
Redox Regulation in Diazotrophic Bacteria in Interaction with Plants

 , ,
, ,
Abstract
:1. Introduction
2. The Bacterial Response to ROS
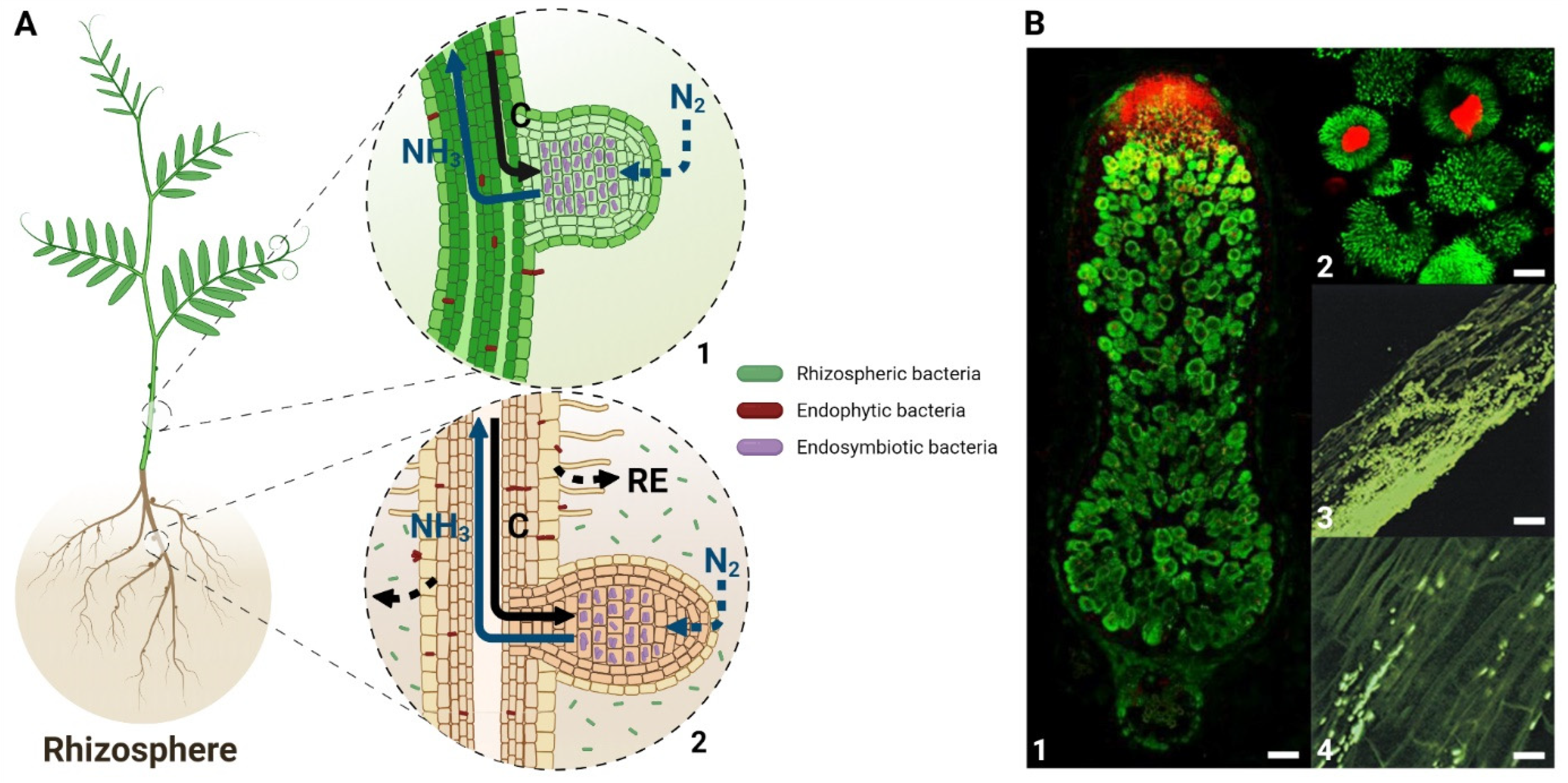
3. Redox Regulation in Rhizosphere and Root Colonization
4. Antioxidant Defense in Legume Symbionts
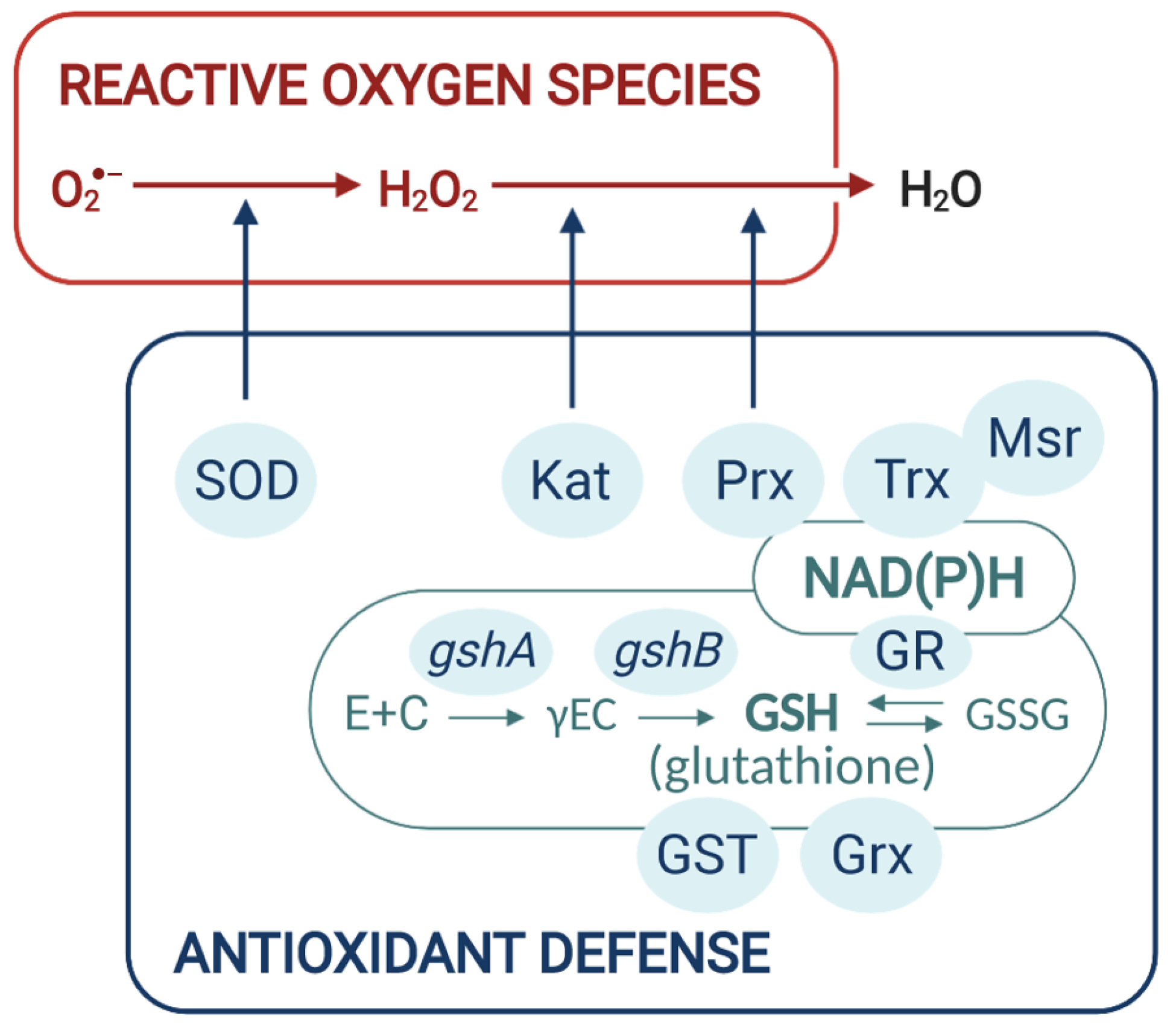
4.1. ROS Scavenging Enzymes
4.2. The Role of Bacterial GSH/Grx System
5. Redox-Based Transcriptional Regulation in Endophytic and Endosymbiotic Bacteria
6. ROS Transcriptional and Post-Translational Control of Nodule Metabolism during Nitrogen Fixation
7. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Jiao, X.; Takishita, Y.; Zhou, G.; Smith, D.L. Plant Associated Rhizobacteria for Biocontrol and Plant Growth Enhancement. Front. Plant Sci. 2021, 12, 420. [Google Scholar] [CrossRef]
- Robertson, G.P.; Vitousek, P.M. Nitrogen in agriculture: Balancing the cost of an essential resource. Annu. Rev. Environ. Resour. 2009, 34, 97–125. [Google Scholar] [CrossRef] [Green Version]
- Soumare, A.; Diedhiou, A.G.; Thuita, M.; Hafidi, M.; Ouhdouch, Y.; Gopalakrishnan, S.; Kouisni, L. Exploiting Biological Nitrogen Fixation: A Route Towards a Sustainable Agriculture. Plants 2020, 9, 1011. [Google Scholar] [CrossRef] [PubMed]
- Welte, C.U. Revival of Archaeal Methane Microbiology. mSystems 2018, 3, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, T.L.G.; Balsemão-Pires, E.; Saraiva, R.M.; Ferreira, P.C.G.; Hemerly, A.S. Nitrogen signalling in plant interactions with associative and endophytic diazotrophic bacteria. J. Exp. Bot. 2014, 65, 5631–5642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, U.; Johansson, C.; Bergman, B. Early communication in the Gunnera-Nostoc symbiosis: Plant-induced cell differentiation and protein synthesis in the cyanobacterium. Mol. Plant Microbe Interact. 1994, 7, 696–702. [Google Scholar] [CrossRef]
- Pawlowski, K.; Demchenko, K.N. The diversity of actinorhizal symbiosis. Protoplasma 2012, 249, 967–979. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Liu, W.; Nandety, R.S.; Crook, A.; Mysore, K.S.; Pislariu, C.I.; Frugoli, J.; Dickstein, R.; Udvardi, M.K. Celebrating 20 Years of Genetic Discoveries in Legume Nodulation and Symbiotic Nitrogen Fixation. Plant Cell 2020, 32, 15–41. [Google Scholar] [CrossRef] [Green Version]
- Chelius, M.K.; Triplett, E.W. Immunolocalization of dinitrogenase reductase produced by Kiebsiella pneumoniae in association with Zea mays L. Appl. Environ. Microbiol. 2000, 66, 783–787. [Google Scholar] [CrossRef] [Green Version]
- Frederickson Matika, D.E.; Loake, G.J. Redox Regulation in Plant Immune Function. Antioxid. Redox Signal. 2014, 21, 1373–1388. [Google Scholar] [CrossRef]
- Syska, C.; Brouquisse, R.; Alloing, G.; Pauly, N.; Frendo, P.; Bosseno, M.; Dupont, L.; Boscari, A. Molecular Weapons Contribute to Intracellular Rhizobia Accommodation Within Legume Host Cell. Front. Plant Sci. 2019, 10, 1496. [Google Scholar] [CrossRef]
- Zamioudis, C.; Pieterse, C.M.J. Modulation of Host Immunity by Beneficial Microbes. Mol. Plant Microbe Interact. 2012, 25, 139–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skelly, M.J.; Loake, G.J. Synthesis of redox-active molecules and their signaling functions during the expression of plant disease resistance. Antioxid. Redox Signal. 2013, 19, 990–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zipfel, C.; Oldroyd, G.E.D. Plant signalling in symbiosis and immunity. Nature 2017, 543, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Alberton, D.; Valdameri, G.; Moure, V.R.; Monteiro, R.A.; Pedrosa, F.O.; Müller-Santos, M.; de Souza, E.M. What Did We Learn from Plant Growth-Promoting Rhizobacteria (PGPR)-Grass Associations Studies through Proteomic and Metabolomic Approaches? Front. Sustain. Food Syst. 2020, 4, e607343. [Google Scholar] [CrossRef]
- Malviya, M.K.; Li, C.-N.; Solanki, M.K.; Singh, R.K.; Htun, R.; Singh, P.; Verma, K.K.; Yang, L.-T.; Li, Y.-R. Comparative analysis of sugarcane root transcriptome in response to the plant growth-promoting Burkholderia anthina MYSP113. PLoS ONE 2020, 15, e0231206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spaepen, S.; Bossuyt, S.; Engelen, K.; Marchal, K.; Vanderleyden, J. Phenotypical and molecular responses of Arabidopsis thaliana roots as a result of inoculation with the auxin-producing bacterium Azospirillum brasilense. New Phytol. 2014, 201, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Lade, S.B.; Román, C.; Cueto-Ginzo, A.I.; Serrano, L.; Sin, E.; Achón, M.A.; Medina, V. Host-specific proteomic and growth analysis of maize and tomato seedlings inoculated with Azospirillum brasilense Sp7. Plant Physiol. Biochem. 2018, 129, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.W.; Park, J.Y.; Lee, S.H.; Hwang, C.H. Comparative proteomic analysis of Soybean nodulation using a supernodulation mutant, SS2-2. Biosci. Biotechnol. Biochem. 2010, 74, 2396–2404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukami, J.; Ollero, F.J.; Megías, M.; Hungria, M. Phytohormones and induction of plant-stress tolerance and defense genes by seed and foliar inoculation with Azospirillum brasilense cells and metabolites promote maize growth. AMB Express 2017, 7, 153. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Gómez, M.; Castro-Mercado, E.; Alexandre, G.; García-Pineda, E. Oxidative and antioxidative responses in the wheat-Azospirillum brasilense interaction. Protoplasma 2016, 253, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Damiani, I.; Puppo, A.; Frendo, P. Redox Changes during the Legume–Rhizobium Symbiosis. Mol. Plant 2009, 2, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Nanda, A.K.; Andrio, E.; Marino, D.; Pauly, N.; Dunand, C. Reactive Oxygen Species during Plant-microorganism Early Interactions. J. Integr. Plant Biol. 2010, 52, 195–204. [Google Scholar] [CrossRef]
- Cárdenas, L.; Martínez, A.; Sánchez, F.; Quinto, C. Fast, transient and specific intracellular ROS changes in living root hair cells responding to Nod factors (NFs). Plant J. 2008, 56, 802–813. [Google Scholar] [CrossRef]
- Fournier, J.; Timmers, A.C.J.; Sieberer, B.J.; Jauneau, A.; Chabaud, M.; Barker, D.G. Mechanism of infection thread elongation in root hairs of Medicago truncatula and dynamic interplay with associated rhizobial colonization. Plant Physiol. 2008, 148, 1985–1995. [Google Scholar] [CrossRef] [Green Version]
- Peleg-Grossman, S.; Volpin, H.; Levine, A. Root hair curling and Rhizobium infection in Medicago truncatula are mediated by phosphatidylinositide-regulated endocytosis and reactive oxygen species. J. Exp. Bot. 2007, 58, 1637–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arthikala, M.K.; Montiel, J.; Sánchez-López, R.; Nava, N.; Cárdenas, L.; Quinto, C. Respiratory burst oxidase homolog gene a is crucial for Rhizobium infection and nodule maturation and function in common bean. Front. Plant Sci. 2017, 8, 2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamet, A.; Mandon, K.; Puppo, A.; Hérouart, D.; Hérouart, D. H2O2 is required for optimal establishment of the Medicago sativa/Sinorhizobium meliloti symbiosis. J. Bacteriol. 2007, 189, 8741–8745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gucciardo, S.; Rathbun, E.A.; Shanks, M.; Jenkyns, S.; Mak, L.; Durrant, M.C.; Brewin, N.J. Epitope Tagging of Legume Root Nodule Extensin Modifies Protein Structure and Crosslinking in Cell Walls of Transformed Tobacco Leaves. Mol. Plant Microbe Interact. 2005, 18, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.; Bhattacharyya, S.; Kumar, R.; Kumar, A.; Ibañez, F.; Wang, J.; Guo, B.; Sudini, H.K.; Gopalakrishnan, S.; DasGupta, M.; et al. Molecular Basis of Root Nodule Symbiosis between Bradyrhizobium and ‘Crack-Entry’ Legume Groundnut (Arachis hypogaea L.). Plants 2020, 9, 276. [Google Scholar] [CrossRef] [Green Version]
- D’Haeze, W.; de Rycke, R.; Mathis, R.; Goormachtig, S.; Pagnotta, S.; Verplancke, C.; Capoen, W.; Holsters, M. Reactive oxygen species and ethylene play a positive role in lateral root base nodulation of a semiaquatic legume. Proc. Natl. Acad. Sci. USA 2003, 100, 11789–11794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrio, E.; Marino, D.; Marmeys, A.; de Segonzac, M.D.; Damiani, I.; Genre, A.; Huguet, S.; Frendo, P.; Puppo, A.; Pauly, N. Hydrogen peroxide-regulated genes in the Medicago truncatula-Sinorhizobium meliloti symbiosis. New Phytol. 2013, 198, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Haag, A.F.; Mergaert, P. Terminal bacteroid differentiation in the Medicago–Rhizobium interaction—A tug of war between plant and bacteria. In The Model Legume Medicago Truncatula; de Bruijn, F., Ed.; Wiley: New York, NY, USA, 2020; pp. 600–616. [Google Scholar]
- Haag, A.F.; Kerscher, B.; Dall’Angelo, S.; Sani, M.; Longhi, R.; Baloban, M.; Wilson, H.M.; Mergaert, P.; Zanda, M.; Ferguson, G.P. Role of cysteine residues and disulfide bonds in the activity of a legume root nodule-specific, cysteine-rich peptide. J. Biol. Chem. 2012, 287, 10791–10798. [Google Scholar] [CrossRef] [Green Version]
- Shabab, M.; Arnold, M.F.F.; Penterman, J.; Wommack, A.J.; Bocker, H.T.; Price, P.A.; Griffitts, J.S.; Nolan, E.M.; Walker, G.C. Disulfide cross-linking influences symbiotic activities of nodule peptide NCR247. Proc. Natl. Acad. Sci. USA 2016, 113, 10157–10162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, C.W.; Baldacci-Cresp, F.; Pierre, O.; Larousse, M.; Benyamina, S.; Lambert, A.; Hopkins, J.; Castella, C.; Cazareth, J.; Alloing, G.; et al. Regulation of Differentiation of Nitrogen-Fixing Bacteria by Microsymbiont Targeting of Plant Thioredoxin s1. Curr. Biol. 2017, 27, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Becana, M.; Ibaliz Larrainzar, E.; Villar, I.; Rubio, M.C.; Erez-Rontom, E.C.P.; Huertas, R.; Sato, S.; Mun, J.-H. Research review Hemoglobins in the legume-Rhizobium symbiosis. New Phytol. 2020, 228, 472–484. [Google Scholar] [CrossRef]
- Marino, D.; Andrio, E.; Danchin, E.G.J.; Oger, E.; Gucciardo, S.; Lambert, A.; Puppo, A.; Pauly, N. A Medicago truncatula NADPH oxidase is involved in symbiotic nodule functioning. New Phytol. 2011, 189, 580–592. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhao, Y.; Feng, X.; Luo, Z.; Kong, S.; Zhang, C.; Gong, A.; Yuan, H.; Cheng, L.; Wang, X. Genomic, molecular evolution, and expression analysis of NOX genes in soybean (Glycine max). Genomics 2019, 111, 619–628. [Google Scholar] [CrossRef]
- Frendo, P.; Matamoros, M.A.; Alloing, G.; Becana, M. Thiol-based redox signaling in the nitrogen-fixing symbiosis. Front. Plant Sci. 2013, 4, e376. [Google Scholar] [CrossRef] [Green Version]
- Frendo, P.; Harrison, J.; Norman, C.; Jiménez, M.J.H.; van de Sype, G.; Gilabert, A.; Puppo, A. Glutathione and homoglutathione play a critical role in the nodulation process of Medicago truncatula. Mol. Plant Microbe Interact. 2005, 18, 254–259. [Google Scholar] [CrossRef] [Green Version]
- El Msehli, S.; Lambert, A.; Baldacci-Cresp, F.; Hopkins, J.; Boncompagni, E.; Smiti, S.A.; Hérouart, D.; Frendo, P. Crucial role of (homo)glutathione in nitrogen fixation in Medicago truncatula nodules. New Phytol. 2011, 192, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Si, Z.; Guan, N.; Zhou, Y.; Mei, L.; Li, Y.; Li, Y. A Methionine Sulfoxide Reductase B Is Required for the Establishment of Astragalus sinicus-Mesorhizobium Symbiosis. Plant Cell Physiol. 2020, 61, 1631–1645. [Google Scholar] [CrossRef] [PubMed]
- Mols, M.; Abee, T. Primary and secondary oxidative stress in Bacillus. Environ. Microbiol. 2011, 13, 1387–1394. [Google Scholar] [CrossRef]
- Imlay, J.A. The molecular mechanisms and physiological consequences of oxidative stress: Lessons from a model bacterium. Nat. Rev. Microbiol. 2013, 11, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reniere, M.L. Reduce, Induce, Thrive: Bacterial Redox Sensing during Pathogenesis. J. Bacteriol. 2018, 200, e00128-18. [Google Scholar] [CrossRef] [Green Version]
- D’Autréaux, B.; Toledano, M.B. ROS as signalling molecules: Mechanisms that generate specificity in ROS homeostasis. Nat. Rev. Mol. Cell Biol. 2007, 8, 813–824. [Google Scholar] [CrossRef]
- Ezraty, B.; Aussel, L.; Barras, F. Methionine sulfoxide reductases in prokaryotes. Biochim. Biophys. Acta Proteins Proteom. 2005, 1703, 221–229. [Google Scholar] [CrossRef]
- Mishra, S.; Imlay, J. Why do bacteria use so many enzymes to scavenge hydrogen peroxide? Arch. Biochem. Biophys. 2012, 525, 145–160. [Google Scholar] [CrossRef] [Green Version]
- Deponte, M. The Incomplete Glutathione Puzzle: Just Guessing at Numbers and Figures? Antioxid. Redox Signal. 2017, 27, 1130–1161. [Google Scholar] [CrossRef] [Green Version]
- Van Loi, V.; Rossius, M.; Antelmann, H. Redox regulation by reversible protein S-thiolation in bacteria. Front. Microbiol. 2015, 6, e187. [Google Scholar] [CrossRef] [Green Version]
- Tadra-Sfeir, M.Z.; Souza, E.M.; Faoro, H.; Müller-Santos, M.; Baura, V.A.; Tuleski, T.R.; Rigo, L.U.; Yates, M.G.; Wassem, R.; Pedrosa, F.O.; et al. Naringenin regulates expression of genes involved in cell wall synthesis in Herbaspirillum seropedicae. Appl. Environ. Microbiol. 2011, 77, 2180–2183. [Google Scholar] [CrossRef] [Green Version]
- Drogue, B.; Sanguin, H.; Borland, S.; Prigent-Combaret, C.; Wisniewski-Dyé, F. Genome wide profiling of Azospirillum lipoferum 4B gene expression during interaction with rice roots. FEMS Microbiol. Ecol. 2014, 87, 543–555. [Google Scholar] [CrossRef] [Green Version]
- Fukami, J.; Cerezini, P.; Hungria, M. Azospirillum: Benefits that go far beyond biological nitrogen fixation. AMB Express 2018, 8, 73. [Google Scholar] [CrossRef]
- Wisniewski-Dyé, F.; Lozano, L.; Acosta-Cruz, E.; Borland, S.; Drogue, B.; Prigent-Combaret, C.; Rouy, Z.; Barbe, V.; Herrera, A.M.; González, V.; et al. Genome sequence of Azospirillum brasilense CBG497 and comparative analyses of Azospirillum core and accessory genomes provide insight into niche adaptation. Genes 2012, 3, 576–602. [Google Scholar] [CrossRef] [PubMed]
- Camilios-Neto, D.; Bonato, P.; Wassem, R.; Tadra-Sfeir, M.Z.; Brusamarello-Santos, L.C.; Valdameri, G.; Donatti, L.; Faoro, H.; Weiss, V.A.; Chubatsu, L.S.; et al. Dual RNA-seq transcriptional analysis of wheat roots colonized by Azospirillum brasilense reveals up-regulation of nutrient acquisition and cell cycle genes. BMC Genom. 2014, 15, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasim, M.; Bible, A.N.; Xie, Z.; Alexandre, G. Alkyl hydroperoxide reductase has a role in oxidative stress resistance and in modulating changes in cell-surface properties in Azospirillum brasilense Sp245. Microbiology 2009, 155, 1192–1202. [Google Scholar] [CrossRef] [Green Version]
- Rai, A.K.; Singh, S.; Dwivedi, S.K.; Srivastava, A.; Pandey, P.; Kumar, S.; Singh, B.N.; Tripathi, A.K. Catalase Expression in Azospirillum brasilense Sp7 Is Regulated by a Network Consisting of OxyR and Two RpoH Paralogs and Including an RpoE1→RpoH5 Regulatory Cascade. Appl. Environ. Microbiol. 2018, 84, e01787-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battu, L.; Ulaganathan, K. Whole genome sequencing and identification of host-interactive genes in the rice endophytic Leifsonia sp. ku-ls. Funct. Integr. Genom. 2020, 20, 237–243. [Google Scholar] [CrossRef]
- Chaves, D.F.S.; de Souza, E.M.; Monteiro, R.A.; de Oliveira Pedrosa, F. A two-dimensional electrophoretic profile of the proteins secreted by Herbaspirillum seropedicae strain Z78. J. Proteom. 2009, 73, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Alquéres, S.M.C.; Oliveira, J.H.M.; Nogueira, E.M.; Guedes, H.V.; Oliveira, P.L.; Câmara, F.; Baldani, J.I.; Martins, O.B. Antioxidant pathways are up-regulated during biological nitrogen fixation to prevent ROS-induced nitrogenase inhibition in Gluconacetobacter diazotrophicus. Arch. Microbiol. 2010, 192, 835–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alquéres, S.; Meneses, C.; Rouws, L.; Rothballer, M.; Baldani, I.; Schmid, M.; Hartmann, A. The Bacterial Superoxide Dismutase and Glutathione Reductase Are Crucial for Endophytic Colonization of Rice Roots by Gluconacetobacter diazotrophicus PAL5. Mol. Plant Microbe Interact. 2013, 26, 937–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, L.P.S.; do Amaral, F.P.; Kim, D.; Bom, M.T.; Gavídia, M.P.; Teixeira, C.S.; Holthman, F.; de Oliveira Pedrosa, F.; de Souza, E.M.; Chubatsu, L.S.; et al. Importance of poly-3-hydroxybutyrate metabolism to the ability of Herbaspirillum seropedicae to promote plant growth. Appl. Environ. Microbiol. 2019, 85, e02586-18. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.; Herouart, D.; Puppo, A.; Touati, D. Critical protective role of bacterial superoxide dismutase in Rhizobium-legume symbiosis. Mol. Microbiol. 2000, 38, 750–759. [Google Scholar] [CrossRef] [Green Version]
- Jamet, A.; Sigaud, S.; van de Sype, G.; Puppo, A.; Hérouart, D. Expression of the bacterial catalase genes during Sinorhizobium meliloti-Medicago sativa symbiosis and their crucial role during the infection process. Mol. Plant Microbe Interact. 2003, 16, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Hanyu, M.; Fujimoto, H.; Tejima, K.; Saeki, K. Functional differences of two distinct catalases in Mesorhizobium loti MAFF303099 under free-living and symbiotic conditions. J. Bacteriol. 2009, 191, 1463–1471. [Google Scholar] [CrossRef] [Green Version]
- Dombrecht, B.; Heusdens, C.; Beullens, S.; Verreth, C.; Mulkers, E.; Proost, P.; Vanderleyden, J.; Michiels, J. Defence of Rhizobium etli bacteroids against oxidative stress involves a complexly regulated atypical 2-Cys peroxiredoxin. Mol. Microbiol. 2005, 55, 1207–1221. [Google Scholar] [CrossRef]
- Wang, S.; Lu, T.; Xue, Q.; Xu, K.; Cheng, G. Antioxidation and symbiotic nitrogen fixation function of prxA gene in Mesorhizobium huakuii. Microbiologyopen 2019, 8, e889. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Nickels, L.M.; Wang, H.; Ling, J.; Zhong, Z.; Zhu, J. OxyR-regulated catalase activity is critical for oxidative stress resistance, nodulation and nitrogen fixation in Azorhizobium caulinodans. FEMS Microbiol. Lett. 2016, 363, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Jiang, G.; Yang, J.; Li, X.; Cao, Y.; Liu, X.X.; Ling, J.; Wang, H.; Zhong, Z.; Zhu, J. Alkyl hydroperoxide reductase is important for oxidative stress resistance and symbiosis in Azorhizobium caulinodans. FEMS Microbiol. Lett. 2019, 366, 1–9. [Google Scholar] [CrossRef]
- Si, Y.; Guo, D.; Deng, S.; Lu, X.; Zhu, J.; Rao, B.; Cao, Y.; Jiang, G.; Yu, D.; Zhong, Z.; et al. Ohr and ohrR are critical for organic peroxide resistance and symbiosis in Azorhizobium caulinodans ORS571. Genes 2020, 11, 335. [Google Scholar] [CrossRef] [Green Version]
- Wheatley, R.M.; Ford, B.L.; Li, L.; Aroney, S.T.N.; Knights, H.E.; Ledermann, R.; East, A.K.; Ramachandran, V.K.; Poole, P.S. Lifestyle adaptations of Rhizobium from rhizosphere to symbiosis. Proc. Natl. Acad. Sci. USA 2020, 117, 23823–23834. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Karunakaran, R.; East, A.K.; Munoz-Azcarate, O.; Poole, P.S. Glutathione affects the transport activity of Rhizobium leguminosarum 3841 and is essential for efficient nodulation. FEMS Microbiol. Lett. 2017, 364, 1–7. [Google Scholar] [CrossRef]
- Harrison, J.; Jamet, A.; Muglia, C.I.; van de Sype, G.; Aguilar, O.M.; Puppo, A.; Frendo, P. Glutathione plays a fundamental role in growth and symbiotic capacity of Sinorhizobium meliloti. J. Bacteriol. 2005, 187, 168–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; El Msehli, S.; Benyamina, S.; Lambert, A.; Hopkins, J.; Cazareth, J.; Pierre, O.; Hérouart, D.; Achi-Smiti, S.; Boncompagni, E.; et al. Glutathione Deficiency in Sinorhizobium meliloti Does Not Impair Bacteroid Differentiation But Induces Early Senescence in the Interaction With Medicago truncatula. Front. Plant Sci. 2020, 11, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Taté, R.; Cermola, M.; Riccio, A.; Diez-Roux, G.; Patriarca, E.J. Glutathione Is Required by Rhizobium etli for Glutamine Utilization and Symbiotic Effectiveness. Mol. Plant Microbe Interact. 2012, 25, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Muglia, C.; Comai, G.; Spegazzini, E.; Riccillo, P.M.; Aguilar, O.M. Glutathione produced by Rhizobium tropici is important to prevent early senescence in common bean nodules. FEMS Microbiol. Lett. 2008, 286, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.; Yin, J.; Peng, Y.; Xie, J.; Wu, H.; He, D.; Li, X.; Cheng, G. Glutathione is Involved in Detoxification of Peroxide and Root Nodule Symbiosis of Mesorhizobium huakuii. Curr. Microbiol. 2020, 77, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.; Li, N.; Liu, Y.; Yu, L.; Yan, J.; Luo, L. Sinorhizobium meliloti Glutathione Reductase Is Required for both Redox Homeostasis and Symbiosis. Appl. Environ. Microbiol. 2017, 84, e01937-17. [Google Scholar] [CrossRef] [Green Version]
- Benyamina, S.M.; Baldacci-Cresp, F.; Couturier, J.; Chibani, K.; Hopkins, J.; Bekki, A.; de Lajudie, P.; Rouhier, N.; Jacquot, J.P.; Alloing, G.; et al. Two Sinorhizobium meliloti glutaredoxins regulate iron metabolism and symbiotic bacteroid differentiation. Environ. Microbiol. 2013, 15, 795–810. [Google Scholar] [CrossRef]
- Cao, Y.; Jiang, G.; Li, M.; Fang, X.; Zhu, D.; Qiu, W.; Zhu, J.; Yu, D.; Xu, Y.; Zhong, Z.; et al. Glutaredoxins Play an Important Role in the Redox Homeostasis and Symbiotic Capacity of Azorhizobium caulinodans ORS571. Mol. Plant Microbe Interact. 2020, 33, 1381–1393. [Google Scholar] [CrossRef]
- Wang, C.; Saldanha, M.; Sheng, X.; Shelswell, K.J.; Walsh, K.T.; Sobral, B.W.S.; Charles, T.C. Roles of poly-3-hydroxybutyrate (PHB) and glycogen in symbiosis of Sinorhizobium meliloti with Medicago sp. Microbiology 2007, 153, 388–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandon, K.; Michel-Reydellet, N.; Encarnación, S.; Kaminski, P.A.; Leija, A.; Cevallos, M.A.; Elmerich, C.; Mora, J. Poly-β-hydroxybutyrate turnover in Azorhizobium caulinodans is required for growth and affects nifA expression. J. Bacteriol. 1998, 180, 5070–5076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quelas, J.I.; Mongiardini, E.J.; Perez-Gimenez, J.; Parisi, G.; Lodeiro, A.R. Analysis of Two Polyhydroxyalkanoate Synthases in Bradyrhizobium japonicum USDA 110. J. Bacteriol. 2013, 195, 3145–3155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramachandran, V.K.; East, A.K.; Karunakaran, R.; Downie, J.A.; Poole, P.S. Adaptation of Rhizobium leguminosarum to pea, alfalfa and sugar beet rhizospheres investigated by comparative transcriptomics. Genome Biol. 2011, 12, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klonowska, A.; Melkonian, R.; Miché, L.; Tisseyre, P.; Moulin, L. Transcriptomic profiling of Burkholderia phymatum STM815, Cupriavidus taiwanensis LMG19424 and Rhizobium mesoamericanum STM3625 in response to Mimosa pudica root exudates illuminates the molecular basis of their nodulation competitiveness and symbiotic evolutionary history. BMC Genom. 2018, 19, 1–22. [Google Scholar] [CrossRef]
- Salas, M.E.; Lozano, M.J.; López, J.L.; Draghi, W.O.; Serrania, J.; Torres Tejerizo, G.A.; Albicoro, F.J.; Nilsson, J.F.; Pistorio, M.; del Papa, M.F.; et al. Specificity traits consistent with legume-rhizobia coevolution displayed by Ensifer meliloti rhizosphere colonization. Environ. Microbiol. 2017, 19, 3423–3438. [Google Scholar] [CrossRef] [Green Version]
- Puppo, A.; Pauly, N.; Boscari, A.; Mandon, K.; Brouquisse, R. Hydrogen Peroxide and Nitric Oxide: Key Regulators of the Legume—Rhizobium and Mycorrhizal Symbioses. Antioxid. Redox Signal. 2013, 18, 2202–2219. [Google Scholar] [CrossRef]
- Ribeiro, C.W.; Alloing, G.; Mandon, K.; Frendo, P. Redox regulation of differentiation in symbiotic nitrogen fixation. Biochim. Biophys. Acta Gen. Subj. 2015, 1850, 1469–1478. [Google Scholar] [CrossRef]
- Montiel, J.; Nava, N.; Cárdenas, L.; Sánchez-López, R.; Arthikala, M.K.; Santana, O.; Sánchez, F.; Quinto, C. A Phaseolus vulgaris NADPH Oxidase Gene is Required for Root Infection by Rhizobia. Plant Cell Physiol. 2012, 53, 1751–1767. [Google Scholar] [CrossRef] [Green Version]
- Tani, A.; Takai, Y.; Suzukawa, I.; Akita, M.; Murase, H.; Kimbara, K. Practical application of methanol-mediated mutualistic symbiosis between methylobacterium species and a roof greening moss, Racomitrium japonicum. PLoS ONE 2012, 7, 2–10. [Google Scholar] [CrossRef] [Green Version]
- Damiani, I.; Pauly, N.; Puppo, A.; Brouquisse, R.; Boscari, A. Reactive oxygen species and nitric oxide control early steps of the legume—Rhizobium symbiotic interaction. Front. Plant Sci. 2016, 7, 1–8. [Google Scholar] [CrossRef]
- Muñoz, V.; Ibáñez, F.; Figueredo, M.S.; Fabra, A. An oxidative burst and its attenuation by bacterial peroxidase activity is required for optimal establishment of the Arachis hypogaea-Bradyrhizobium sp. symbiosis. J. Appl. Microbiol. 2016, 121, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Barloy-Hubler, F.; Chéron, A.; Hellégouarch, A.; Galibert, F. Smc01944, a secreted peroxidase induced by oxidative stresses in Sinorhizobium meliloti 1021. Microbiology 2004, 150, 657–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queiroux, C.; Washburn, B.K.; Davis, O.M.; Stewart, J.; Brewer, T.E.; Lyons, M.R.; Jones, K.M. A comparative genomics screen identifies a Sinorhizobium meliloti 1021 sodM-like gene strongly expressed within host plant nodules. BMC Microbiol. 2012, 12, e74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krehenbrink, M.; Edwards, A.; Downie, J.A. The superoxide dismutase SodA is targeted to the periplasm in a SecA-dependent manner by a novel mechanism. Mol. Microbiol. 2011, 82, 164–179. [Google Scholar] [CrossRef] [PubMed]
- Asensio, A.C.; Marino, D.; James, E.K.; Ariz, I.; Arrese-Igor, C.; Aparicio-Tejo, P.M.; Arredondo-Peter, R.; Moran, J.F. Expression and localization of a Rhizobium-derived cambialistic superoxide dismutase in pea (Pisum sativum) nodules subjected to oxidative stress. Mol. Plant Microbe Interact. 2011, 24, 1247–1257. [Google Scholar] [CrossRef] [Green Version]
- Jeon, J.M.; Lee, H.I.; Donati, A.J.; So, J.S.; Emerich, D.W.; Chang, W.S. Whole-genome expression profiling of Bradyrhizobium japonicum in response to hydrogen peroxide. Mol. Plant Microbe Interact. 2011, 24, 1472–1481. [Google Scholar] [CrossRef] [Green Version]
- Djordjevic, M.A.; Chen, H.C.; Natera, S.; van Noorden, G.; Menzel, C.; Taylor, S.; Renard, C.; Geiger, O.; Weiller, G.F. A Global Analysis of Protein Expression Profiles in Sinorhizobium meliloti: Discovery of New Genes for Nodule Occupancy and Stress Adaptation. Mol. Plant Microbe Interact. 2003, 16, 508–524. [Google Scholar] [CrossRef] [Green Version]
- Fontenelle, C.; Blanco, C.; Arrieta, M.; Dufour, V.; Trautwetter, A. Resistance to organic hydroperoxides requires ohr and ohrR genes in Sinorhizobium meliloti. BMC Microbiol. 2011, 11, e100. [Google Scholar] [CrossRef] [Green Version]
- Lehman, A.P.; Long, R. OxyR-Dependent Transcription Response of Sinorhizobium meliloti to Oxidative Stress. J. Bacteriol. 2018, 200, e00622-17. [Google Scholar] [CrossRef] [Green Version]
- Panek, H.R.; O’Brian, M.R. KatG is the primary detoxifier of hydrogen peroxide produced by aerobic metabolism in Bradyrhizobium japonicum. J. Bacteriol. 2004, 186, 7874–7880. [Google Scholar] [CrossRef] [Green Version]
- Delmotte, N.; Ahrens, C.H.; Knief, C.; Qeli, E.; Koch, M.; Fischer, H.-M.; Vorholt, J.A.; Hennecke, H.; Pessi, G. An integrated proteomics and transcriptomics reference data set provides new insights into the Bradyrhizobium japonicum bacteroid metabolism in soybean root nodules. Proteomics 2010, 10, 1391–1400. [Google Scholar] [CrossRef]
- del Carmen Vargas, M.; Encarnación, S.; Dávalos, A.; Reyes-Pérez, A.; Mora, Y.; García-de los Santos, A.; Brom, S.; Mora, J. Only one catalase, katG, is detectable in Rhizobium etli, and is encoded along with the regulator OxyR on a plasmid replicon. Microbiology 2003, 149, 1165–1176. [Google Scholar] [CrossRef] [Green Version]
- Alloisio, N.; Queiroux, C.; Fournier, P.; Pujic, P.; Normand, P.; Vallenet, D.; Médigue, C.; Yamaura, M.; Kakoi, K.; Kucho, K.I. The frankia alni symbiotic transcriptome. Mol. Plant Microbe Interact. 2010, 23, 593–607. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, S.; Minamisawa, K.; Mitsui, H. A Sinorhizobium meliloti RpoH-regulated gene is involved in iron-sulfur protein metabolism and effective plant symbiosis under intrinsic iron limitation. J. Bacteriol. 2016, 198, 2297–2306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, S.; Aono, T.; Lee, K.B.; Suzuki, T.; Liu, C.T.; Miwa, H.; Wakao, S.; Iki, T.; Oyaizu, H. Rhizobial factors required for stem nodule maturation and maintenance in Sesbania rostrata-Azorhizobium caulinodans ORS571 symbiosis. Appl. Environ. Microbiol. 2007, 73, 6650–6659. [Google Scholar] [CrossRef] [Green Version]
- Hillion, M.; Antelmann, H. Thiol-based redox switches in prokaryotes. Biol. Chem. 2015, 396, 415–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamet, A.; Kiss, E.; Batut, J.; Puppo, A.; Hérouart, D. The katA catalase gene is regulated by OxyR in both free-living and symbiotic Sinorhizobium meliloti. J. Bacteriol. 2005, 187, 376–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakjarung, K.; Mongkolsuk, S.; Vattanaviboon, P. The oxyR from Agrobacterium tumefaciens: Evaluation of its role in the regulation of catalase and peroxide responses. Biochem. Biophys. Res. Commun. 2003, 304, 41–47. [Google Scholar] [CrossRef]
- Luo, L.; Yao, S.-Y.; Becker, A.; Rüberg, S.; Yu, G.-Q.; Zhu, J.-B.; Cheng, H.-P. Two New Sinorhizobium meliloti LysR-type Transcriptional Regulators Required for Nodulation. J. Bacteriol. 2005, 187, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, G.; Wang, Y.; Luo, L. Transcriptional regulator LsrB of Sinorhizobium meliloti positively regulates the expression of genes involved in lipopolysaccharide biosynthesis. Appl. Environ. Microbiol. 2014, 80, 5265–5273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, G.; Xing, S.; Wang, S.; Yu, L.; Li, X.; Staehelin, C.; Yang, M.; Luo, L. Regulation of cysteine residues in LsrB proteins from Sinorhizobium meliloti under free-living and symbiotic oxidative stress. Environ. Microbiol. 2017, 19, 5130–5145. [Google Scholar] [CrossRef]
- Wilkinson, S.P.; Grove, A. Ligand-responsive transcriptional regulation by members of the MarR family of winged helix proteins. Curr. Issues Mol. Biol. 2006, 8, 51–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roux, B.; Rodde, N.; Jardinaud, M.F.; Timmers, T.; Sauviac, L.; Cottret, L.; Carrère, S.; Sallet, E.; Courcelle, E.; Moreau, S.; et al. An integrated analysis of plant and bacterial gene expression in symbiotic root nodules using laser-capture microdissection coupled to RNA sequencing. Plant J. 2014, 77, 817–837. [Google Scholar] [CrossRef] [PubMed]
- Oke, V.; Rushing, B.G.; Fisher, E.J.; Moghadam-Tabrizi, M.; Long, S.R. Identification of the heat-shock sigma factor RpoH and a second RpoH-like protein in Sinorhizobium meliloti. Microbiology 2001, 147, 2399–2408. [Google Scholar] [CrossRef] [Green Version]
- De Lucena, D.K.C.; Pühler, A.; Weidner, S. The role of sigma factor RpoH1 in the pH stress response of Sinorhizobium meliloti. BMC Microbiol. 2010, 10, e265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnett, M.J.; Bittner, A.N.; Toman, C.J.; Oke, V.; Long, S.R. Dual RpoH sigma factors and transcriptional plasticity in a symbiotic bacterium. J. Bacteriol. 2012, 194, 4983–4994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Salazar, J.M.; Sandoval-Calderón, M.; Guo, X.; Castillo-Ramírez, S.; Reyes, A.; Loza, M.G.; Rivera, J.; Alvarado-Affantranger, X.; Sánchez, F.; González, V.; et al. The Rhizobium etli RpoH1 and RpoH2 sigma factors are involved in different stress responses. Microbiology 2009, 155, 386–397. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N.; Gupta, A.; Kumar, S.; Mishra, R.; Singh, C.; Tripathi, A.K. Cross-Talk Between Cognate and Noncognate RpoE Sigma Factors and Zn2+-Binding Anti-Sigma Factors Regulates Photooxidative Stress Response in Azospirillum brasilense. Antioxid. Redox Signal. 2014, 20, 42–59. [Google Scholar] [CrossRef]
- Sauviac, L.; Philippe, H.; Phok, K.; Bruand, C. An extracytoplasmic function sigma factor acts as a general stress response regulator in Sinorhizobium meliloti. J. Bacteriol. 2007, 189, 4204–4216. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Salazar, J.M.; Salazar, E.; Encarnación, S.; Ramírez-Romero, M.A.; Rivera, J. Role of the extracytoplasmic function sigma factor RpoE4 in oxidative and osmotic stress responses in Rhizobium etli. J. Bacteriol. 2009, 191, 4122–4132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flechard, M.; Fontenelle, C.; Trautwetter, A.; Ermel, G.; Blanco, C. Sinorhizobium meliloti rpoE2 is necessary for H2O2 stress resistance during the stationary growth phase. FEMS Microbiol. Lett. 2009, 290, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, N.; Kumar, S.; Mishra, M.N.; Tripathi, A.K. A constitutively expressed pair of rpoE2–chrR2 in Azospirillum brasilense Sp7 is required for survival under antibiotic and oxidative stress. Microbiology 2013, 159, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Rai, A.K.; Dubey, A.P.; Kumar, S.; Dutta, D.; Mishra, M.N.; Singh, B.N.; Tripathi, A.K. Carotenoid Biosynthetic Pathways Are Regulated by a Network of Multiple Cascades of Alternative Sigma Factors in Azospirillum brasilense Sp7. J. Bacteriol. 2016, 198, 2955–2964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Dwivedi, S.K.; Singh, V.S.; Tripathi, A.K. Expression of alkyl hydroperoxide reductase is regulated negatively by OxyR1 and positively by RpoE2 sigma factor in Azospirillum brasilense Sp7. Microbiology 2016, 162, 1870–1883. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Rees, D.C. Nitrogenase and Biological Nitrogen Fixation. Biochemistry 1994, 33, 389–397. [Google Scholar] [CrossRef]
- Rutten, P.J.; Poole, P.S. Oxygen regulatory mechanisms of nitrogen fixation in rhizobia. In Advances in Microbial Physiology; Elsevier Ltd.: Amsterdam, The Netherlands, 2019; Volume 75, pp. 325–389. ISBN 9780128177143. [Google Scholar]
- Hill, S.; Austin, S.; Eydmann, T.; Jones, T.; Dixon, R. Azotobacter vinelandii NIFL is a flavoprotein that modulates transcriptional activation of nitrogen-fixation genes via a redox-sensitive switch. Proc. Natl. Acad. Sci. USA 1996, 93, 2143–2148. [Google Scholar] [CrossRef] [Green Version]
- Das, H.K. Azotobacters as biofertilizer. Adv. Appl. Microbiol. 2019, 108, 1–43. [Google Scholar] [CrossRef]
- Grabbe, R.; Schmitz, R.A. Oxygen control of nif gene expression in Klebsiella pneumoniae depends on NifL reduction at the cytoplasmic membrane by electrons derived from the reduced quinone pool. Eur. J. Biochem. 2003, 270, 1555–1566. [Google Scholar] [CrossRef] [Green Version]
- Fischer, H.M.; Bruderer, T.; Hennecke, H. Essential and non-essential domains in the Bradyrhizobium japonicum NifA protein: Identification of indispensable cysteine residues potentially involved in redox reactivity and/or metal binding. Nucleic Acids Res. 1988, 16, 2207–2224. [Google Scholar] [CrossRef]
- Honorio-Felício, N.; Carepo, M.S.P.; de F Paulo, T.; de França Lopes, L.G.; Sousa, E.H.S.; Diógenes, I.C.N.; Bernhardt, P.V. The Heme-Based Oxygen Sensor Rhizobium etli FixL: Influence of Auxiliary Ligands on Heme Redox Potential and Implications on the Enzyme Activity. J. Inorg. Biochem. 2016, 164, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Mesa, S.; Reutimann, L.; Fischer, H.M.; Hennecke, H. Posttranslational control of transcription factor FixK2, a key regulator for the Bradyrhizobium japonicum—Soybean symbiosis. Proc. Natl. Acad. Sci. USA 2009, 106, 21860–21865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, E.; Kaspar, T.; Fischer, H.-M.; Hennecke, H. Expression of the fixR-nifA Operon in Bradyrhizobium japonicum Depends on a New Response Regulator, RegR. J. Bacteriol. 1998, 180, 3853–3863. [Google Scholar] [CrossRef] [Green Version]
- Bueno, E.; Mesa, S.; Bedmar, E.J.; Richardson, D.J.; Delgado, M.J. Bacterial Adaptation of Respiration from Oxic to Microoxic and Anoxic Conditions: Redox Control. Antioxid. Redox Signal. 2012, 16, 819–852. [Google Scholar] [CrossRef]
- Oger, E.; Marino, D.; Guigonis, J.M.; Pauly, N.; Puppo, A. Sulfenylated proteins in the Medicago truncatula-Sinorhizobium meliloti symbiosis. J. Proteom. 2012, 75, 4102–4113. [Google Scholar] [CrossRef] [PubMed]
- Udvardi, M.; Poole, P.S. Transport and Metabolism in Legume-Rhizobia Symbioses. Annu. Rev. Plant Biol. 2013, 64, 781–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadouri, D.; Burdman, S.; Jurkevitch, E.; Okon, Y. Identification and isolation of genes involved in poly(β-hydroxybutyrate) biosynthesis in Azospirillum brasilense and characterization of a phbC mutant. Appl. Environ. Microbiol. 2002, 68, 2943–2949. [Google Scholar] [CrossRef] [Green Version]
- Batista, M.B.; Teixeira, C.S.; Sfeir, M.Z.T.T.; Alves, L.P.S.S.; Valdameri, G.; de Oliveira Pedrosa, F.; Sassaki, G.L.; Steffens, M.B.R.; de Souza, E.M.; Dixon, R.; et al. PHB biosynthesis counteracts redox stress in Herbaspirillum seropedicae. Front. Microbiol. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cevallos, M.A.; Encarnacion, S.; Leija, A.; Mora, Y.; Mora, J. Genetic and physiological characterization of a Rhizobium etli mutant strain unable to synthesize poly-β-hydroxybutyrate. J. Bacteriol. 1996, 178, 1646–1654. [Google Scholar] [CrossRef] [Green Version]
- Walshaw, D.L.; Wilkinson, A.; Mundy, M.; Smith, M.; Poole, P.S. Regulation of the TCA cycle and the general amino acid permease by overflow metabolism in Rhizobium leguminosarum. Microbiology 1997, 143, 2209–2221. [Google Scholar] [CrossRef] [Green Version]
- Kadouri, D.; Jurkevitch, E.; Okon, Y. Involvement of the reserve material poly-β-hydroxybutyrate in Azospirillum brasilense stress endurance and root colonization. Appl. Environ. Microbiol. 2003, 69, 3244–3250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trainer, M.A.; Charles, T.C. The role of PHB metabolism in the symbiosis of rhizobia with legumes. Appl. Microbiol. Biotechnol. 2006, 71, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Terpolilli, J.J.; Masakapalli, S.K.; Karunakaran, R.; Webb, I.U.C.C.; Green, R.; Watmough, N.J.; Kruger, N.J.; Ratcliffe, R.G.; Poole, P.S. Lipogenesis and redox balance in nitrogen-fixing pea bacteroids. J. Bacteriol. 2016, 198, 2864–2875. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Genes | Bacterium | Mutant Phenotype In planta | References |
|---|---|---|---|
| ENDOPHYTES | |||
| sod | G. diazotrophicus | impaired root colonization | [62] |
| gr | G. diazotrophicus | impaired root colonization | [62] |
| phbC | H. seropedicae | loss of plant growth promoting ability | [63] |
| ENDOSYMBIONTS | |||
| ROS scavenging | |||
| sodA | S. meliloti | depending on the S. meliloti strain genotype | [64] |
| katB | S. meliloti | delayed nodulation; aberrant and enlarged infection threads | [28] |
| katB/katC | S. meliloti | altered bacteroid differentiation | [65] |
| katA/katC | S. meliloti | early nodule senescence | [65] |
| katE | M. loti | lower nitrogen fixation capacity | [66] |
| katG/prxS | R. etli | lower nitrogen fixation capacity | [67] |
| prxA | M. huakii | lower nitrogen fixation capacity | [68] |
| katG | A. caulinodans | decreased number and nitrogenase activity of stem nodules | [69] |
| ahpCD | A. caulinodans | decreased number and nitrogenase activity of stem nodules | [70] |
| ohr | A. caulinodans | decreased number and nitrogenase activity of stem nodules | [71] |
| GSH/Grx system | |||
| gshA | R. leguminosarum | impaired root colonization, lower nitrogen fixation capacity | [72] |
| gshB | R. leguminosarum | impaired root colonization, lower nitrogen fixation capacity | [72,73] |
| gshB | S. meliloti | delayed nodulation, early nodule senescence | [74,75] |
| gshB | R. etli | delayed nodulation, early nodule senescence | [76] |
| gshB | R. tropici | low competitiveness for nodule occupancy, early nodule senescence | [77] |
| gshB | M. huakii | early nodule senescence | [78] |
| gor | S. meliloti | low competitiveness for nodule occupancy, lower nitrogen fixation capacity | [79] |
| grx (RL2615) | R. leguminosarum | impaired root colonization | [72] |
| grx1 | S. meliloti | altered bacteroid differentiation | [80] |
| grx2 | S. meliloti | early nodule senescence | [80] |
| grx1/grx2 | A. caulinodans | lower nitrogen fixation capacity | [81] |
| grx3/grx4 | A. caulinodans | lower nitrogen fixation capacity | [81] |
| PHB metabolism | |||
| phbC | S. meliloti | lower nitrogen fixation capacity | [82] |
| phbC | A. caulinodans | loss of nitrogen fixation ability | [83] |
| phbC | B. japonicum | low competitiveness for root colonization | [84] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mandon, K.; Nazaret, F.; Farajzadeh, D.; Alloing, G.; Frendo, P. Redox Regulation in Diazotrophic Bacteria in Interaction with Plants. Antioxidants 2021, 10, 880. https://doi.org/10.3390/antiox10060880
Mandon K, Nazaret F, Farajzadeh D, Alloing G, Frendo P. Redox Regulation in Diazotrophic Bacteria in Interaction with Plants. Antioxidants. 2021; 10(6):880. https://doi.org/10.3390/antiox10060880
Chicago/Turabian StyleMandon, Karine, Fanny Nazaret, Davoud Farajzadeh, Geneviève Alloing, and Pierre Frendo. 2021. "Redox Regulation in Diazotrophic Bacteria in Interaction with Plants" Antioxidants 10, no. 6: 880. https://doi.org/10.3390/antiox10060880
APA StyleMandon, K., Nazaret, F., Farajzadeh, D., Alloing, G., & Frendo, P. (2021). Redox Regulation in Diazotrophic Bacteria in Interaction with Plants. Antioxidants, 10(6), 880. https://doi.org/10.3390/antiox10060880