
Comparison of Phenolic Contents and Scavenging Activities of Miang Extracts Derived from Filamentous and Non-Filamentous Fungi-Based Fermentation Processes

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Preparation of Tea Extracts
2.3. Analysis of the Chemical Composition
2.3.1. Total Polyphenol Content
2.3.2. Total Flavonoid Content
2.3.3. Total Tannin Content
2.3.4. Condensed Tannin Content
2.3.5. High Performance Liquid Chromatography (HPLC)
2.4. In Vitro Antioxidant Assays
2.4.1. DPPH Radical Scavenging Activity Assay
2.4.2. ABTS Radical Scavenging Activity Assay
2.4.3. Ferric Reducing Antioxidant Power (FRAP) Assay
2.5. Inhibition of Induced Intracellular ROS Production
2.6. Inhibition of Pro-Inflammatory Nitric Oxide (NO) Production
2.7. Statistical Analysis
3. Results and Discussion
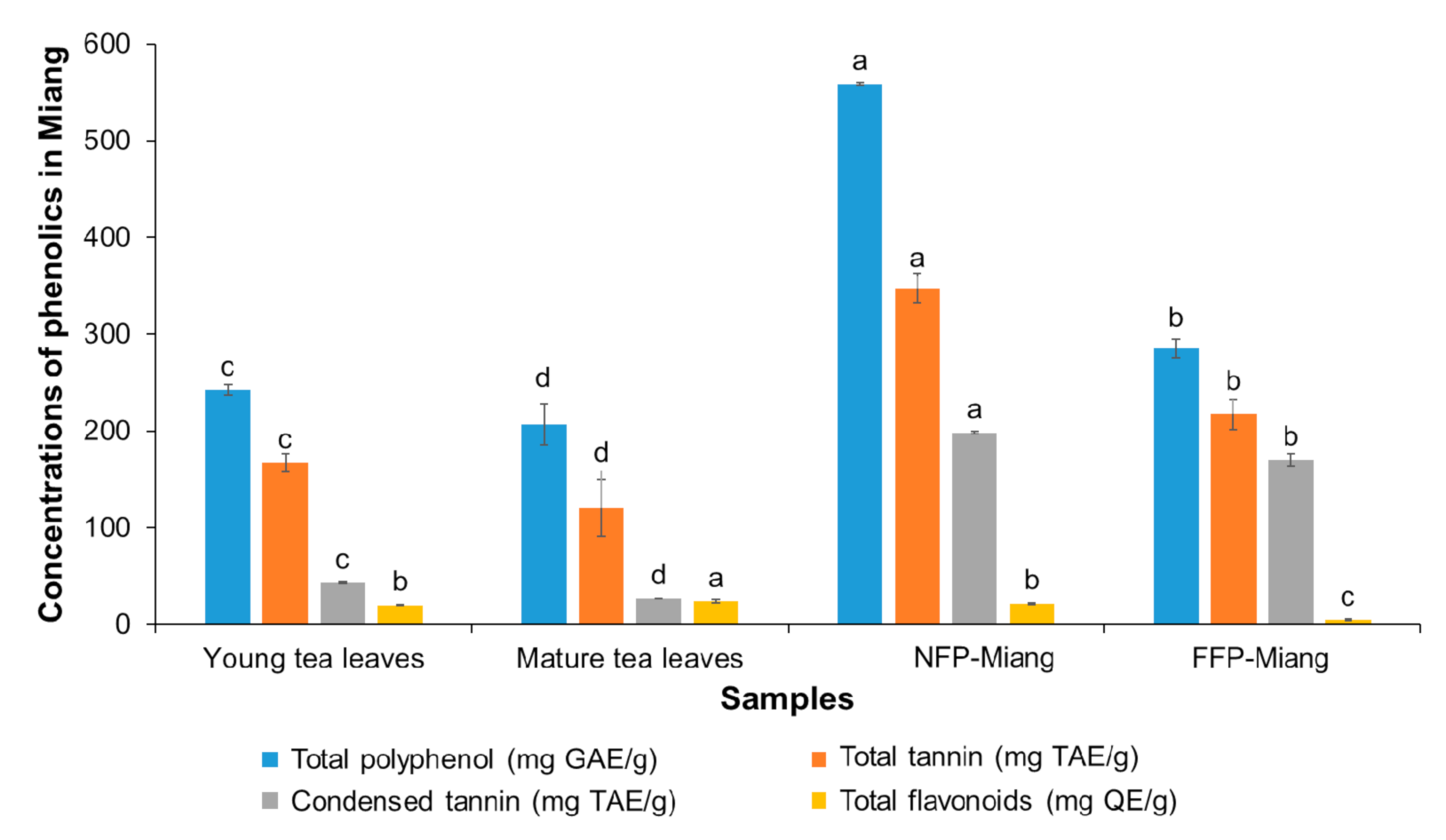
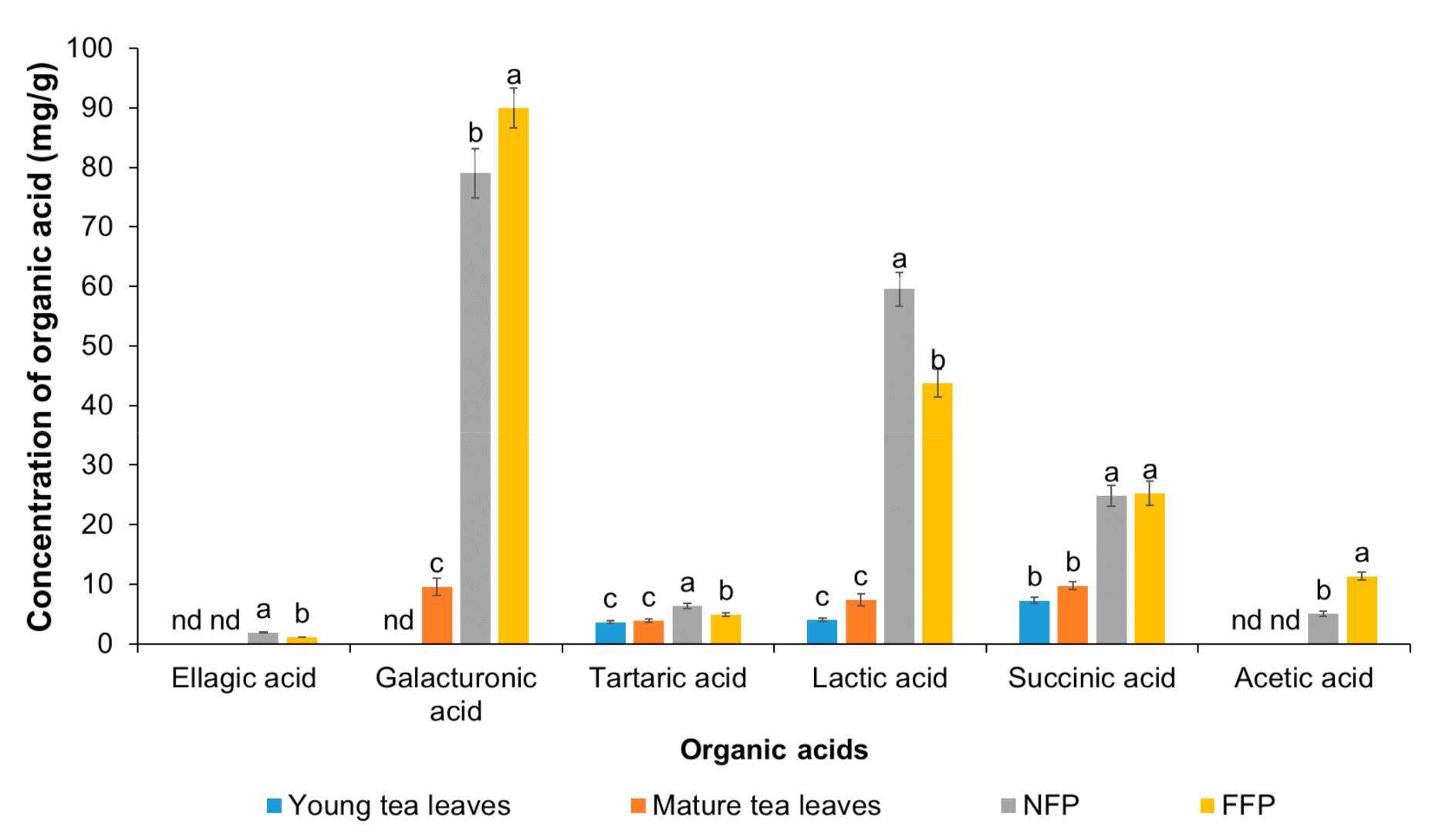
3.1. Chemical Composition of Miang
3.2. Antioxidant Activities of Miang Extracts
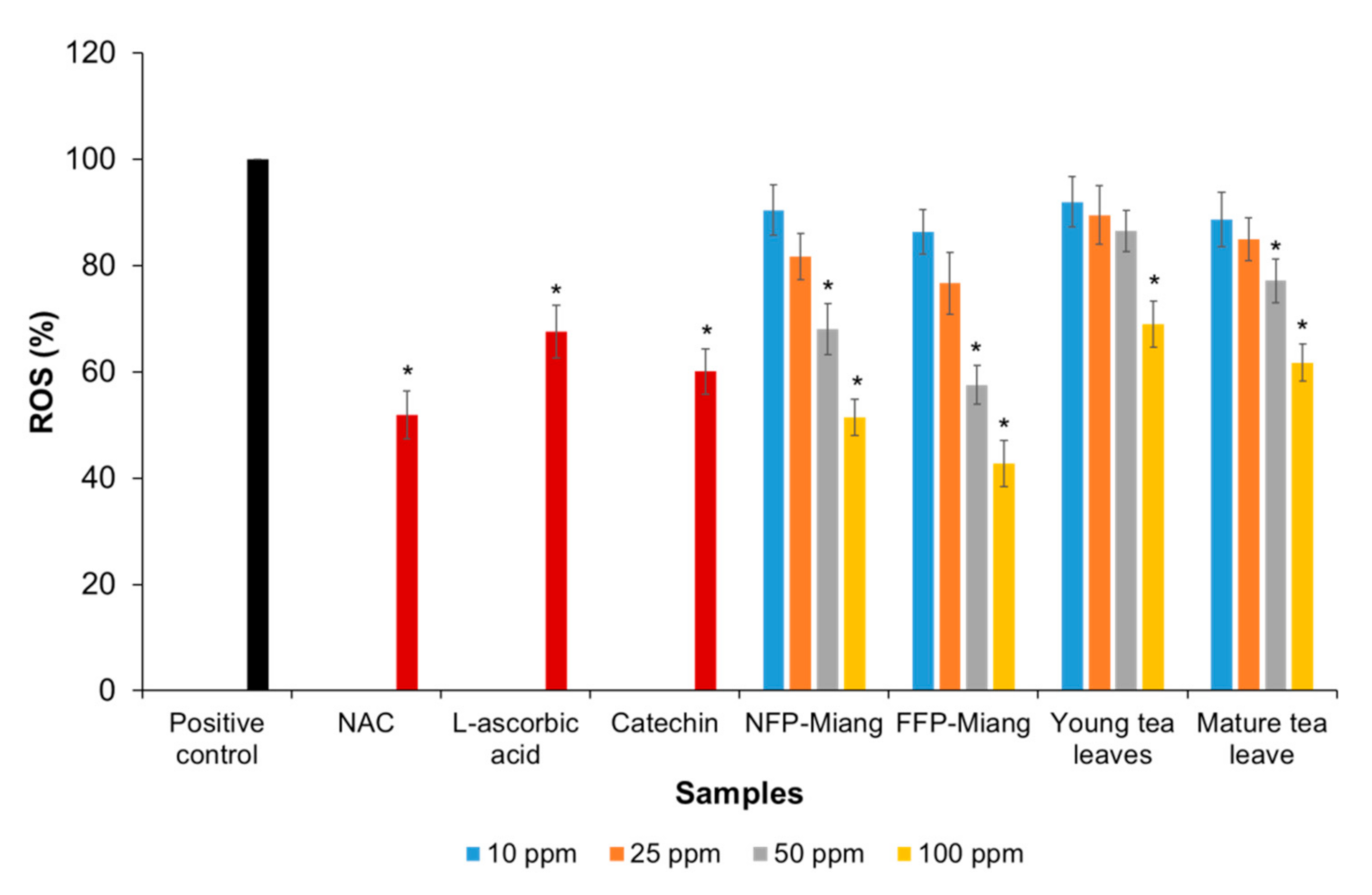
3.3. Inhibition of the Intracellular Production of Reactive Oxygen Species (ROS) in a Cell-Based Study
3.4. Inhibition of Nitric Oxide (NO) Production
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, N.; Mukhtar, H. Tea polyphenols in promotion of human health. Nutrients 2018, 11, 39. [Google Scholar] [CrossRef] [Green Version]
- Shukla, Y. Tea and cancer chemoprevention: A comprehensive review. Asian Pac. J. Cancer Prev. 2007, 8, 155–166. [Google Scholar]
- Sinha, D.; Biswas, J.; Nabavi, S.M.; Bishayee, A. Tea phytochemicals for breast cancer prevention and intervention: From bench to bedside and beyond. Sem. Cancer Biol. 2017, 46, 33–54. [Google Scholar] [CrossRef]
- Chakravorty, S.; Bhattacharya, S.; Bhattacharya, D.; Sarkar, S.; Gachhui, R. Kombucha: A promising functional beverage prepared from tea. In Non-Alcoholic Beverages; Grumezescu, A.M., Holban, A.M., Eds.; Elsevier: Cambridge, UK, 2019; Volume 6, pp. 285–327. [Google Scholar]
- Zhao, X.; Song, J.-L.; Kim, J.-D.; Lee, J.-S.; Park, K.-Y. Fermented pu-erh tea increases in vitro anticancer activities in HT-29 cells and has antiangiogenetic effects on HUVECS. J. Environ. Pathol. Toxicol. Oncol. 2013, 32, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Khanongnuch, C.; Unban, K.; Kanpiengjai, A.; Saenjum, C. Recent research advances and ethno-botanical history of miang, a traditional fermented tea (Camellia sinensis var. assamica) of northern Thailand. J. Ethn. Foods 2017, 4, 135–144. [Google Scholar] [CrossRef]
- Kodchasee, P.; Nain, K.; Abdullahi, A.D.; Unban, K.; Saenjum, C.; Shetty, K.; Khanongnuch, C. Microbial dynamics-linked properties and functional metabolites during miang fermentation using the filamentous fungi growth-based process. Food Biosci. 2021, 41, 14. [Google Scholar] [CrossRef]
- Unban, K.; Khatthongngam, N.; Pattananandecha, T.; Saenjum, C.; Shetty, K.; Khanongnuch, C. Microbial community dynamics during the non-filamentous fungi growth-based fermentation process of miang, a traditional fermented tea of north Thailand and their product characterizations. Front. Microbiol. 2020, 11, 1515. [Google Scholar] [CrossRef]
- Unban, K.; Khatthongngam, N.; Shetty, K.; Khanongnuch, C. Nutritional biotransformation in traditional fermented tea (miang) from north Thailand and its impact on antioxidant and antimicrobial activities. J. Food Sci. Technol. 2019, 56, 2687–2699. [Google Scholar] [CrossRef]
- Chen, X.; Li, Y.; Lin, Q.; Wang, Y.; Sun, H.; Wang, J.; Cui, G.; Cai, L.; Dong, X. Tea polyphenols induced apoptosis of breast cancer cells by suppressing the expression of Survivin. Sci. Rep. 2014, 4, 4416. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.-P.; Peng, C.-X.; Gao, B.; Gong, J.-S. Influence of large molecular polymeric pigments isolated from fermented zijuan tea on the activity of key enzymes involved in lipid metabolism in rat. Exp. Gerontol. 2012, 47, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.-Y.; Zhao, C.-N.; Xu, X.-Y.; Gan, R.-Y.; Cao, S.-Y.; Liu, Q.; Shang, A.; Mao, Q.-Q.; Li, H.-B. Phytochemical composition and antioxidant capacity of 30 Chinese teas. Antioxidants 2019, 8, 180. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.A.; Huber, S.A.; Lucas, Z.J. Inhibition of DNA synthesis in cultured lymphocytes and tumor cells by extracts of betel nut, tobacco, and miang leaf, plant substances associated with cancer of the ororespiratory epithelium. Cancer Res. 1979, 39, 4802–4809. [Google Scholar]
- Padma, R.; Parvathy, N.G.; Renjith, V.; Rahate, K. Quantitative estimation of tannins, phenols and antioxidant activity of methanolic extract of Imperata cylindrica. Int. J. Res. Pharm. Sci. 2013, 4, 73–77. [Google Scholar]
- Eom, S.-H.; Park, H.-J.; Jin, C.-W.; Kim, D.-O.; Seo, D.-W.; Jeong, Y.-H.; Cho, D.-H. Changes in antioxidant activity with temperature and time in Chrysanthemum indicum L. (gamguk) teas during elution processes in hot water. Food Sci. Biotechnol. 2008, 17, 408–412. [Google Scholar]
- Makkar, H.P.S.; Blümmel, M.; Borowy, N.K.; Becker, K. Gravimetric determination of tannins and their correlations with chemical and protein precipitation methods. J. Sci. Food Agric. 1993, 61, 161–165. [Google Scholar] [CrossRef]
- Porter, L.J.; Hrstich, L.N.; Chan, B.G. The conversion of procyanidins and prodelphinidins to cyanidin and delphinidin. Phytochemistry 1985, 25, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Moreno, C.; Larrauri, J.A.; Saura-Calixto, F. A procedure to measure the antiradical efficiency of polyphenols. J. Sci. Food Agric. 1998, 76, 270–276. [Google Scholar] [CrossRef]
- Radulović, N.S.; Randjelović, P.J.; Stojanović, N.M.; Blagojević, P.D.; Stojanović-Radić, Z.Z.; Ilić, I.R.; Djordjević, V.B. Toxic essential oils. Part II: Chemical, toxicological, pharmacological and microbiological profiles of Artemisia annua L. volatiles. Food Chem. Toxicol. 2013, 58, 37–49. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saenjum, C.; Pattananandecha, T.; Nakagawa, K. Detection of antioxidant phytochemicals isolated from Camellia japonica seeds using HPLC and EPR imaging. Antioxidants 2020, 9, 493. [Google Scholar] [CrossRef]
- Phromnoi, K.; Suttajit, M.; Saenjum, C.; Limtrakul (Dejkriengkraikul), P. Inhibitory effect of a rosmarinic acid-enriched fraction prepared from nga-mon (Perilla frutescens) seed meal on osteoclastogenesis through the RANK signaling pathway. Antioxidants 2021, 10, 307. [Google Scholar] [CrossRef]
- Sirithunyalug, B.; Saenjum, C.; Charumanee, S.; Sivamaruthi, B.S.; Chaiyasut, C.; Sirithunyalug, J.; Tipduangta, P. Development of colorectal-targeted dietary supplement tablets containing natural purple rice bran oil as a colorectal chemopreventive. Nutrients 2018, 10, 444. [Google Scholar] [CrossRef] [Green Version]
- Ares, A.M.; Valverde, S.; Bernal, J.L.; Nozal, M.J.; Bernal, J. Extraction and determination of bioactive compounds from bee pollen. J. Pharm. Biomed. Anal. 2018, 147, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Kuźma, P.; Drużyńska, B.; Obiedziński, M. Optimization of extraction conditions of some polyphenolic compounds from parsley leaves (Petroselinum crispum). Acta Sci. Pol. Technol. Aliment. 2014, 13, 145–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adebo, O.A.; Medina-Meza, I.G. Impact of fermentation on the phenolic compounds and antioxidant activity of whole cereal grains: A mini review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salar, R.K.; Certik, M.; Brezova, V. Modulation of phenolic content and antioxidant activity of maize by solid state fermentation with Thamnidium elegans CCF 1456. Biotechnol. Bioprocess. Eng. 2012, 17, 109–116. [Google Scholar] [CrossRef]
- Wang, Y.; Kong, D.; Gao, Y.; Ying, L.; Huang, Q.; Xu, P. Chemical characterization and bioactivity of phenolics from tieguanyin oolong tea. J. Food Biochem. 2019, 43, e12894. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Compaoré-Sérémé, D.; Sawadogo-Lingani, H.; Coda, R.; Katina, K.; Maina, N.H. Influence of dextran synthesized in situ on the rheological, technological and nutritional properties of whole grain pearl millet bread. Food Chem. 2019, 285, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Monagas, M.; Urpi-Sarda, M.; Sánchez-Patán, F.; Llorach, R.; Garrido, I.; Gómez-Cordovés, C.; Andres-Lacueva, C.; Bartolomé, B. Insights into the metabolism and microbial biotransformation of dietary flavan-3-ols and the bioactivity of their metabolites. Food Funct. 2010, 1, 233–253. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-C.; Li, C.-W.; Shih, Y.-T.; Chuang, L.-T. Antioxidant and anti-inflammatory properties of lower-polymerized polyphenols in oolong. Int. J. Food Prop. 2014, 17, 752–764. [Google Scholar] [CrossRef]
- Kongpichitchoke, T.; Chiu, M.-T.; Huang, T.; Hsu, J.-L. Gallic acid content in Taiwanese teas at different degrees of fermentation and its antioxidant activity by inhibiting PKCδ activation: In vitro and in silico studies. Molecules 2016, 21, 1346. [Google Scholar] [CrossRef] [Green Version]
- Yildiz, E.; Guldas, M.; Gurbuz, O.; Yildiz, E.; Guldas, M.; Gurbuz, O. Determination of in-vitro phenolics, antioxidant capacity and bio-accessibility of kombucha tea produced from black carrot varieties grown in Turkey. Food Sci. Technol. 2021, 41, 180–187. [Google Scholar] [CrossRef]
- Weng, Y.-P.; Hung, P.-F.; Ku, W.-Y.; Chang, C.-Y.; Wu, B.-H.; Wu, M.-H.; Yao, J.-Y.; Yang, J.-R.; Lee, C.-H. The inhibitory activity of gallic acid against DNA methylation: Application of gallic acid on epigenetic therapy of human cancers. Oncotarget 2018, 9, 361–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Gennaro, P.; Sabatini, V.; Fallarini, S.; Pagliarin, R.; Sello, G. Polyphenol polymerization by an alternative oxidative microbial enzyme and characterization of the biological activity of oligomers. Biomed. Res. Int. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, W.D. Condensed tannins. In Toxic Substances in Crop Plants; D’Mello, F.J.P., Duffus, C.M., Duffus, J.H., Eds.; Elsevier: Cambridge, UK, 1991; pp. 180–201. [Google Scholar]
- Schofield, P.; Mbugua, D.M.; Pell, A.N. Analysis of condensed tannins: A review. Anim. Feed Sci. Technol. 2001, 91, 21–40. [Google Scholar] [CrossRef]
- Monobe, M.; Ema, K.; Tokuda, Y.; Maeda-Yamamoto, M. Green tea catechin induced phagocytosis can be blocked by catalase and an inhibitor of transient receptor potential melastatin 2 (TRPM2). Cytotechnology 2014, 66, 561–566. [Google Scholar] [CrossRef] [Green Version]
- Laks, P.E. Chemistry of the condensed tannin b-ring. In Chemistry and Significance of Condensed Tannins; Hemingway, R.W., Karchesy, J.J., Branham, S.J., Eds.; Springer US: Boston, MA, USA, 1989; pp. 249–263. [Google Scholar]
- Weerawatanakorn, M. Chemistry and health beneficial effects of oolong tea and theasinensins. Food Sci. Hum. Well. 2015, 4, 133–146. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.C.; Chang, F.S.; Chen, I.Z.; Lu, F.M.; Cheng, T.J.; Chen, R.L. Redox potential of tea infusion as an index for the degree of fermentation. Anal. Chim. Acta 2007, 594, 32–36. [Google Scholar] [CrossRef]
- Pereira-Caro, G.; Moreno-Rojas, J.M.; Brindani, N.; Del Rio, D.; Lean, M.E.J.; Hara, Y.; Crozier, A. Bioavailability of black tea theaflavins: Absorption, metabolism, and colonic catabolism. J. Agric. Food Chem. 2017, 65, 5365–5374. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Guo, Y.; Yang, J.; Henning, S.M.; Lee, R.-P.; Rasmussen, A.; Zhang, L.; Lu, Q.-Y.; Heber, D.; Li, Z. Bioavailability and bioactivity of free ellagic acid compared to pomegranate juice. Food Funct. 2019, 10, 6582–6588. [Google Scholar] [CrossRef] [PubMed]
- Belwal, T.; Nabavi, S.F.; Nabavi, S.M.; Habtemariam, S. Dietary anthocyanins and insulin resistance: When food becomes a medicine. Nutrients 2017, 9, 1111. [Google Scholar] [CrossRef]
- Larrosa, M.; González-Sarrías, A.; García-Conesa, M.T.; Tomás-Barberán, F.A.; Espín, J.C. Urolithins, ellagic acid-derived metabolites produced by human colonic microflora, exhibit estrogenic and antiestrogenic activities. J. Agric. Food Chem. 2006, 54, 1611–1620. [Google Scholar] [CrossRef]
- Kasimsetty, S.G.; Bialonska, D.; Reddy, M.K.; Ma, G.; Khan, S.I.; Ferreira, D. Colon cancer chemopreventive activities of pomegranate ellagitannins and urolithins. J. Agric. Food Chem. 2010, 58, 2180–2187. [Google Scholar] [CrossRef]
- Larrosa, M.; González-Sarrías, A.; Yáñez-Gascón, M.J.; Selma, M.V.; Azorín-Ortuño, M.; Toti, S.; Tomás-Barberán, F.; Dolara, P.; Espín, J.C. Anti-inflammatory properties of a pomegranate extract and its metabolite urolithin-A in a colitis rat model and the effect of colon inflammation on phenolic metabolism. J. Nutr. Biochem. 2010, 21, 717–725. [Google Scholar] [CrossRef]
- Vicinanza, R.; Zhang, Y.; Henning, S.M.; Heber, D. Pomegranate juice metabolites, ellagic acid and urolithin a, synergistically inhibit androgen-independent prostate cancer cell growth via distinct effects on cell cycle control and apoptosis. Evid. Based Complement. Alternat. Med. 2013, 2013, 247504. [Google Scholar] [CrossRef] [Green Version]
- Hoegger, P.; Majcherczyk, A.; Dwivedi, R.; Svobodová, K.; Kilaru, S.; Kües, U. Enzymes in wood degradation. In Wood Production, Wood Technology and Biotechnological Impacts; Kües, U., Ed.; Göttingen University Press: Göttingen, Germany, 2007; pp. 383–432. [Google Scholar]
- Pinkian, N.; Phuengjayaem, S.; Tanasupawat, S.; Teeradakorn, S. Optimization of succinic acid production by succinic acid bacteria isolated in Thailand. Warasan Songkhla Nakharin 2018, 40, 1281–1290. [Google Scholar]
- Wee, Y.-J.; Yun, J.-S.; Kang, K.-H.; Ryu, H.-W. Continuous production of succinic acid by a fumarate-reducing bacterium immobilized in a hollow-fiber bioreactor. Appl. Biochem. Biotechnol. 2002, 98–100, 1093–1104. [Google Scholar] [CrossRef]
- Abramovič, H.; Grobin, B.; Poklar Ulrih, N.; Cigić, B. Relevance and standardization of in vitro antioxidant assays: ABTS, DPPH, and folin–Ciocalteu. J. Chem. 2018, 2018, e4608405. [Google Scholar] [CrossRef] [Green Version]
- Shalaby, E.; Shanab, S. Comparison of DPPH and ABTS assays for determining antioxidant potential of water and methanol extracts of Spirulina platensis. Indian J. Mar. Sci. 2013, 42, 556–564. [Google Scholar]
- Barros, A.I.R.N.A.; Nunes, F.M.; Gonçalves, B.; Bennett, R.N.; Silva, A.P. Effect of cooking on total vitamin C contents and antioxidant activity of sweet chestnuts (Castanea sativa Mill.). Food Chem. 2011, 128, 165–172. [Google Scholar] [CrossRef]
- Ozturk Sarikaya, S.B. Acethylcholinesterase inhibitory potential and antioxidant properties of pyrogallol. J. Enzym. Inhib. Med. Chem. 2015, 30, 761–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes, V.; Vilaça, R.; de Freitas, V.; Ferreira, P.M.; Mateus, N.; Costa, V. Effect of myricetin, pyrogallol, and phloroglucinol on yeast resistance to oxidative stress. Oxid. Med. Cell. Longev. 2015, 2015, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Sroka, Z.; Cisowski, W. Hydrogen peroxide scavenging, antioxidant and anti-radical activity of some phenolic acids. Food Chem. Toxicol. 2003, 41, 753–758. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Sevgi, K.; Tepe, B.; Sarikurkcu, C. Antioxidant and DNA damage protection potentials of selected phenolic acids. Food Chem. Toxicol. 2015, 77, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Verma, B.; Hucl, P.; Chibbar, R.N. Phenolic content and antioxidant properties of bran in 51 wheat cultivars. Cereal Chem. 2008, 85, 544–549. [Google Scholar] [CrossRef]
- Ahmed, E.; Holmström, S.J.M. Siderophores in environmental research: Roles and applications. Microb. Biotechnol. 2014, 7, 196–208. [Google Scholar] [CrossRef]
- Gülçin, İ. Antioxidant activity of food constituents: An overview. Arch. Toxicol. 2012, 86, 345–391. [Google Scholar] [CrossRef]
- Gargi, S.; Nilanjan, S.; Moutusi, N.; Subhasis, M. Bioactive components of tea. Arch. Food Nutr. Sci. 2020, 4, 1–9. [Google Scholar]
- Brune, M.; Rossander, L.; Hallberg, L. Iron absorption and phenolic compounds: Importance of different phenolic structures. Eur. J. Clin. Nutr. 1989, 43, 547–557. [Google Scholar]
- Rp, G.; Gm, W.; Pk, S.; Dd, M. Inhibition of iron uptake by phytic acid, tannic acid, and ZnCl2: Studies using an in vitro digestion/Caco-2 cell model. J. Agric. Food Chem. 2002, 50, 390–395. [Google Scholar]
- Welch, R.M.; Graham, R.D. Breeding for micronutrients in staple food crops from a human nutrition perspective. J. Exp. Bot. 2004, 55, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Groussard, C.; Morel, I.; Chevanne, M.; Monnier, M.; Cillard, J.; Delamarche, A. Free radical scavenging and antioxidant effects of lactate ion: An in vitro study. J. Appl. Physiol. 2000, 89, 169–175. [Google Scholar] [CrossRef]
- Kumar, A.P.; Chougala, M.; Nandini, C.D.; Salimath, P.V. Effect of butyric acid supplementation on serum and renal antioxidant enzyme activities in streptozotocin-induced diabetic rats: Butyric acid supplementation on antioxidant enzymes. J. Food Biochem. 2010, 34, 15–30. [Google Scholar] [CrossRef]
- Badhani, B.; Sharma, N.; Kakkar, R. Gallic acid: A versatile antioxidant with promising therapeutic and industrial applications. R. Soc. Chem. Adv. 2015, 5, 27540–27557. [Google Scholar] [CrossRef]
- Vattem, D.A.; Shetty, K. Biological functionality of ellagic acid: A review. J. Food Biochem. 2005, 29, 234–266. [Google Scholar] [CrossRef]
- Ivanišová, E.; Meňhartová, K.; Terentjeva, M.; Harangozo, Ľ.; Kántor, A.; Kačániová, M. The evaluation of chemical, antioxidant, antimicrobial and sensory properties of kombucha tea beverage. J. Food Sci. Technol. 2020, 57, 1840–1846. [Google Scholar] [CrossRef] [PubMed]
- Villarreal-Soto, S.A.; Beaufort, S.; Bouajila, J.; Souchard, J.-P.; Renard, T.; Rollan, S.; Taillandier, P. Impact of fermentation conditions on the production of bioactive compounds with anticancer, anti-inflammatory and antioxidant properties in kombucha tea extracts. Process. Biochem. 2019, 83, 44–54. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; Guo, Q.; Xin, W. Free radical scavenging by green tea polyphenols. Meth. Enzymol. 2001, 335, 217–231. [Google Scholar]
- Chen, C.; Kong, A.-N.T. Dietary chemopreventive compounds and ARE/EpRE signaling. Free Radic. Biol. Med. 2004, 36, 1505–1516. [Google Scholar] [CrossRef]
- Masella, R.; Di Benedetto, R.; Varì, R.; Filesi, C.; Giovannini, C. Novel mechanisms of natural antioxidant compounds in biological systems: Involvement of glutathione and glutathione-related enzymes. J. Nutr. Biochem. 2005, 16, 577–586. [Google Scholar] [CrossRef]
- De Vries, H.E.; Witte, M.; Hondius, D.; Rozemuller, A.J.M.; Drukarch, B.; Hoozemans, J.; van Horssen, J. Nrf2-induced antioxidant protection: A promising target to counteract ROS-mediated damage in neurodegenerative disease? Free Radic. Biol. Med. 2008, 45, 1375–1383. [Google Scholar] [CrossRef]
- Heiss, E.H.; Schachner, D.; Zimmermann, K.; Dirsch, V.M. Glucose availability is a decisive factor for Nrf2-mediated gene expression. Redox Biol. 2013, 1, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.; Zhong, Y.; Duan, Y.; Chen, Q.; Li, F. Antioxidant mechanism of tea polyphenols and its impact on health benefits. Anim. Nutr. 2020, 6, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Priscilla, D.H.; Prince, P.S.M. Cardioprotective effect of gallic acid on cardiac troponin-T, cardiac marker enzymes, lipid peroxidation products and antioxidants in experimentally induced myocardial infarction in wistar rats. Chem. Biol. Interact. 2009, 179, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Debnath, B.; Singh, W.; Das, M.; Goswami, S.; Manna, K. Biodynamic activities of ellagic acid: A dietary polyphenol. J. Nat. Sci. Med. 2020, 3, 83–90. [Google Scholar]
- Bhattacharya, S.; Chatterjee, S.; Manna, P.; Das, J.; Ghosh, J.; Gachhui, R.; Sil, P.C. Prophylactic role of D-saccharic acid-1,4-lactone in tertiary butyl hydroperoxide induced cytotoxicity and cell death of murine hepatocytes via mitochondria-dependent pathways. J. Biochem. Mol. Toxicol. 2011, 25, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Shah, N.P. Synergistic application of black tea extracts and lactic acid bacteria in protecting human colonocytes against oxidative damage. J. Agric. Food Chem. 2016, 64, 2238–2246. [Google Scholar] [CrossRef]
- Luceri, C.; Caderni, G.; Sanna, A.; Dolara, P. Red wine and black tea polyphenols modulate the expression of cycloxygenase-2, inducible nitric oxide synthase and glutathione-related enzymes in azoxymethane-induced f344 rat colon tumors. J. Nutr. 2002, 132, 1376–1379. [Google Scholar] [CrossRef]
- Luczaj, W.; Skrzydlewska, E. Antioxidative properties of black tea. Prev. Med. 2005, 40, 910–918. [Google Scholar] [CrossRef]
- Marzban, F.; Azizi, G.; Afraei, S.; Sedaghat, R.; Seyedzadeh, M.H.; Razavi, A.; Mirshafiey, A. Kombucha tea ameliorates experimental autoimmune encephalomyelitis in mouse model of multiple sclerosis. Food Agric. Immunol. 2015, 26, 782–793. [Google Scholar] [CrossRef] [Green Version]
- Mo, H.; Zhu, Y.; Chen, Z. Microbial fermented tea – a potential source of natural food preservatives. Trends Food Sci. Technol. 2008, 19, 124–130. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, Q.; Li, G.; Chen, F.; Qian, Y.; Wang, R. In vitro antioxidant, anti-mutagenic, anti-cancer and anti-angiogenic effects of Chinese Bowl tea. J. Funct. Foods 2014, 7, 590–598. [Google Scholar] [CrossRef]
- Wang, Q.; Peng, C.; Gong, J.; Sirisansaneeyakul, S. Antioxidative effect of large molecular polymeric pigments extracted from zijuan pu-erh tea in vitro and in vivo. Kasetsart J. Nat. Sci. 2013, 47, 734–747. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Bioactive Compounds (mg/g Sample) | ||||||
|---|---|---|---|---|---|---|---|
| Catechin (C) | Epicatechin (EC) | Epigallocatechin Gallate (EGCG) | Epicatechin Gallate (ECG) | Pyrogallol | Gallic Acid (GA) | Caffeine | |
| Young leaves | 6.97 ± 0.14 b | 2.07 ± 0.08 a | 0.04 ± 0.003 d | 1.13 ± 0.06 b | nd | 0.57 ± 0.07 b | 1.04 ± 0.08 c |
| Mature leaves | 21.82 ± 1.13 a | 1.75 ± 0.06 b | 0.05 ± 0.002 c | 2.03 ± 0.07 a | nd | 0.26 ± 0.03 c | 1.98 ± 0.07 a |
| NFP-Miang | 6.89 ± 0.11 b | nd | 0.07 ± 0.003 a | 1.15 ± 0.07 b | 0.21 ± 0.007 b | 1.06 ± 0.02 a | 1.23 ± 0.08 b |
| FFP-Miang | 4.75 ± 0.13 c | nd | 0.06 ± 0.003 b | nd | 0.26 ± 0.005 a | 0.68 ± 0.03 b | 0.73 ± 0.05 d |
| Sample | DPPH IC50 (μg/mL) | ABTS IC50 (μg/mL) | FRAP (μmol Fe II/g) |
|---|---|---|---|
| Young leaves | 34.52 ± 4.57 b | 36.98 ± 5.10 b | 7880.49 ± 561.82 b |
| Mature leaves | 37.76 ± 7.42 c | 42.10 ± 6.74 c | 5004.69 ± 358.92 c |
| NFP-Miang | 13.15 ± 2.26 a | 18.50 ± 2.88 a | 9266.47 ± 707.67 a |
| FFP-Miang | 9.31 ± 2.92 a | 13.87 ± 6.38 a | 7236.44 ± 788.23 b |
| Ascorbic acid | 7.39 ± 0.67 a | 8.85 ± 1.96 a | - |
| Iron (II) sulfate | - | - | 3597.12 ± 0.00 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdullahi, A.D.; Kodchasee, P.; Unban, K.; Pattananandecha, T.; Saenjum, C.; Kanpiengjai, A.; Shetty, K.; Khanongnuch, C. Comparison of Phenolic Contents and Scavenging Activities of Miang Extracts Derived from Filamentous and Non-Filamentous Fungi-Based Fermentation Processes. Antioxidants 2021, 10, 1144. https://doi.org/10.3390/antiox10071144
Abdullahi AD, Kodchasee P, Unban K, Pattananandecha T, Saenjum C, Kanpiengjai A, Shetty K, Khanongnuch C. Comparison of Phenolic Contents and Scavenging Activities of Miang Extracts Derived from Filamentous and Non-Filamentous Fungi-Based Fermentation Processes. Antioxidants. 2021; 10(7):1144. https://doi.org/10.3390/antiox10071144
Chicago/Turabian StyleAbdullahi, Aliyu Dantani, Pratthana Kodchasee, Kridsada Unban, Thanawat Pattananandecha, Chalermpong Saenjum, Apinun Kanpiengjai, Kalidas Shetty, and Chartchai Khanongnuch. 2021. "Comparison of Phenolic Contents and Scavenging Activities of Miang Extracts Derived from Filamentous and Non-Filamentous Fungi-Based Fermentation Processes" Antioxidants 10, no. 7: 1144. https://doi.org/10.3390/antiox10071144
APA StyleAbdullahi, A. D., Kodchasee, P., Unban, K., Pattananandecha, T., Saenjum, C., Kanpiengjai, A., Shetty, K., & Khanongnuch, C. (2021). Comparison of Phenolic Contents and Scavenging Activities of Miang Extracts Derived from Filamentous and Non-Filamentous Fungi-Based Fermentation Processes. Antioxidants, 10(7), 1144. https://doi.org/10.3390/antiox10071144