
Blood Pressure-Lowering Effect of Wine Lees Phenolic Compounds Is Mediated by Endothelial-Derived Factors: Role of Sirtuin 1

 ,
,  , ,
, ,  ,
,  , and
, and 
Abstract
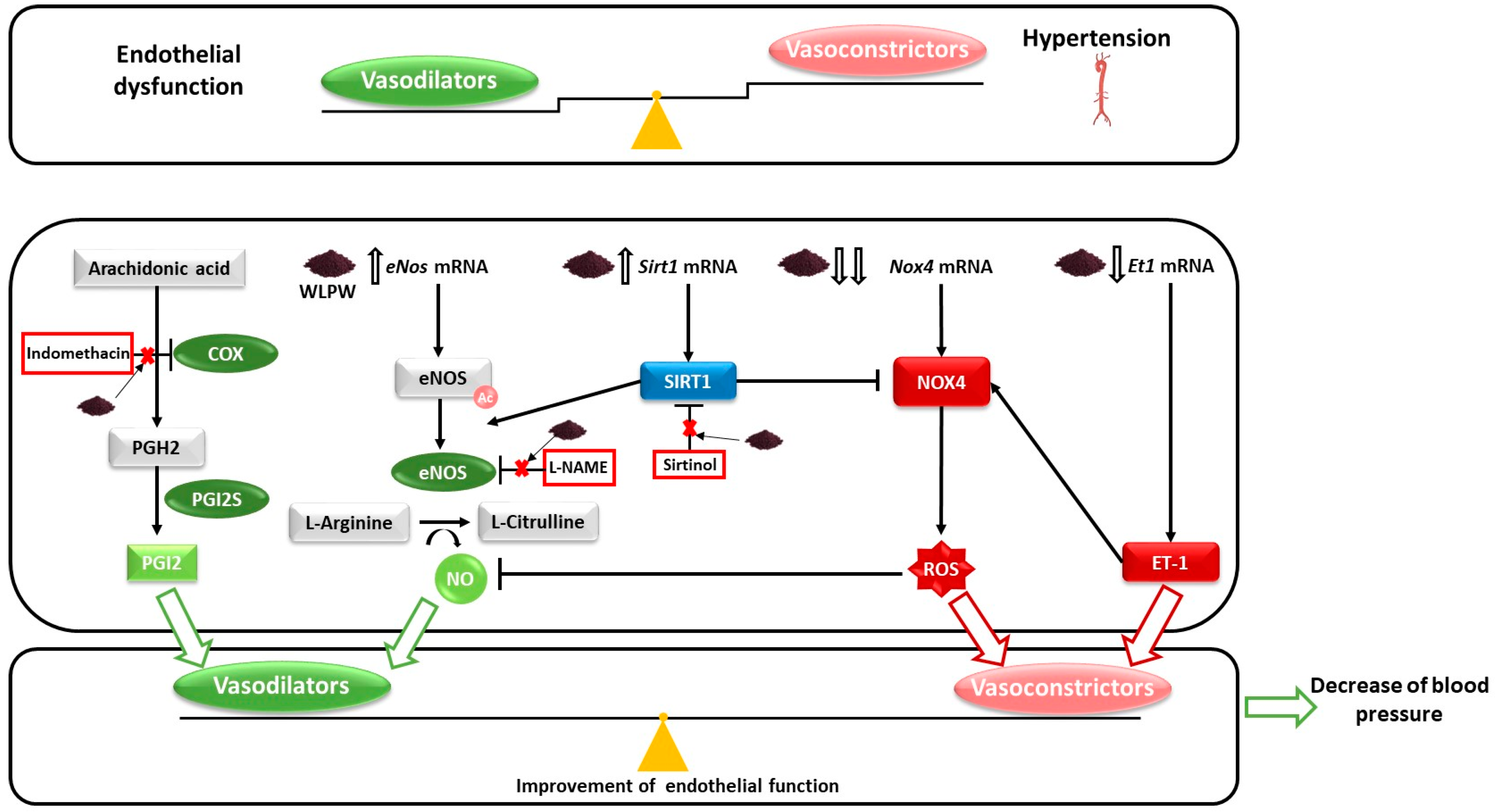
:
1. Introduction
2. Materials and Methods
2.1. Obtaining and Characterization of Wine Lees
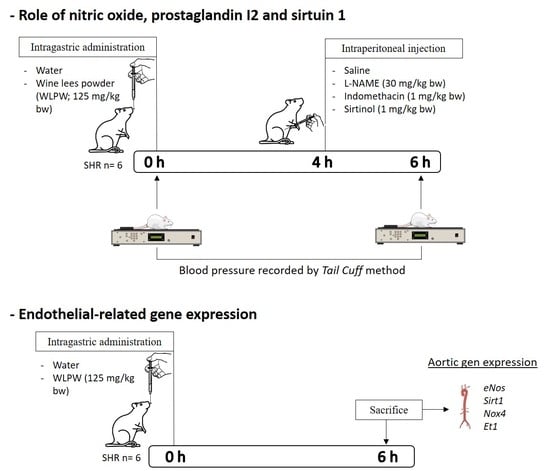
2.2. Dosage Regimen and Experimental Procedure in Animal
2.3. RNA Extraction and mRNA Quantification by Real-Time qPCR
2.4. Statistical Analysis
3. Results
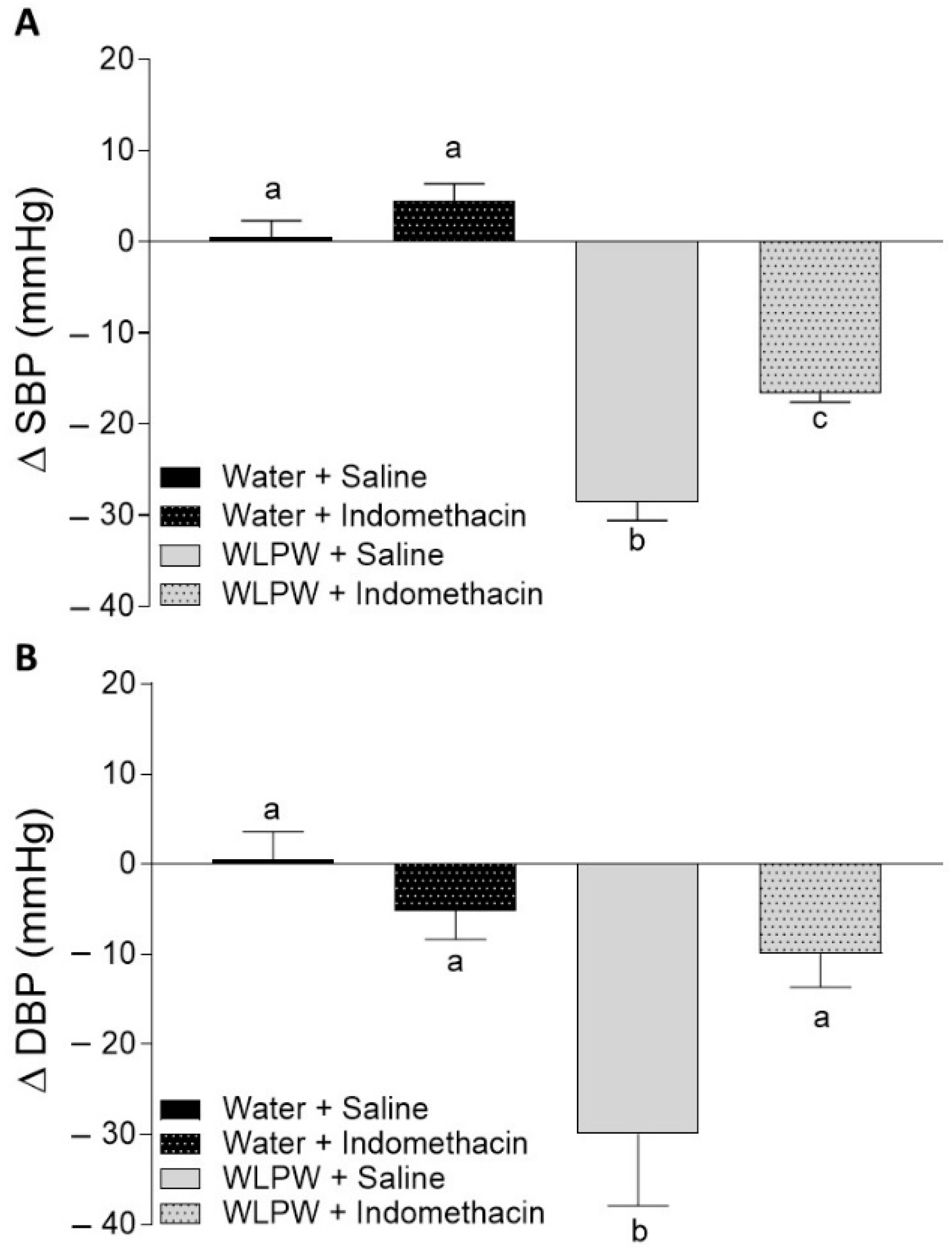
3.1. Role of Nitric Oxide, Prostacyclin and Sirtuin in the BP-Lowering Effect of WLPW
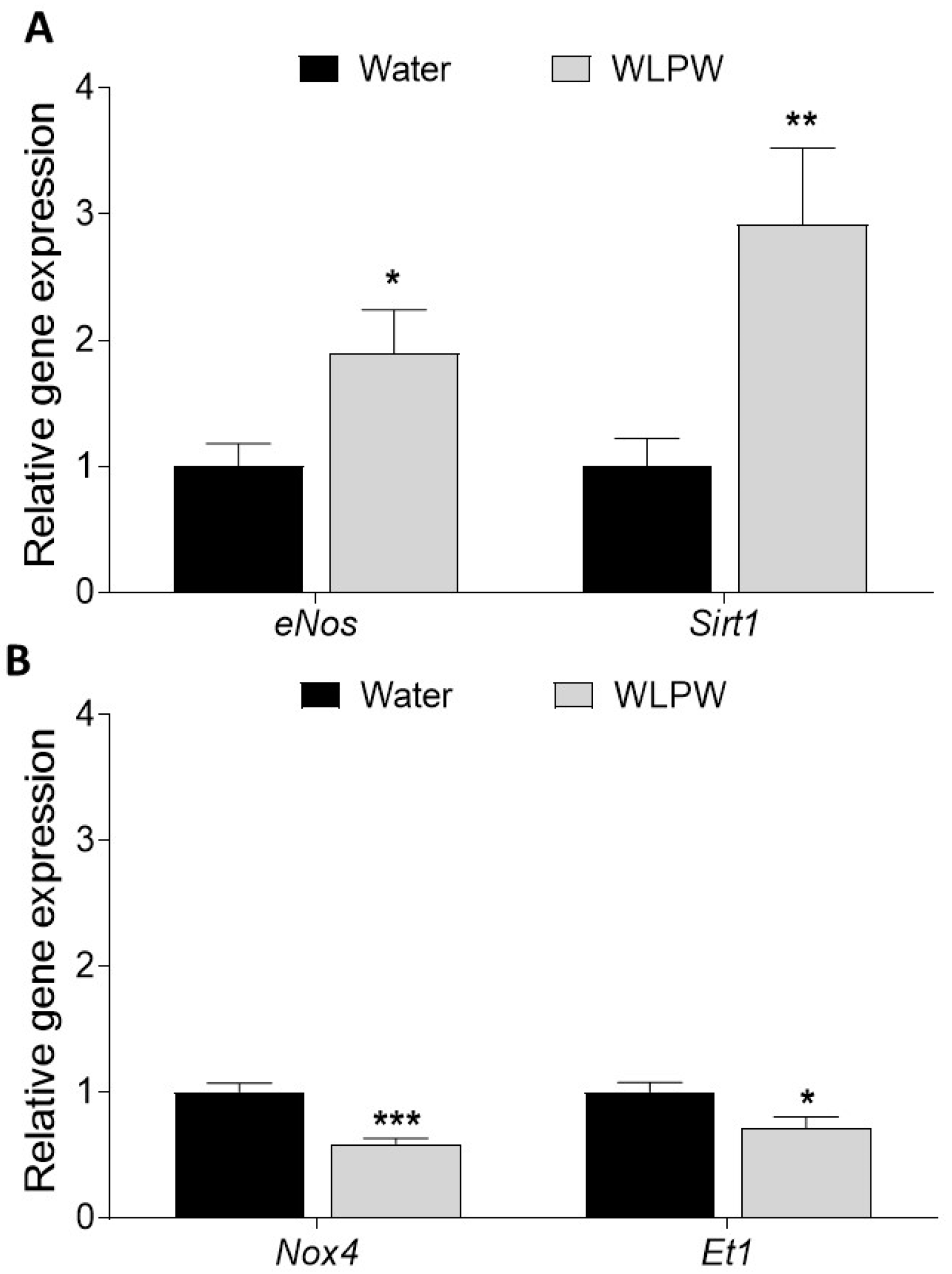
3.2. Endothelial-Related Gene Expression
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spieker, L.E.; Flammer, A.J.; Lüscher, T.F. The vascular endothelium in hypertension. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2006; pp. 249–283. [Google Scholar] [CrossRef]
- Konukoglu, D.; Uzun, H. Endothelial dysfunction and hypertension. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2016; Volume 956, pp. 511–540. [Google Scholar]
- Vallance, P.; Collier, J.; Moncada, S. Effects of Endothelium-Derived Nitric Oxide on Peripheral Arteriolar Tone in Man. Lancet 1989, 334, 997–1000. [Google Scholar] [CrossRef]
- Förstermann, U.; Sessa, W.C. Nitric Oxide Synthases: Regulation and Function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bucci, M.; Gratton, J.-P.; Rudic, R.D.; Acevedo, L.; Roviezzo, F.; Cirino, G.; Sessa, W.C. In Vivo Delivery of the Caveolin-1 Scaffolding Domain Inhibits Nitric Oxide Synthesis and Reduces Inflammation. Nat. Med. 2000, 6, 1362–1367. [Google Scholar] [CrossRef] [PubMed]
- Mattagajasingh, I.; Kim, C.S.; Naqvi, A.; Yamamori, T.; Hoffman, T.A.; Jung, S.B.; DeRicco, J.; Kasuno, K.; Irani, K. SIRT1 Promotes Endothelium-Dependent Vascular Relaxation by Activating Endothelial Nitric Oxide Synthase. Proc. Natl. Acad. Sci. USA 2007, 104, 14855–14860. [Google Scholar] [CrossRef] [Green Version]
- Man, A.W.C.; Li, H.; Xia, N. The Role of Sirtuin1 in Regulating Endothelial Function, Arterial Remodeling and Vascular Aging. Front. Physiol. 2019, 10, 1173. [Google Scholar] [CrossRef] [Green Version]
- Zarzuelo, M.J.; López-Sepúlveda, R.; Sánchez, M.; Romero, M.; Gómez-Guzmán, M.; Ungvary, Z.; Pérez-Vizcaíno, F.; Jiménez, R.; Duarte, J. SIRT1 Inhibits NADPH Oxidase Activation and Protects Endothelial Function in the Rat Aorta: Implications for Vascular Aging. Biochem. Pharmacol. 2013, 85, 1288–1296. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Luo, N.; Chi, Y. Aging-Shifted Prostaglandin Profile in Endothelium as a Factor in Cardiovascular Disorders. J. Aging Res. 2012, 2012, 121390. [Google Scholar] [CrossRef] [Green Version]
- Sandoo, A.; Veldhuijzen van Zanten, J.J.C.; Metsios, G.S.; Carroll, D.; Kitas, G.D. The Endothelium and Its Role in Regulating Vascular Tone. Open Cardiovasc. Med. J. 2015, 4, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.A.; Kirkby, N.S. Eicosanoids, Prostacyclin and Cyclooxygenase in the Cardiovascular System. Br. J. Pharmacol. 2019, 176, 1038–1050. [Google Scholar] [CrossRef] [PubMed]
- Iglarz, M.; Schiffrin, E.L. Role of Endothelin-1 in Hypertension. Curr. Hypertens. Rep. 2003, 5, 144–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stauffer, B.L.; Westby, C.M.; DeSouza, C.A. Endothelin-1, Aging and Hypertension. Curr. Opin. Cardiol. 2008, 23, 350–355. [Google Scholar] [CrossRef] [Green Version]
- López-Fernández-Sobrino, R.; Soliz-Rueda, J.R.; Margalef, M.; Arola-Arnal, A.; Suárez, M.; Bravo, F.I.; Muguerza, B. ACE Inhibitory and Antihypertensive Activities of Wine Lees and Relationship among Bioactivity and Phenolic Profile. Nutrients 2021, 13, 679. [Google Scholar] [CrossRef]
- López-Fernández-Sobrino, R.; Soliz-Rueda, J.R.; Suárez, M.; Mulero, M.; Arola, L.; Bravo, F.I.; Muguerza, B. Blood Pressure-Lowering Effect of Wine Lees: Dose-Response Study, Effect of Dealcoholization and Possible Mechanisms of Action. Nutrients 2021, 13, 1142. [Google Scholar] [CrossRef]
- Kondrashov, A.; Vranková, S.; Dovinová, I.; Ševčík, R.; Parohová, J.; Barta, A.; Pecháňová, O.; Kovacsová, M. The Effects of New Alibernet Red Wine Extract on Nitric Oxide and Reactive Oxygen Species Production in Spontaneously Hypertensive Rats. Oxid. Med. Cell. Longev. 2012, 2012, 806285. [Google Scholar] [CrossRef] [PubMed]
- Mas-Capdevila, A.; Iglesias-Carres, L.; Arola-Arnal, A.; Suárez, M.; Bravo, F.I.; Muguerza, B. Changes in Arterial Blood Pressure Caused by Long-Term Administration of Grape Seed Proanthocyanidins in Rats with Established Hypertension. Food Funct. 2020, 11, 8735–8742. [Google Scholar] [CrossRef] [PubMed]
- Quiñones, M.; Miguel, M.; Muguerza, B.; Aleixandre, A. Effect of a Cocoa Polyphenol Extract in Spontaneously Hypertensive Rats. Food Funct. 2011, 2, 649–653. [Google Scholar] [CrossRef]
- Rodríguez-Monforte, M.; Flores-Mateo, G.; Sánchez, E. Dietary Patterns and CVD: A Systematic Review and Meta-Analysis of Observational Studies. Br. J. Nutr. 2013, 114, 1341–1359. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Li, F.; Wang, L.; Zhang, D. Fruit and Vegetables Consumption and Risk of Hypertension: A Meta-Analysis. J. Clin. Hypertens. 2016, 18, 468–476. [Google Scholar] [CrossRef]
- Fraga, C.G.; Croft, K.D.; Kennedy, D.O.; Tomás-Barberán, F.A. The Effects of Polyphenols and Other Bioactives on Human Health. Food Funct. 2019, 10, 514–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.-N.; Meng, X.; Li, Y.; Li, S.; Liu, Q.; Tang, G.-Y.; Li, H.-B. Fruits for Prevention and Treatment of Cardiovascular Diseases. Nutrients 2017, 9, 598. [Google Scholar] [CrossRef] [Green Version]
- Pons, Z.; Margalef, M.; Bravo, F.I.; Arola-Arnal, A.; Muguerza, B. Grape Seed Flavanols Decrease Blood Pressure via Sirt-1 and Confer a Vasoprotective Pattern in Rats. J. Funct. Foods 2016, 24, 164–172. [Google Scholar] [CrossRef]
- Quiñones, M.; Guerrero, L.; Fernández-Vallinas, S.; Pons, Z.; Arola, L.; Aleixandre, A.; Muguerza, B. Involvement of Nitric Oxide and Prostacyclin in the Antihypertensive Effect of Low-Molecular-Weight Procyanidin Rich Grape Seed Extract in Male Spontaneously Hypertensive Rats. J. Funct. Foods 2014, 6, 419–427. [Google Scholar] [CrossRef]
- Kang, K.T. Endothelium-Derived Relaxing Factors of Small Resistance Arteries in Hypertension. Toxicol. Res. 2014, 30, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Roth, I.; Casas, R.; Ribó-Coll, M.; Estruch, R. Consumption of Aged White Wine under a Veil of Flor Reduces Blood Pressure-Increasing Plasma Nitric Oxide in Men at High Cardiovascular Risk. Nutrients 2019, 11, 1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pons, Z.; Margalef, M.; Bravo, F.I.; Arola-Arnal, A.; Muguerza, B. Acute Administration of Single Oral Dose of Grape Seed Polyphenols Restores Blood Pressure in a Rat Model of Metabolic Syndrome: Role of Nitric Oxide and Prostacyclin. Eur. J. Nutr. 2015, 55, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Quiñones, M.; Muguerza, B.; Miguel, M.; Aleixandre, A. Evidence That Nitric Oxide Mediates the Blood Pressure Lowering Effect of a Polyphenol-Rich Cocoa Powder in Spontaneously Hypertensive Rats. Pharmacol. Res. 2011, 64, 478–481. [Google Scholar] [CrossRef]
- Rodriguez-Mateos, A.; Istas, G.; Boschek, L.; Feliciano, R.P.; Mills, C.E.; Boby, C.; Gomez-Alonso, S.; Milenkovic, D.; Heiss, C. Circulating Anthocyanin Metabolites Mediate Vascular Benefits of Blueberries: Insights from Randomized Controlled Trials, Metabolomics, and Nutrigenomics. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2019, 74, 967–976. [Google Scholar] [CrossRef]
- Fairlie-Jones, L.; Davison, K.; Fromentin, E.; Hill, A. The Effect of Anthocyanin-Rich Foods or Extracts on Vascular Function in Adults: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. Nutrients 2017, 9, 908. [Google Scholar] [CrossRef] [Green Version]
- Calfío, C.; Huidobro-Toro, J.P. Potent Vasodilator and Cellular Antioxidant Activity of Endemic Patagonian Calafate Berries (Berberis Microphylla) with Nutraceutical Potential. Molecules 2019, 24, 2700. [Google Scholar] [CrossRef] [Green Version]
- Potente, M.; Dimmeler, S. NO Targets SIRT1. Arter. Thromb. Vasc. Biol. 2008, 28, 1577–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kratz, E.M.; Sołkiewicz, K.; Kubis-Kubiak, A.; Piwowar, A. Sirtuins as Important Factors in Pathological States and the Role of Their Molecular Activity Modulators. Int. J. Mol. Sci. 2021, 22, 630. [Google Scholar] [CrossRef]
- Kitada, M.; Ogura, Y.; Monno, I.; Koya, D. Supplementation with Red Wine Extract Increases Insulin Sensitivity and Peripheral Blood Mononuclear Sirt1 Expression in Nondiabetic Humans. Nutrients 2020, 12, 3108. [Google Scholar] [CrossRef] [PubMed]
- Ray, R.; Murdoch, C.E.; Wang, M.; Santos, C.X.; Zhang, M.; Alom-Ruiz, S.; Anilkumar, N.; Ouattara, A.; Cave, A.C.; Walker, S.J.; et al. Endothelial Nox4 NADPH Oxidase Enhances Vasodilatation and Reduces Blood Pressure in Vivo. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 1368–1376. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.I.; Griendling, K.K. Regulation of Signal Transduction by Reactive Oxygen Species in the Cardiovascular System. Circ. Res. 2015, 116, 531–549. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.; Swei, A.; Zweifach, B.W.; Schmid-Schönbein, G.W. In Vivo Evidence for Microvascular Oxidative Stress in Spontaneously Hypertensive Rats: Hydroethidine Microfluorography. Hypertension 1995, 25, 1083–1089. [Google Scholar] [CrossRef]
- Negishi, H.; Njelekela, M.; Ikeda, K.; Sagara, M.; Noguchi, T.; Kuga, S.; Kanda, T.; Liu, L.; Nara, Y.; Tagami, M.; et al. Assessment of in Vivo Oxidative Stress in Hypertensive Rats and Hypertensive Subjects in Tanzania, Africa. Hypertension Res. 2000, 23, 285–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.; Hutabarat, R.P.; Chai, Z.; Zheng, T.; Zhang, W.; Li, D. Antioxidant Blueberry Anthocyanins Induce Vasodilation via PI3K/Akt Signaling Pathway in High-glucose-induced Human Umbilical Vein Endothelial Cells. Int. J. Mol. Sci. 2020, 21, 1575. [Google Scholar] [CrossRef] [Green Version]
- Galindo, P.; González-Manzano, S.; Zarzuelo, M.J.; Gómez-Guzmán, M.; Quintela, A.M.; González-Paramás, A.; Santos-Buelga, C.; Pérez-Vizcaíno, F.; Duarte, J.; Jiménez, R. Different Cardiovascular Protective Effects of Quercetin Administered Orally or Intraperitoneally in Spontaneously Hypertensive Rats. Food Funct. 2012, 3, 643–650. [Google Scholar] [CrossRef]
- Schiffrin, E.L. Role of Endothelin-1 in Hypertension and Vascular Disease. Am. J. Hypertens. 2001, 14, 83S–89S. [Google Scholar] [CrossRef] [Green Version]
- Markovics, A.; Biró, A.; Kun-Nemes, A.; Fazekas, M.É.; Rácz, A.A.; Paholcsek, M.; Lukács, J.; Stündl, L.; Remenyik, J. Effect of Anthocyanin-Rich Extract of Sour Cherry for Hyperglycemia-Induced Inflammatory Response and Impaired Endothelium-Dependent Vasodilation. Nutrients 2020, 12, 3373. [Google Scholar] [CrossRef] [PubMed]
- Lazzè, M.C.; Pizzala, R.; Perucca, P.; Cazzalini, O.; Savio, M.; Forti, L.; Vannini, V.; Bianchi, L. Anthocyanidins Decrease Endothelin-1 Production and Increase Endothelial Nitric Oxide Synthase in Human Endothelial Cells. Mol. Nutr. Food Res. 2006, 50, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, R.; López-Sepúlveda, R.; Kadmiri, M.; Romero, M.; Vera, R.; Sánchez, M.; Vargas, F.; O’Valle, F.; Zarzuelo, A.; Dueñas, M.; et al. Polyphenols Restore Endothelial Function in DOCA-Salt Hypertension: Role of Endothelin-1 and NADPH Oxidase. Free Radic. Biol. Med. 2007, 43, 462–473. [Google Scholar] [CrossRef]
- López-Sepúlveda, R.; Jiménez, R.; Romero, M.; Zarzuelo, M.J.; Sánchez, M.; Gómez-Guzmán, M.; Vargas, F.; O’Valle, F.; Zarzuelo, A.; Pérez-Vizcaíno, F.; et al. Wine Polyphenols Improve Endothelial Function in Large Vessels of Female Spontaneously Hypertensive Rats. Hypertension 2008, 51, 1088–1095. [Google Scholar] [CrossRef] [PubMed]
- Schramm, D.D.; Wang, J.F.; Holt, R.R.; Ensunsa, J.L.; Gonsalves, J.L.; Lazarus, S.A.; Schmitz, H.H.; German, J.B.; Keen, C.L. Chocolate Procyanidins Decrease the Leukotriene-Prostacyclin Ratio in Humans and Human Aortic Endothelial Cells. Am. J. Clin. Nutr. 2001, 73, 36–40. [Google Scholar] [CrossRef] [Green Version]
- Facino, R.M.; Carini, M.; Aldini, G.; Berti, F.; Rossoni, G.; Bombardelli, E.; Morazzoni, P. Diet Enriched with Procyanidins Enhances Antioxidant Activity and Reduces Myocardial Post-Ischaemic Damage in Rats. Life Sci. 1999, 64, 627–642. [Google Scholar] [CrossRef]
- Sacks, D.; Baxter, B.; Campbell, B.C.V.; Carpenter, J.S.; Cognard, C.; Dippel, D.; Eesa, M.; Fischer, U.; Hausegger, K.; Hirsch, J.A.; et al. Multisociety Consensus Quality Improvement Revised Consensus Statement for Endovascular Therapy of Acute Ischemic Stroke. Int. J. Stroke 2018, 13, 612–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rat Primers | Sequence (5′…3′) | Amplicon Size | Efficiency | GenBank Accesion No. |
|---|---|---|---|---|
| eNOS Fw | GGATTCTGGCAAGACCGATTAC | 159 | 2.23 (111.5%) | NM_021838.2 |
| eNOS Rv | GGTGAGGACTTGTCCAAACACT | |||
| Sirt1 Fw | TTGGCACCGATCCTCGAA | 217 | 1.97 (98.5) | XM_006223877.1 |
| Sirt1 Rv | ACAGAAACCCCAGCTCCA | |||
| Nox4 Fw | GTGTCTGCATGGTGGTGGTA | 150 | 1.86 (93%) | NM_053524.1 |
| Nox4 Rv | TCAACAAGCCACCCGAAACA | |||
| Et1 Fw | TGATTCTCTTGCCTCTTCTTG | 110 | 2.23 (111.5%) | NM_012548.2 |
| Et1 Rv | TATGGAATCTCCTGGCTCTC | |||
| Ppia Fw | CTTCGAGCTGTTTGCAGACAA | 118 | 2.28 (114%) | NM_017101.1 |
| Ppia Rv | AAGTCACCACCCTGGCACATG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Fernández-Sobrino, R.; Soliz-Rueda, J.R.; Ávila-Román, J.; Arola-Arnal, A.; Suárez, M.; Muguerza, B.; Bravo, F.I. Blood Pressure-Lowering Effect of Wine Lees Phenolic Compounds Is Mediated by Endothelial-Derived Factors: Role of Sirtuin 1. Antioxidants 2021, 10, 1073. https://doi.org/10.3390/antiox10071073
López-Fernández-Sobrino R, Soliz-Rueda JR, Ávila-Román J, Arola-Arnal A, Suárez M, Muguerza B, Bravo FI. Blood Pressure-Lowering Effect of Wine Lees Phenolic Compounds Is Mediated by Endothelial-Derived Factors: Role of Sirtuin 1. Antioxidants. 2021; 10(7):1073. https://doi.org/10.3390/antiox10071073
Chicago/Turabian StyleLópez-Fernández-Sobrino, Raúl, Jorge R. Soliz-Rueda, Javier Ávila-Román, Anna Arola-Arnal, Manuel Suárez, Begoña Muguerza, and Francisca Isabel Bravo. 2021. "Blood Pressure-Lowering Effect of Wine Lees Phenolic Compounds Is Mediated by Endothelial-Derived Factors: Role of Sirtuin 1" Antioxidants 10, no. 7: 1073. https://doi.org/10.3390/antiox10071073
APA StyleLópez-Fernández-Sobrino, R., Soliz-Rueda, J. R., Ávila-Román, J., Arola-Arnal, A., Suárez, M., Muguerza, B., & Bravo, F. I. (2021). Blood Pressure-Lowering Effect of Wine Lees Phenolic Compounds Is Mediated by Endothelial-Derived Factors: Role of Sirtuin 1. Antioxidants, 10(7), 1073. https://doi.org/10.3390/antiox10071073