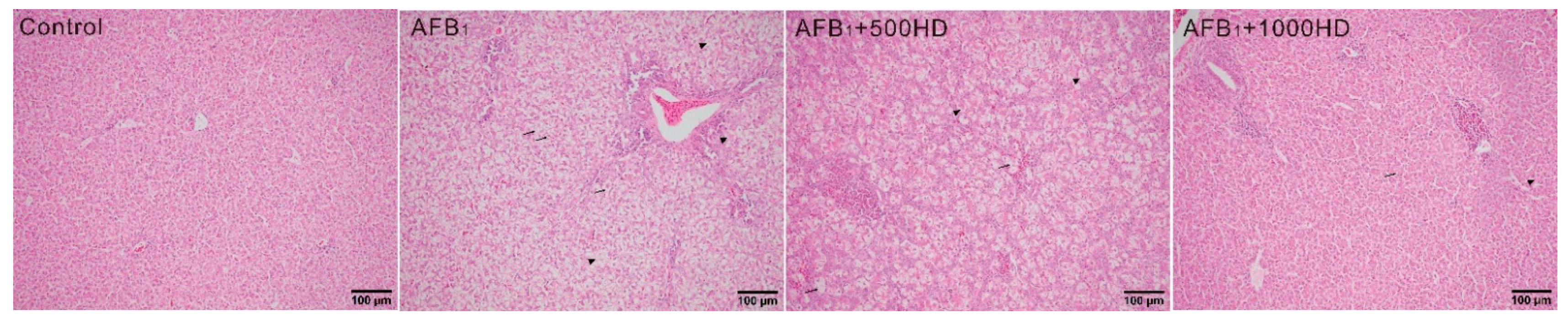
3.1. Growth Performance, Serum Biochemistry, and Liver Histology
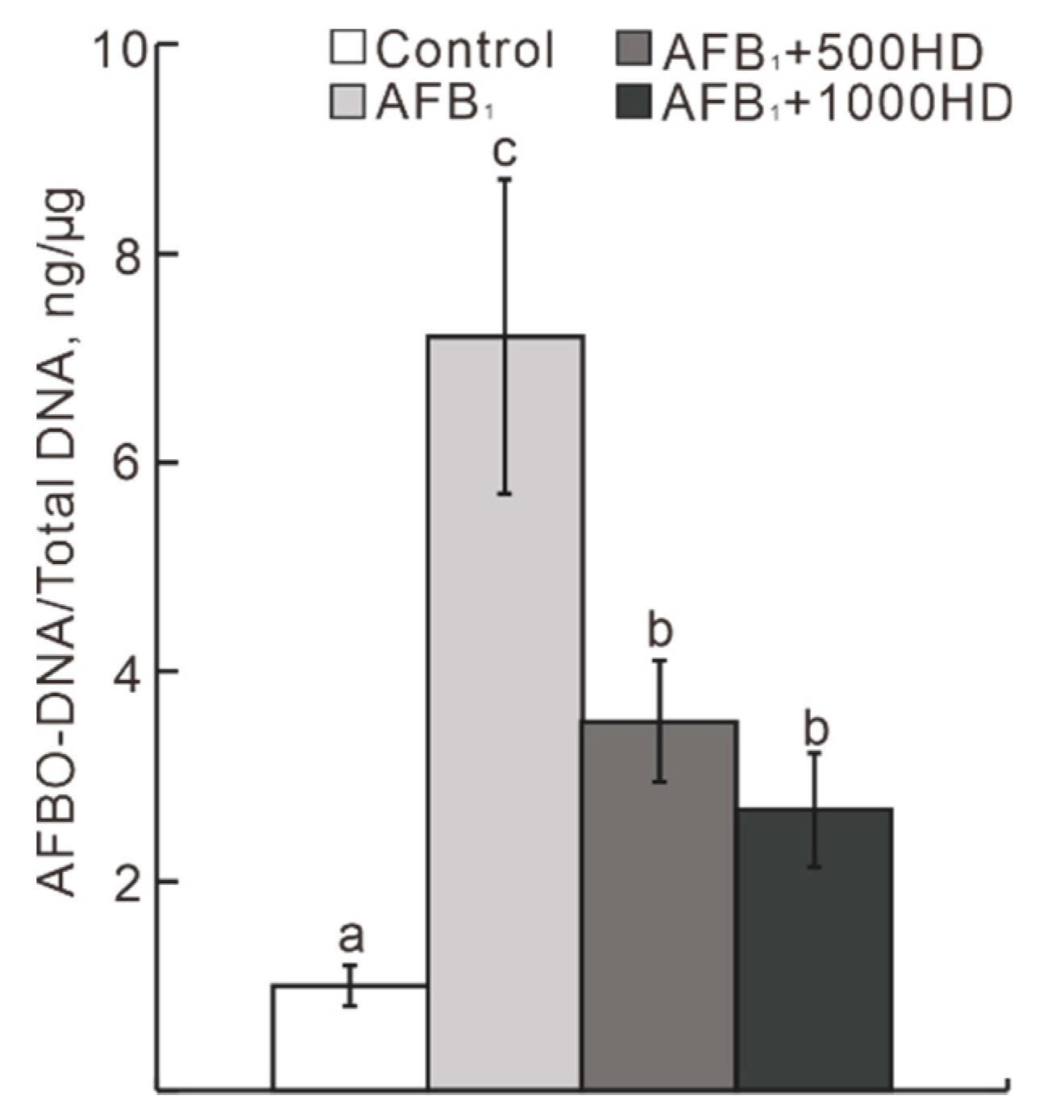
3.2. Hepatic Antioxidant Variables and AFBO–DNA Adduct Concentration
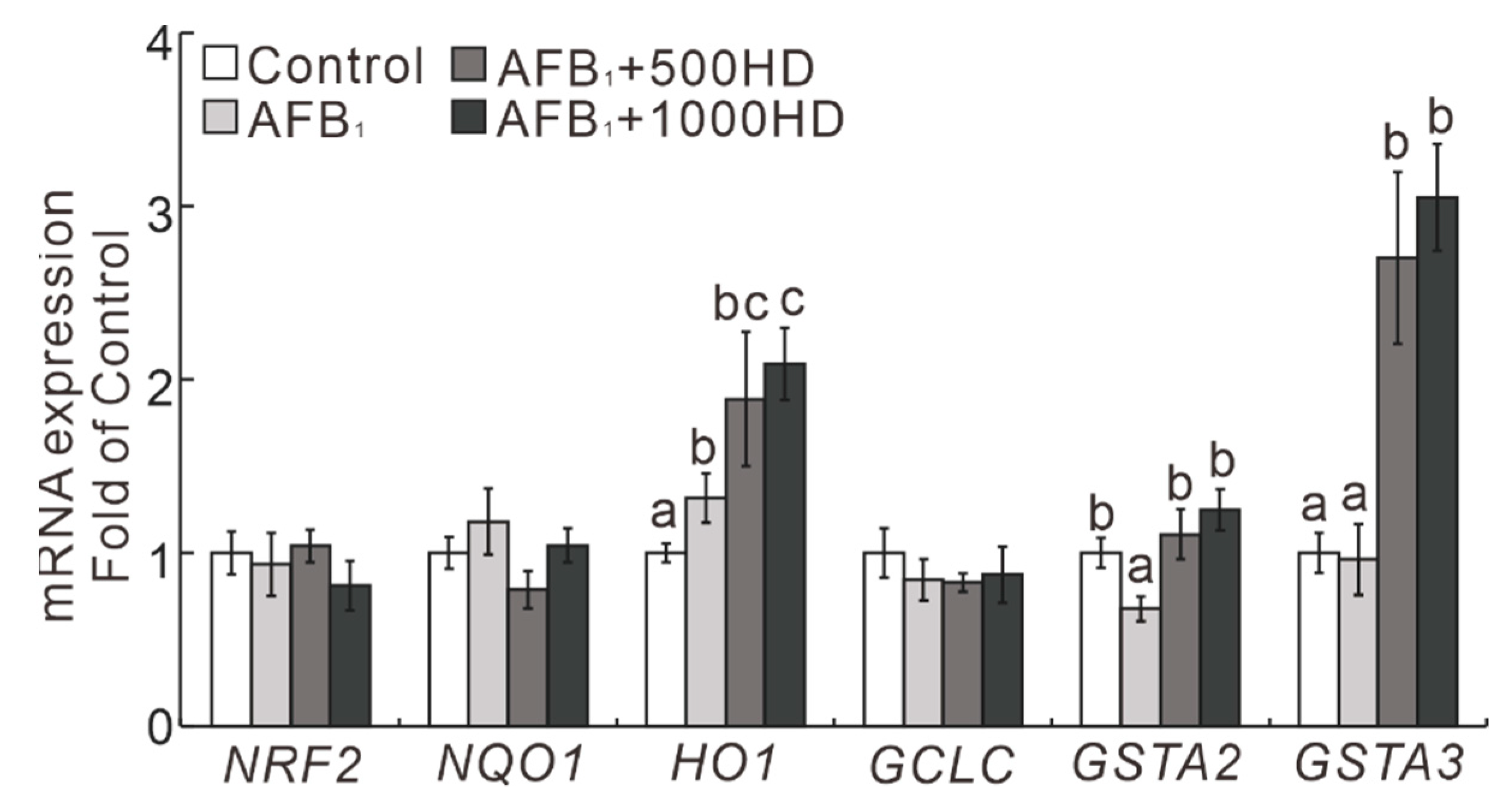
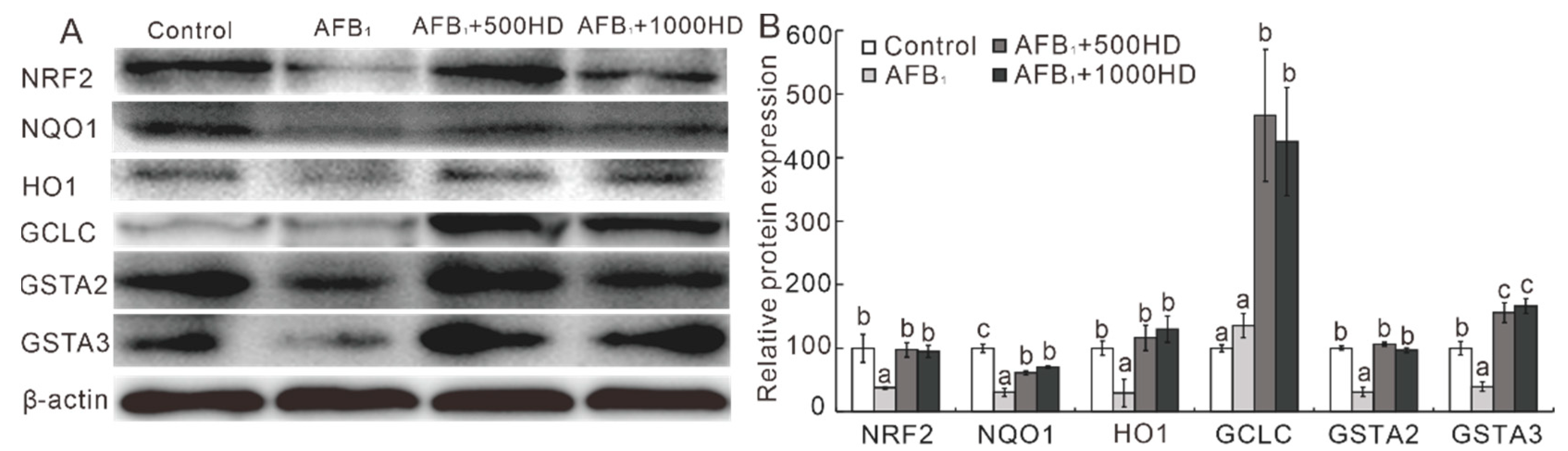
3.3. Expression of the NRF2/ARE Signaling-Related Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koehler, P.E.; Hanlin, R.T.; Beraha, L. Production of aflatoxins B1 and G1 by Aspergillus flavus and Aspergillus parasiticus isolated from market pecans. Appl. Microbiol. 1975, 30, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.H.; Lei, M.Y.; Zhang, N.Y.; Zhao, L.; Krumm, C.S.; Qi, D.S. Hepatotoxic effects of mycotoxin combinations in mice. Food Chem. Toxicol. 2014, 74, 289–293. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer (IARC). IARC Monographs on the Evaluation of Carcinogenic Risk of Chemicals to Humans; Overall Evaluation of Carcinogenicity: An Updating of IARC Monographs; IARC: Lyon, France, 1987. [Google Scholar]
- Guengerich, F.P.; Johnson, W.W.; Shimada, T.; Ueng, Y.F.; Yamazaki, H.; Langouët, S. Activation and detoxication of aflatoxin B1. Mutat. Res. 1998, 402, 121–128. [Google Scholar] [CrossRef]
- Yunus, A.W.; Razzazi-Fazeli, E.; Bohm, J. Aflatoxin B1 in affecting broiler’s performance, immunity, and gastrointestinal tract: A review of history and contemporary issues. Toxins 2011, 3, 566–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.H.; Lei, M.Y.; Zhang, N.Y.; Gao, X.; Li, C.; Krumm, C.S.; Qi, D.S. Individual and combined cytotoxic effects of aflatoxin B1, zearalenone, deoxynivalenol and fumonisin B1 on BRL 3A rat liver cells. Toxicon 2015, 95, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Hussein, H.S.; Brasel, J.M. Toxicity, metabolism, and impact of mycotoxins on humans and animals. Toxicology 2001, 167, 101–134. [Google Scholar] [CrossRef]
- Deng, J.; Zhao, L.; Zhang, N.Y.; Karrow, N.A.; Krumm, C.S.; Qi, D.S.; Sun, L.H. Aflatoxin B1 metabolism: Regulation by phase I and II metabolizing enzymes and chemoprotective agents. Mutat. Res. 2018, 778, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Mary, V.S.; Theumer, M.G.; Arias, S.L.; Rubinstein, H.R. Reactive oxygen species sources and biomolecular oxidative damage induced by aflatoxin B1 and fumonisin B1 in rat spleen mononuclear cells. Toxicology 2012, 302, 299–307. [Google Scholar] [CrossRef]
- Sun, L.H.; Zhang, N.Y.; Zhu, M.K.; Zhao, L.; Zhou, J.C.; Qi, D.S. Prevention of Aflatoxin B1 Hepatoxicity by Dietary Selenium Is Associated with Inhibition of Cytochrome P450 Isozymes and Up-Regulation of 6 Selenoprotein Genes in Chick Liver. J. Nutr. 2016, 146, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Abidin, Z.; Khatoon, A.; Numan, M. Mycotoxins in broilers: Pathological alterations induced by aflatoxins and ochratoxins, diagnosis and determination, treatment and control of mycotoxicosis. Worlds Poult. Sci. J. 2011, 67, 485–496. [Google Scholar] [CrossRef]
- Manafi, M.; Heydari, M.; Yari, M. Aflatoxicosis and herbal detoxification: The effectiveness of thyme essence on performance parameters and antibody titers of commercial broilers fed aflatoxin B1. Res. Zool. 2014, 4, 43–50. [Google Scholar]
- Nazarizadeh, H.; Hosseini, S.M.; Pourreza, J. Effect of plant extracts derived from thyme and chamomile on the growth performance, gut morphology and immune system of broilers fed aflatoxin B1 and ochratoxin A contaminated diets. Ital. J. Anim. Sci. 2019, 18, 1073–1081. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Zhou, X.; Wang, Y.; Wei, D.; Deng, C.; Xu, X.; Xin, P.; Sun, S. The Antitumor Constituents from Hedyotis Diffusa Willd. Molecules 2017, 22, 2101. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Zhang, D.; Yi, J.; Shao, J. Anticancer activities of Oldenlandia diffusa. J. Herb. Pharmacother. 2004, 4, 21–33. [Google Scholar] [CrossRef]
- Ahmad, R.; Ali, A.M.; Israf, D.A.; Ismail, N.H.; Shaari, K.; Lajis, N.H. Antioxidant, radical-scavenging, anti-inflammatory, cytotoxic and antibacterial activities of methanolic extracts of some Hedyotis species. Life Sci. 2005, 76, 1953–1964. [Google Scholar] [CrossRef]
- Lin, C.C.; Ng, L.T.; Yang, J.J.; Hsu, Y.F. Anti-inflammatory and hepatoprotective activity of peh-hue-juwa-chi-cao in male rats. Am. J. Chin. Med. 2002, 30, 225–234. [Google Scholar] [CrossRef]
- Ren, F.; Liu, G.; Zhang, L.; Niu, G. Studies on chemical constituents of Hedyotis diffusa Willd. Chin. Pharm. J. 2005, 40, 502–504. [Google Scholar]
- Ahmad, R.; Shaari, K.; Lajis, N.H.; Hamzah, A.S.; Ismail, N.H.; Kitajima, M. Anthraquinones from Hedyotis capitellata. Phytochemistry 2005, 66, 1141–1147. [Google Scholar] [CrossRef] [PubMed]
- Wong, B.; Lau, B.; Yamasaki, T.; Teel, R. Inhibition of dexamethasone-induced cytochrome P450-mediated mutagenicity and metabolism of aflatoxin B1 by Chinese medicinal herbs. Eur. J. Cancer Prev. 1993, 2, 351–356. [Google Scholar] [CrossRef]
- Wong, B.Y.; Lau, B.H.; Tadi, P.P.; Teel, R.W. Chinese medicinal herbs modulate mutagenesis, DNA binding and metabolism of aflatoxin B1. Mutat. Res. 1992, 279, 209–216. [Google Scholar] [CrossRef]
- Yarmohammadi, F.; Rezaee, R.; Karimi, G. Natural compounds against doxorubicin-induced cardiotoxicity: A review on the involvement of Nrf2/ARE signaling pathway. Phytother. Res. 2021, 35, 1163–1175. [Google Scholar] [CrossRef]
- Guo, Y.; Yu, S.; Zhang, C.; Kong, A.T. Epigenetic regulation of Keap1-Nrf2 signaling. Free Radic. Biol. Med. 2015, 88, 337–349. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.M.; Johnson, J.A. An important role of Nrf2-ARE pathway in the cellular defense mechanism. J. Biochem. Mol. Biol. 2004, 37, 139–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, P.; Chattopadhyay, A. Nrf2-ARE signaling in cellular protection: Mechanism of action and the regulatory mechanisms. J. Cell Physiol. 2020, 235, 3119–3130. [Google Scholar] [CrossRef] [PubMed]
- Gu, G.W.; Barone, I.; Gelsomino, L.; Giordano, C.; Bonofiglio, D.; Statti, G.; Menichini, F.; Catalano, S.; Andò, S. Oldenlandia diffusa extracts exert antiproliferative and apoptotic effects on human breast cancer cells through ERα/Sp1-mediated p53 activation. J. Cell Physiol. 2012, 227, 3363–3372. [Google Scholar] [CrossRef]
- Zhao, L.; Deng, J.; Ma, L.B.; Zhang, W.P.; Khalil, M.M.; Karrow, N.A.; Qi, D.S.; Sun, L.H. Dietary Se deficiency dysregulates metabolic and cell death signaling in aggravating the AFB1 hepatotoxicity of chicks. Food Chem. Toxicol. 2021, 149, 111938. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Sun, L.H.; Huang, J.Q.; Briens, M.; Qi, D.S.; Xu, S.W.; Lei, X.G. A novel organic selenium compound exerts unique regulation of selenium speciation, selenogenome, and selenoproteins in broiler chicks. J. Nutr. 2017, 147, 789–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.H.; Zhang, N.Y.; Sun, R.R.; Gao, X.; Gu, C.; Krumm, C.S.; Qi, D.S. A novel strain of Cellulosimicrobium funkei can biologically detoxify aflatoxin B1 in ducklings. Microb. Biotechnol. 2015, 8, 490–498. [Google Scholar] [CrossRef]
- Huang, J.Q.; Ren, F.Z.; Jiang, Y.Y.; Xiao, C.; Lei, X.G. Selenoproteins protect against avian nutritional muscular dystrophy by metabolizing peroxides and regulating redox/apoptotic signaling. Free Radic. Biol. Med. 2015, 83, 129–138. [Google Scholar] [CrossRef]
- Liu, J.; Song, W.J.; Zhang, N.Y.; Tan, J.; Krumm, C.S.; Sun, L.H.; Qi, D.S. Biodetoxification of aflatoxin B1 in cottonseed meal by fermentation of Cellulosimicrobium funkei in duckling diet. Poult. Sci. 2017, 96, 923–930. [Google Scholar] [CrossRef]
- Zhao, L.; Feng, Y.; Deng, J.; Zhang, N.Y.; Zhang, W.P.; Liu, X.L.; Rajput, S.A.; Qi, D.S.; Sun, L.H. Selenium Deficiency Aggravates Aflatoxin B1-Induced Immunotoxicity in Chick Spleen by Regulating 6 Selenoprotein Genes and Redox/Inflammation/Apoptotic Signaling. J. Nutr. 2019, 149, 894–901. [Google Scholar] [CrossRef] [PubMed]
- Kasmani, F.B.; Torshizi, M.A.K.; Allameh, A.; Shariatmadari, F. A novel aflatoxin-binding Bacillus probiotic: Performance, serum biochemistry, and immunological parameters in Japanese quail. Poult. Sci. 2012, 91, 1846–1853. [Google Scholar] [CrossRef]
- Khaleghipour, B.; Khosravinia, H.; Toghiyani, M.; Azarfar, A. Effects of silymarin on productive performance, liver function and serum biochemical profile in broiler Japanese quail challenged with dietary aflatoxins. Ital. J. Anim. Sci. 2019, 18, 564–573. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; He, J.; Tong, X.; Tang, L.; Liu, M. The Hedyotis diffusa Willd. (Rubiaceae): A Review on Phytochemistry, Pharmacology, Quality Control and Pharmacokinetics. Molecules 2016, 21, 710. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Li, C.; Tang, Y.L.; Zhang, H.; Chan, S.W. Effect of Hedyotis diffusa water extract on protecting human hepatocyte cells (LO2) from H2O2-induced cytotoxicity. Pharm. Biol. 2016, 54, 1148–1155. [Google Scholar] [PubMed] [Green Version]
- Dinkova-Kostova, A.T.; Talalay, P. NAD(P)H:quinone acceptor oxidoreductase 1 (NQO1), a multifunctional antioxidant enzyme and exceptionally versatile cytoprotector. Arch. Biochem. Biophys. 2010, 501, 116–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell. Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Jin, Y.; Yu, H.; Shan, A.; Shen, J.; Zhou, C.; Zhao, Y.; Fang, H.; Wang, X.; Wang, J.; et al. Resveratrol inhibits aflatoxin B1-induced oxidative stress and apoptosis in bovine mammary epithelial cells and is involved the Nrf2 signaling pathway. Toxicon 2019, 164, 10–15. [Google Scholar] [CrossRef]
- Ji, Y.; Nyamagoud, S.B.; SreeHarsha, N.; Mishra, A.; Gubbiyappa, S.K.; Singh, Y. Sitagliptin protects liver against aflatoxin B1-induced hepatotoxicity through upregulating Nrf2/ARE/HO-1 pathway. Biofactors 2020, 46, 76–82. [Google Scholar] [CrossRef]
- Lu, S.C. Glutathione synthesis. Biochim. Biophys. Acta. 2013, 1830, 3143–3153. [Google Scholar] [CrossRef] [Green Version]
- Sriwattanapong, K.; Slocum, S.L.; Chawanthayatham, S.; Fedeles, B.I.; Egner, P.A.; Groopman, J.D.; Satayavivad, J.; Croy, R.G.; Essigmann, J.M. Editor’s Highlight: Pregnancy Alters Aflatoxin B1 Metabolism and Increases DNA Damage in Mouse Liver. Toxicol. Sci. 2017, 160, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Balasubramanian, B.; Zhao, Z.H.; Liu, W.C. Marine algal polysaccharides alleviate aflatoxin B1-induced bursa of Fabricius injury by regulating redox and apoptotic signaling pathway in broilers. Poult. Sci. 2021, 100, 844–857. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.Y.; Qi, M.; Gao, X.; Zhao, L.; Liu, J.; Gu, C.Q.; Song, W.J.; Krumm, C.S.; Sun, L.H.; Qi, D.S. Response of the hepatic transcriptome to aflatoxin B1 in ducklings. Toxicon 2016, 111, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.Y.; Qi, M.; Zhao, L.; Zhu, M.K.; Guo, J.; Liu, J.; Gu, C.Q.; Rajput, S.A.; Krumm, C.S.; Qi, D.S.; et al. Curcumin Prevents Aflatoxin B1 Hepatoxicity by Inhibition of Cytochrome P450 Isozymes in Chick Liver. Toxins 2016, 8, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | AFB1 | AFB1 + 500HD | AFB1 + 1000HD | |
|---|---|---|---|---|
| Initial Body Weight, g | 69.4 ± 0.1 | 69.2 ± 0.1 | 69.3 ± 0.1 | 69.4 ± 0.2 |
| Final Body Weight, g | 647± 8.3 c | 575 ± 12.5 a | 592 ± 17.4 ab | 619 ± 8.8 b |
| Body Weight Gain, g | 577 ± 8.3 c | 506 ± 12.5 a | 523 ± 17.4 ab | 550 ± 8.8 b |
| Feed Intake, g | 676 ± 9.8 b | 632 ± 12.9 a | 636± 16.7 ab | 662± 8.8 ab |
| Gain/Feed, g/kg | 854 ± 9 b | 801 ± 24 a | 822 ± 17 ab | 831 ± 5 a |
| Control | AFB1 | AFB1 + 500HD | AFB1 + 1000HD | |
|---|---|---|---|---|
| ALT, U/L | 3.40 ± 0.43 | 2.73 ± 0.55 | 3.79 ± 0.56 | 3.23 ± 0.72 |
| AST, U/L | 268 ± 18.7 | 270 ± 27.4 | 230 ± 5.8 | 243 ± 21.7 |
| TBIL, µmol/L | 3.64 ± 0.06 | 3.65 ±0.08 | 3.70 ± 0.15 | 3.75 ±0.2 |
| ALB, g/L | 16.3 ± 0.5 b | 14.1 ± 0.9 a | 17.6 ± 1.7 ab | 16.6 ± 0.5 b |
| TP, g/L | 26.1 ± 1.2 b | 20.7 ± 3.0 a | 23.8 ± 2.0 ab | 37.5 ± 5.0 c |
| Control | AFB1 | AFB1 + 500HD | AFB1 + 1000HD | |
|---|---|---|---|---|
| SOD, U/mg Protein | 88.7 ± 12.1 ab | 83.7 ± 7.1 a | 94.5 ± 12.4 ab | 103 ±17.5 b |
| GPX, U/mg Protein | 26.2 ± 4.8 b | 21.0 ± 2.1 a | 24.0 ± 5.3 ab | 30.6 ± 7.8 b |
| CAT, U/mg Protein | 28.4 ± 5.1 ab | 25.9 ± 3.0 a | 27.5 ± 5.4 ab | 36.5 ± 9.9 b |
| MDA, nmol/mg Protein | 1.15 ± 0.26 | 1.01 ± 0.24 | 1.18 ± 0.22 | 1.25 ± 0.32 |
| PC, nmol/mg Protein | 6.19 ± 1.04 a | 8.26 ± 2.09 b | 6.08 ± 0.92 a | 5.58 ± 0.18 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, L.; Deng, J.; Xu, Z.-J.; Zhang, W.-P.; Khalil, M.M.; Karrow, N.A.; Sun, L.-H. Mitigation of Aflatoxin B1 Hepatoxicity by Dietary Hedyotis diffusa Is Associated with Activation of NRF2/ARE Signaling in Chicks. Antioxidants 2021, 10, 878. https://doi.org/10.3390/antiox10060878
Zhao L, Deng J, Xu Z-J, Zhang W-P, Khalil MM, Karrow NA, Sun L-H. Mitigation of Aflatoxin B1 Hepatoxicity by Dietary Hedyotis diffusa Is Associated with Activation of NRF2/ARE Signaling in Chicks. Antioxidants. 2021; 10(6):878. https://doi.org/10.3390/antiox10060878
Chicago/Turabian StyleZhao, Ling, Jiang Deng, Zi-Jian Xu, Wan-Po Zhang, Mahmoud Mohamed Khalil, Niel Alexander Karrow, and Lv-Hui Sun. 2021. "Mitigation of Aflatoxin B1 Hepatoxicity by Dietary Hedyotis diffusa Is Associated with Activation of NRF2/ARE Signaling in Chicks" Antioxidants 10, no. 6: 878. https://doi.org/10.3390/antiox10060878
APA StyleZhao, L., Deng, J., Xu, Z. -J., Zhang, W. -P., Khalil, M. M., Karrow, N. A., & Sun, L. -H. (2021). Mitigation of Aflatoxin B1 Hepatoxicity by Dietary Hedyotis diffusa Is Associated with Activation of NRF2/ARE Signaling in Chicks. Antioxidants, 10(6), 878. https://doi.org/10.3390/antiox10060878